
Association of Gut Lachnospiraceae and Chronic Spontaneous Urticaria


 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Extraction and 16S rRNA Gene Sequencing
2.2. Bioinformatics and Statistical Analysis
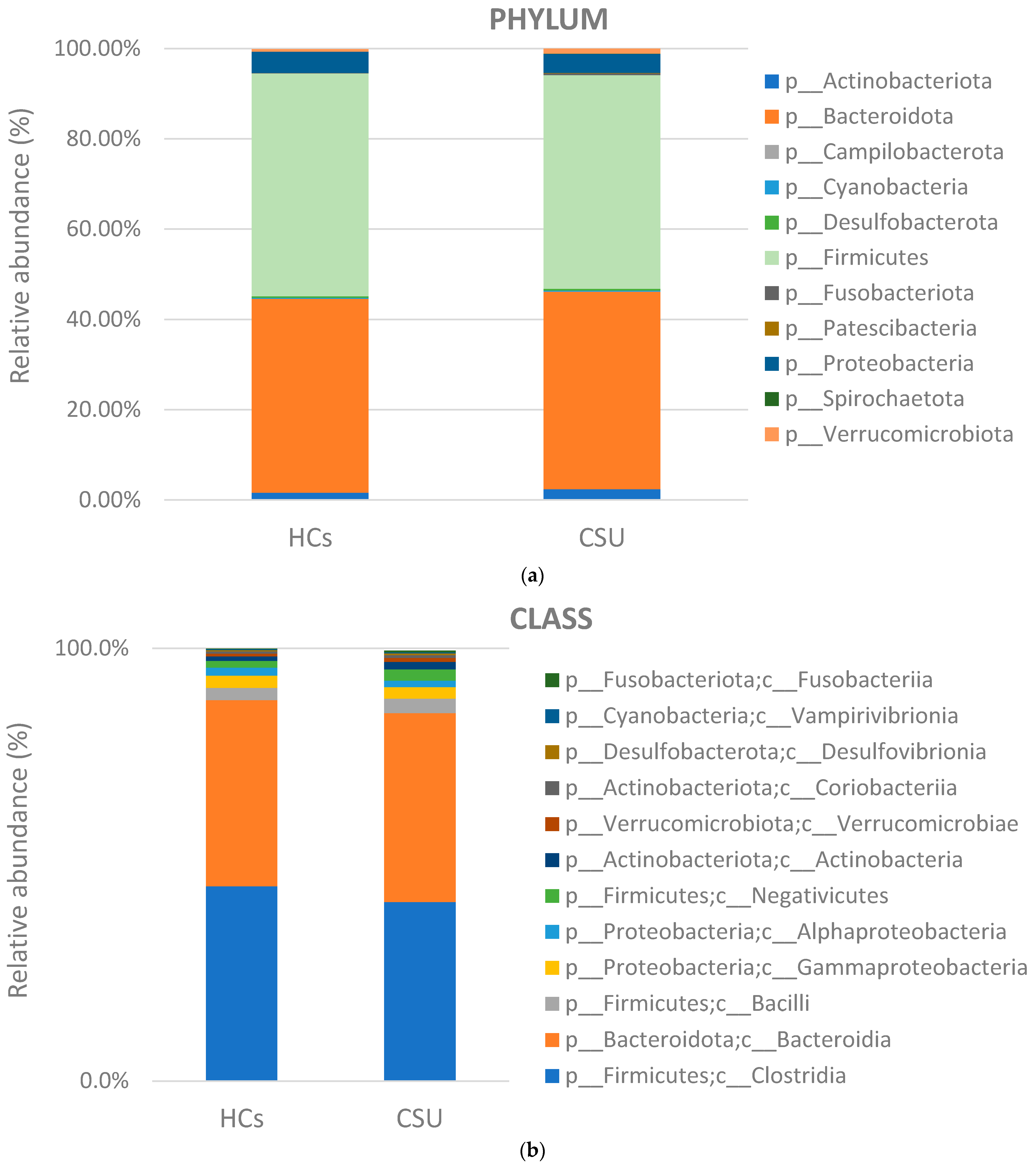
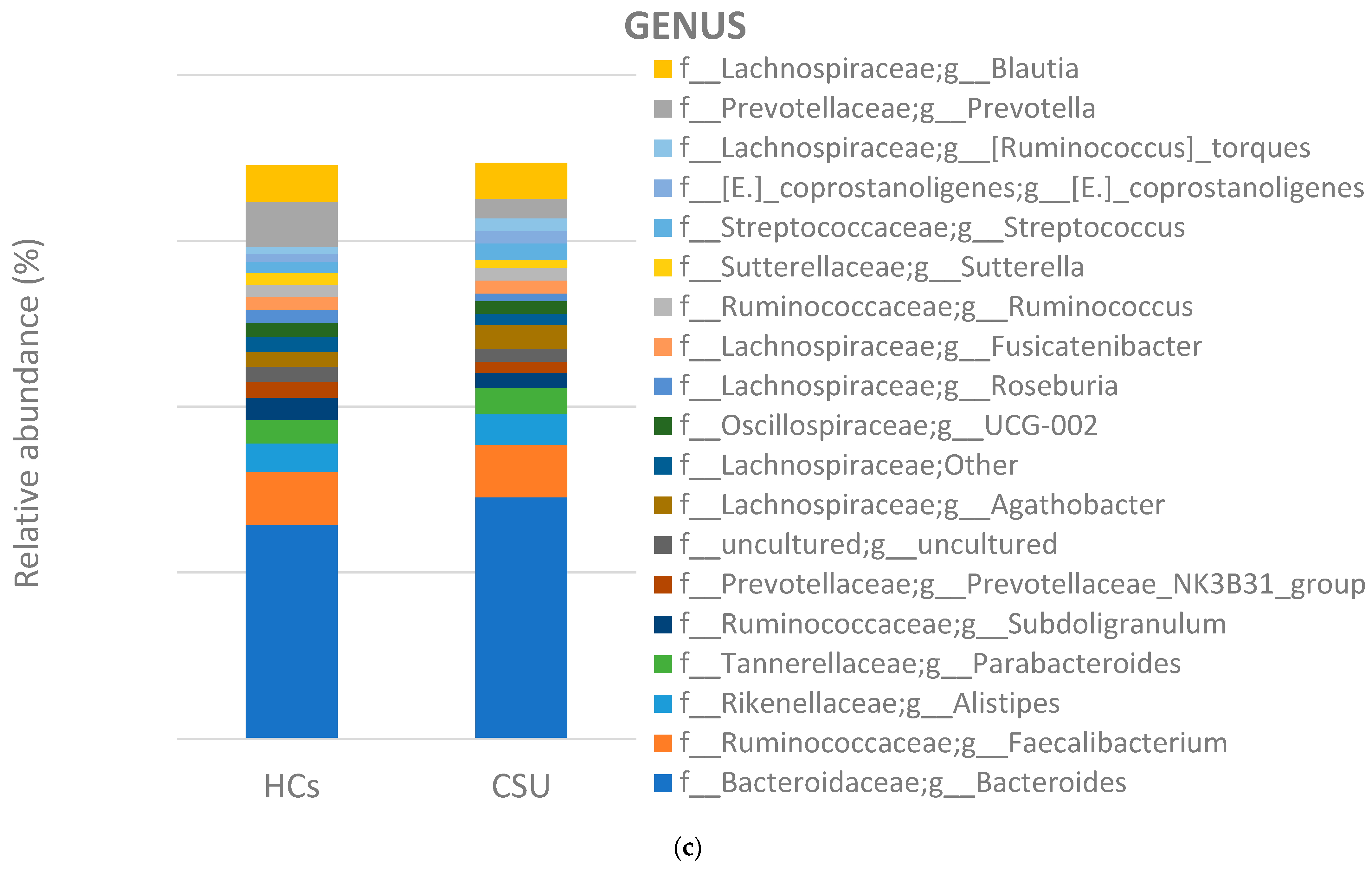
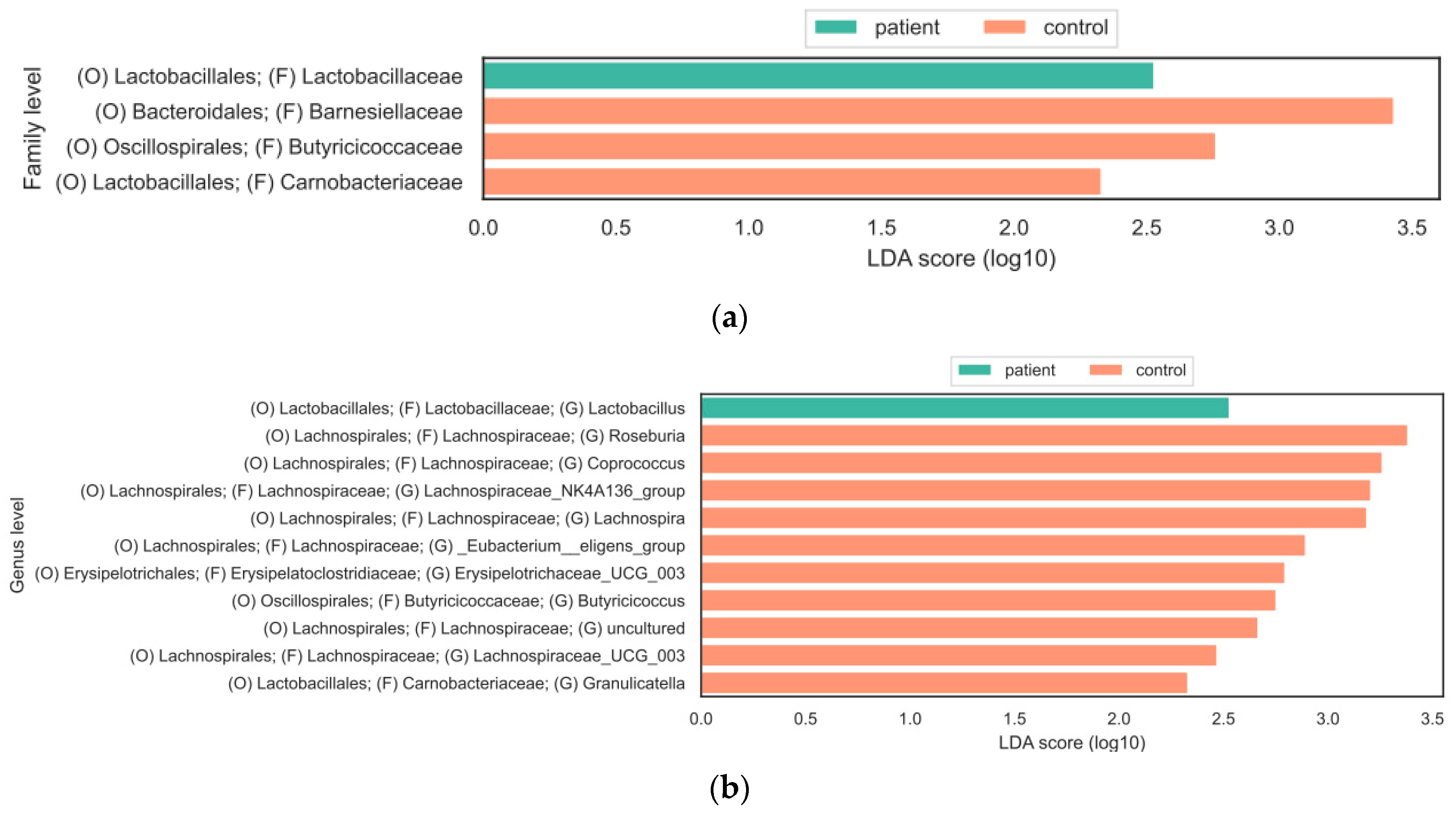
3. Results
3.1. Characterization of Participants
3.2. Composition of Gut Microbiota
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuberbier, T.; Abdul Latiff, A.H.; Abuzakouk, M.; Aquilina, S.; Asero, R.; Baker, D.; Ballmer-Weber, B.; Bangert, C.; Ben-Shoshan, M.; Bernstein, J.A.; et al. The international EAACI/GA2LEN/EuroGuiDerm/APAAACI guideline for the definition, classification, diagnosis, and management of urticaria. Allergy 2022, 77, 734–766. [Google Scholar] [CrossRef] [PubMed]
- Gonçalo, M.; Gimenéz-Arnau, A.; Al-Ahmad, M.; Ben-Shoshan, M.; Bernstein, J.; Ensina, L.; Fomina, D.; Galvàn, C.A.; Godse, K.; Grattan, C.; et al. The global burden of chronic urticaria for the patient and society. Br. J. Dermatol. 2021, 184, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.; Weller, K.; Bindslev-Jensen, C.; Giménez-Arnau, A.; Bousquet, P.J.; Bousquet, J.; Canonica, G.W.; Church, M.K.; Godse, K.V.; Grattan, C.E.H.; et al. Unmet clinical needs in chronic spontaneous urticaria. A GA2LEN task force report1. Allergy 2010, 66, 317–330. [Google Scholar] [CrossRef]
- Poddighe, D. The prevalence of chronic spontaneous urticaria (CSU) in the pediatric population. J. Am. Acad. Dermatol. 2019, 81, e149. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.; Abuzakouk, M.; Bérard, F.; Canonica, W.; Oude Elberink, H.; Giménez-Arnau, A.; Grattan, C.; Hollis, K.; Knulst, A.; Lacour, J.-P.; et al. The burden of chronic spontaneous urticaria is substantial: Real-world evidence from ASSURE-CSU. Allergy 2017, 72, 2005–2016. [Google Scholar] [CrossRef]
- Rabelo-Filardi, R.; de Oliveira, R.D.; Campos, R.A. Parameters Associated with Chronic Spontaneous Urticaria Duration and Severity: A Systematic Review. Int. Arch. Allergy Immunol. 2013, 161, 197–204. [Google Scholar] [CrossRef]
- Bracken, S.J.; Abraham, S.; MacLeod, A.S. Autoimmune Theories of Chronic Spontaneous Urticaria. Front. Immunol. 2019, 10, 627. [Google Scholar] [CrossRef]
- Chu, C.-Y.; Zuberbier, T. Urticaria and the gut. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 381–385. [Google Scholar]
- Lu, T.; Chen, Y.; Guo, Y.; Sun, J.; Shen, W.; Yuan, M.; Zhang, S.; He, P.; Jiao, X. Altered gut microbiota diversity and composition in chronic urticaria. Dis. Markers 2019, 2019, 6417471. [Google Scholar] [CrossRef]
- Liu, R.; Peng, C.; Jing, D.; Xiao, Y.; Zhu, W.; Zhao, S.; Zhang, J.; Chen, X.; Li, J. Biomarkers of Gut Microbiota in Chronic Spontaneous Urticaria and Symptomatic Dermographism. Front. Cell. Infect. Microbiol. 2021, 11, 703126. [Google Scholar] [CrossRef]
- Wang, D.; Guo, S.; He, H.; Gong, L.; Cui, H. Gut microbiome and serum metabolome analyses identify unsaturated fatty acids and butanoate metabolism induced by gut microbiota in patients with chronic spontaneous urticaria. Front. Cell. Infect. Microbiol. 2020, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yi, W.; He, L.; Luo, S.; Wang, J.; Jiang, L.; Long, H.; Zhao, M.; Lu, Q. Abnormalities in Gut Microbiota and Metabolism in Patients with Chronic Spontaneous Urticaria. Front. Immunol. 2021, 12, 691304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Chu, Z.; Shi, L.; Geng, S.; Guo, K. Gut Microbiome Alterations and Functional Prediction in Chronic Spontaneous Urticaria Patients. J. Microbiol. Biotechnol. 2021, 31, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Krišto, M.; Lugović-Mihić, L.; Muñoz, M.; Rupnik, M.; Mahnic, A.; Ozretić, P.; Jaganjac, M.; Ćesić, D.; Kuna, M. Gut Microbiome Composition in Patients with Chronic Urticaria: A Review of Current Evidence and Data. Life 2023, 13, 152. [Google Scholar] [CrossRef]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The Gut Microbiota and Inflammation: An Overview. Int. J. Environ. Res. Public Health 2020, 17, 7618. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How Colonization by Microbiota in Early Life Shapes the Immune System. Science 2016, 352, 539–544. [Google Scholar] [CrossRef]
- Power, S.E.; O’Toole, P.W.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F. Intestinal Microbiota, Diet and Health. Br. J. Nutr. 2014, 111, 387–402. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The Gut Microbiota—Masters of Host Development and Physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Thorburn, A.N.; Macia, L.; Mackay, C.R. Diet, Metabolites, and “Western-Lifestyle” Inflammatory Diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Vrancken, G.; Gregory, A.C.; Huys, G.R.B.; Faust, K.; Raes, J. Synthetic Ecology of the Human Gut Microbiota. Nat. Rev. Microbiol. 2019, 15, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, M. A Slow-Motion Epidemic. Nature 2016, 540, S98–S99. [Google Scholar] [CrossRef]
- Xiao, X.; Hu, X.; Yao, J.; Cao, W.; Zou, Z.; Wang, L. The Role of Short-Chain Fatty Acids in Inflammatory Skin Diseases. Front. Microbiol. 2023, 13, 1083432. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The Role of Short-Chain Fatty Acids in Immunity, Inflammation, and Metabolism. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–12. [Google Scholar] [CrossRef]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Intestinal Luminal Nitrogen Metabolism: Role of the Gut Microbiota and Consequences for the Host. Pharmacol. Res. 2013, 69, 114–126. [Google Scholar] [CrossRef]
- Abdugheni, R.; Wang, W.-Z.; Wang, Y.-J.; Du, M.-X.; Liu, F.-L.; Zhou, N.; Jiang, C.-Y.; Wang, C.-Y.; Wu, L.; Ma, J.; et al. Metabolite profiling of human-originated Lachnospiraceae at the strain level. iMeta 2022, 1, 100019. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, P.; Hu, R.; Qi, S.; Zhao, Y.; Miao, Y.; Han, Y.; Zhou, L.; Yang, Q. Gut Microbiota Characterization in Chinese Patients with Alopecia Areata. J. Dermatol. Sci. 2021, 102, 109–115. [Google Scholar] [CrossRef]
- Lam, S.Y.; Radjabzadeh, D.; Eppinga, H.; Nossent, Y.R.A.; van der Zee, H.H.; Kraaij, R.; Konstantinov, S.R.; Fuhler, G.M.; Prens, E.P.; Thio, H.B.; et al. A Microbiome Study to Explore the Gut-Skin Axis in Hidradenitis Suppurativa. J. Dermatol. Sci. 2021, 101, 218–220. [Google Scholar] [CrossRef]
- Kazantseva, J.; Malv, E.; Kaleda, A.; Kallastu, A.; Meikas, A. Optimisation of Sample Storage and DNA Extraction for Human Gut Microbiota Studies. BMC Microbiol. 2021, 21, 158. [Google Scholar] [CrossRef]
- Milaković, M.; Križanović, S.; Petrić, I.; Šimatović, A.; González-Plaza, J.J.; Gužvinec, M.; Andrašević, A.T.; Pole, L.; Fuka, M.M.; Udiković-Kolić, N. Characterization of Macrolide Resistance in Bacteria Isolated from Macrolide-Polluted and Unpolluted River Sediments and Clinical Sources in Croatia. Sci. Total Environ. 2020, 749, 142357. [Google Scholar] [CrossRef] [PubMed]
- Panek, M.; Čipčić Paljetak, H.; Barešić, A.; Perić, M.; Matijašić, M.; Lojkić, I.; Vranešić Bender, D.; Krznarić, Ž.; Verbanac, D. Methodology Challenges in Studying Human Gut Microbiota—Effects of Collection, Storage, DNA Extraction and Next Generation Sequencing Technologies. Sci. Rep. 2018, 8, 5143. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and Gut Bacteroidetes: The Food Connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed]
- Iljazovic, A.; Roy, U.; Gálvez, E.J.C.; Lesker, T.R.; Zhao, B.; Gronow, A.; Amend, L.; Will, S.E.; Hofmann, J.D.; Pils, M.C.; et al. Perturbation of the gut microbiome by Prevotella spp. enhances host susceptibility to mucosal inflammation. Mucosal Immunol. 2021, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef]
- Hilty, M.; Burke, C.; Pedro, H.; Cardenas, P.; Bush, A.; Bossley, C.; Davies, J.; Ervine, A.; Poulter, L.; Pachter, L.; et al. Disordered microbial communities in asthmatic airways. PLoS ONE 2010, 5, e8578. [Google Scholar] [CrossRef]
- Chen, T.; Long, W.; Zhang, C.; Liu, S.; Zhao, L.; Hamaker, B.R. Fiber-Utilizing Capacity Varies in Prevotella- versus Bacteroides-Dominated Gut Microbiota. Sci. Rep. 2017, 7, 2594. [Google Scholar] [CrossRef]
- Wickens, K.; Barthow, C.; Mitchell, E.A.; Kang, J.; Van Zyl, N.; Purdie, G.; Stanley, T.; Fitzharris, P.; Murphy, R.; Crane, J. Effects of Lactobacillus Rhamnosus HN001 in Early Life on the Cumulative Prevalence of Allergic Disease to 11 Years. Pediatr. Allergy Immunol. 2018, 29, 808–814. [Google Scholar]
- Cabana, M.D.; McKean, M.; Caughey, A.B.; Fong, L.; Lynch, S.; Wong, A.; Leong, R.; Boushey, H.A.; Hilton, J.F. Early Probiotic Supplementation for Eczema and Asthma Prevention: A Randomized Controlled Trial. Pediatrics 2017, 140, e20163000. [Google Scholar] [CrossRef]
- Hol, J.; Van Leer, E.H.; Elink Schuurman, B.E.; De Ruiter, L.F.; Samsom, J.N.; Hop, W.; Neijens, H.J.; de Jongste, J.C.; Nieuwenhuis, E.E.S.; Cow’s Milk Allergy Modified by Elimination and Lactobacilli Study Group. The Acquisition of Tolerance Toward Cow’s Milk Through Probiotic Supplementation: A Randomized, Controlled Trial. J. Allergy Clin. Immunol. 2008, 121, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Rainey, F.A.; Family, V. Lachnospiraceae fam. nov. In Bergey’s Manual of Systematic Bacteriology, 3rd ed.; De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.H., Whitman, W.B., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 921–968. [Google Scholar]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A New Genomic Blueprint of the Human Gut Microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Oliphant, K.; Ali, M.; D’Souza, M.; Hughes, P.D.; Sulakhe, D.; Wang, A.Z.; Xie, B.; Yeasin, R.; Msall, M.E.; Andrews, B.; et al. Bacteroidota and Lachnospiraceae Integration into the Gut Microbiome at Key Time Points in Early Life Are Linked to Infant Neurodevelopment. Gut Microbes 2021, 13, 1997560. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.J.; Bowyer, R.C.E.; Ni Lochlainn, M.; Wells, P.M.; Roberts, H.C.; Steves, C.J. The Composition of the Gut Microbiome Differs Among Community Dwelling Older People with Good and Poor Appetite. J. Cachexia Sarcopenia Muscle 2021, 12, 368–377. [Google Scholar] [CrossRef]
- Suchodolski, J.S. Gastrointestinal Microbiota. In Canine and Feline Gastroenterology; Steiner, J.M., Ed.; Elsevier: St. Louis, MO, USA, 2013; pp. 32–41. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Devillard, E.; McIntosh, F.M.; Duncan, S.H.; Wallace, R.J. Metabolism of linoleic acid by human gut bacteria: Different routes for biosynthesis of conjugated linoleic acid. J. Bacteriol. 2007, 189, 2566–2570. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease-and uricase-containing, indole-and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Rauf, A.; Khalil, A.A.; Rahman, U.; Khalid, A.; Naz, S.; Shariati, M.A.; Rebezov, M.; Urtecho, E.Z.; Ricardo Diego Galhardo de Albuquerque, R.D.D.; Anwar, S.; et al. Recent advances in the therapeutic application of short-chain fatty acids (SCFAs): An updated review. Crit. Rev. Food Sci. Nutr. 2022, 62, 6034–6054. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, Metabolism and Microbial Ecology of Butyrate-Producing Bacteria from the Human Large Intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Hee, B.V.D.; Wells, J.M. Microbial regulation of host physiology by short-chain fatty acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [PubMed]
- Eeckhaut, V.; Machiels, K.; Perrier, C.; Romero, C.; Maes, S.; Flahou, B.; Steppe, M.; Haesebrouck, F.; Sas, B.; Ducatelle, R.; et al. Butyricicoccus pullicaecorum in Inflammatory Bowel Disease. Gut 2013, 62, 1745–1752. [Google Scholar] [CrossRef]
- Wootten, D.; Christopoulos, A.; Marti-Solano, M.; Babu, M.M.; Sexton, P.M. Mechanisms of Signalling and Biased Agonism in G Protein-Coupled Receptors. Nat. Rev. Mol. Cell Biol. 2018, 19, 638–653. [Google Scholar] [CrossRef] [PubMed]
- Krejner, A.; Bruhs, A.; Mrowietz, U.; Wehkamp, U.; Schwarz, T.; Schwarz, A. Decreased Expression of G-protein-coupled Receptors GPR43 and GPR109a in Psoriatic Skin Can be Restored by Topical Application of Sodium Butyrate. Arch. Dermatol. Res. 2018, 310, 751–758. [Google Scholar] [CrossRef]
- Schlatterer, K.; Peschel, A.; Kretschmer, D. Short-Chain Fatty Acid and FFAR2 Activation—A New Option for Treating Infections? Front. Cell. Infect. Microbiol. 2021, 11, 785833. [Google Scholar] [CrossRef]
- Marinissen, M.J.; Gutkind, J.S. G-Protein-Coupled Receptors and Signaling Networks: Emerging Paradigms. Trends Pharmacol. Sci. 2001, 22, 368–376. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Béguet-Crespel, F.; Marinelli, L.; Jamet, A.; Ledue, F.; Blottière, H.M.; Lapaque, N. Butyrate produced by gut commensal bacteria activates TGF-beta1 expression through the transcription factor SP1 in human intestinal epithelial cells. Sci. Rep. 2018, 8, 9742. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Subramanian, U.; Venkidasamy, B.; Thirupathi, P.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Chung, I.M.; Rengasamy, K.R. Emerging Role of Nutritional Short-Chain Fatty Acids (SCFAs) against Cancer via Modulation of Hematopoiesis. Crit. Rev. Food Sci. Nutr. 2021, 63, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Ćesić, D.; Lugović-Mihić, L.; Ferček, I.; Grginić, A.G.; Jelić, M.; Bešlić, I.; Tambić Andrašević, A. Salivary Microbiota Is Significantly Less Diverse in Patients with Chronic Spontaneous Urticaria Compared to Healthy Controls: Preliminary Results. Life 2021, 11, 1329. [Google Scholar] [CrossRef] [PubMed]
- Arshi, S.; Babaie, D.; Nabavi, M.; Tebianian, M.; Ghalehbaghi, B.; Jalali, F.; Ahmadvand, A.; Gholami, R. Circulating Level of CD4+ CD25+ FOXP3+ T Cells in Patients with Chronic Urticaria. Int. J. Dermatol. 2014, 53, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-C.; Chiang, B.-L.; Liu, H.E.; Leu, S.-J.; Lee, Y.-L. Defective functions of circulating CD4+CD25+ and CD4+CD25− T cells in patients with chronic ordinary urticaria. J. Dermatol. Sci. 2008, 51, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, H.; Nonaka, T.; Kishimoto, T.; Komoriya, K.; Tsuji, K.; Nakahata, T. Peroxisome Proliferator-Activated Receptors Are Expressed in Mouse Bone Marrow-Derived Mast Cells. Jpn. J. Pharmacol. 2000, 82, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, M.; Wada, K.; Katayama, K.; Kamisaki, Y.; Maeyama, K.; Kadowaki, T.; Blumberg, R.S.; Nakajima, A. Activation of Peroxisome Proliferator-Activated Receptor Gamma Suppresses Mast Cell Maturation Involved in Allergic Diseases. Allergy 2008, 63, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Folkerts, J.; Redegeld, F.; Folkerts, G.; Blokhuis, B.; van den Berg, M.P.M.; de Bruijn, M.J.W.; van IJcken, W.F.; Junt, T.; Tam, S.Y.; Galli, S.J.; et al. Butyrate Inhibits Human Mast Cell Activation via Epigenetic Regulation of FcεRI-Mediated Signaling. Allergy 2020, 75, 1966–1978. [Google Scholar] [CrossRef]
- Kasahara, K.; Krautkramer, K.A.; Org, E.; Romano, K.A.; Kerby, R.L.; Vivas, E.I.; Mehrabian, M.; Denu, J.M.; Bäckhed, F.; Lusis, A.J.; et al. Interactions between Roseburia intestinalis and Diet Modulate Atherogenesis in a Murine Model. Nat. Microbiol. 2018, 3, 1461. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | CSU Patients (N = 22) | Healthy Controls (N = 23) | p-Value |
|---|---|---|---|
| Median age, years (range) | 42 (20–73) | 40 (19–74) | 0.928 |
| Sex, n (%) | |||
| Male | 6 (27.3%) | 7 (30.4%) | 0.817 |
| Female | 16 (72.7%) | 16 (69.6%) |
| Variable | CSU Patients (N = 22) |
|---|---|
| Duration of symptoms | |
| 6 weeks–5 months, n (%) | 15 (68%) |
| 6 months–12 months, n (%) | 7 (32%) |
| Appearance of urticaria | |
| everyday, n (%) | 13 (59%) |
| 2–4 times a week, n (%) | 8 (36%) |
| once a week, n (%) | 1 (5%) |
| Taking non-sedating antihistamines n (%) | 20 (91%) |
| 1 tablet daily, n (%) | 5 (23%) |
| 2 tablets daily, n (%) | 8 (36%) |
| 3 tablets daily, n (%) | 3 (14%) |
| 4 tablets daily, n (%) | 4 (18%) |
| Variable | CSU Patients (N = 22) |
|---|---|
| Associated angioedema, n (%) | 7 (32%) |
| Associated atopic disorders, n (%) | 7 (32%) |
| Elevated Anti-Tg, n (%) | 6 (27%) |
| Elevated TPO-Ab, n (%) | 11 (50%) |
| Vitamin D deficiency, n (%) | 15(68%) |
| Elevated IgE, n (%) | 9 (41%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ćesić, D.; Lugović Mihić, L.; Ozretić, P.; Lojkić, I.; Buljan, M.; Šitum, M.; Zovak, M.; Vidović, D.; Mijić, A.; Galić, N.; et al. Association of Gut Lachnospiraceae and Chronic Spontaneous Urticaria. Life 2023, 13, 1280. https://doi.org/10.3390/life13061280
Ćesić D, Lugović Mihić L, Ozretić P, Lojkić I, Buljan M, Šitum M, Zovak M, Vidović D, Mijić A, Galić N, et al. Association of Gut Lachnospiraceae and Chronic Spontaneous Urticaria. Life. 2023; 13(6):1280. https://doi.org/10.3390/life13061280
Chicago/Turabian StyleĆesić, Diana, Liborija Lugović Mihić, Petar Ozretić, Ivana Lojkić, Marija Buljan, Mirna Šitum, Mario Zovak, Dinko Vidović, August Mijić, Nada Galić, and et al. 2023. "Association of Gut Lachnospiraceae and Chronic Spontaneous Urticaria" Life 13, no. 6: 1280. https://doi.org/10.3390/life13061280
APA StyleĆesić, D., Lugović Mihić, L., Ozretić, P., Lojkić, I., Buljan, M., Šitum, M., Zovak, M., Vidović, D., Mijić, A., Galić, N., & Tambić Andrašević, A. (2023). Association of Gut Lachnospiraceae and Chronic Spontaneous Urticaria. Life, 13(6), 1280. https://doi.org/10.3390/life13061280