
Phase-Amplitude Coupling Localizes Pathologic Brain with Aid of Behavioral Staging in Sleep


{kind=link}
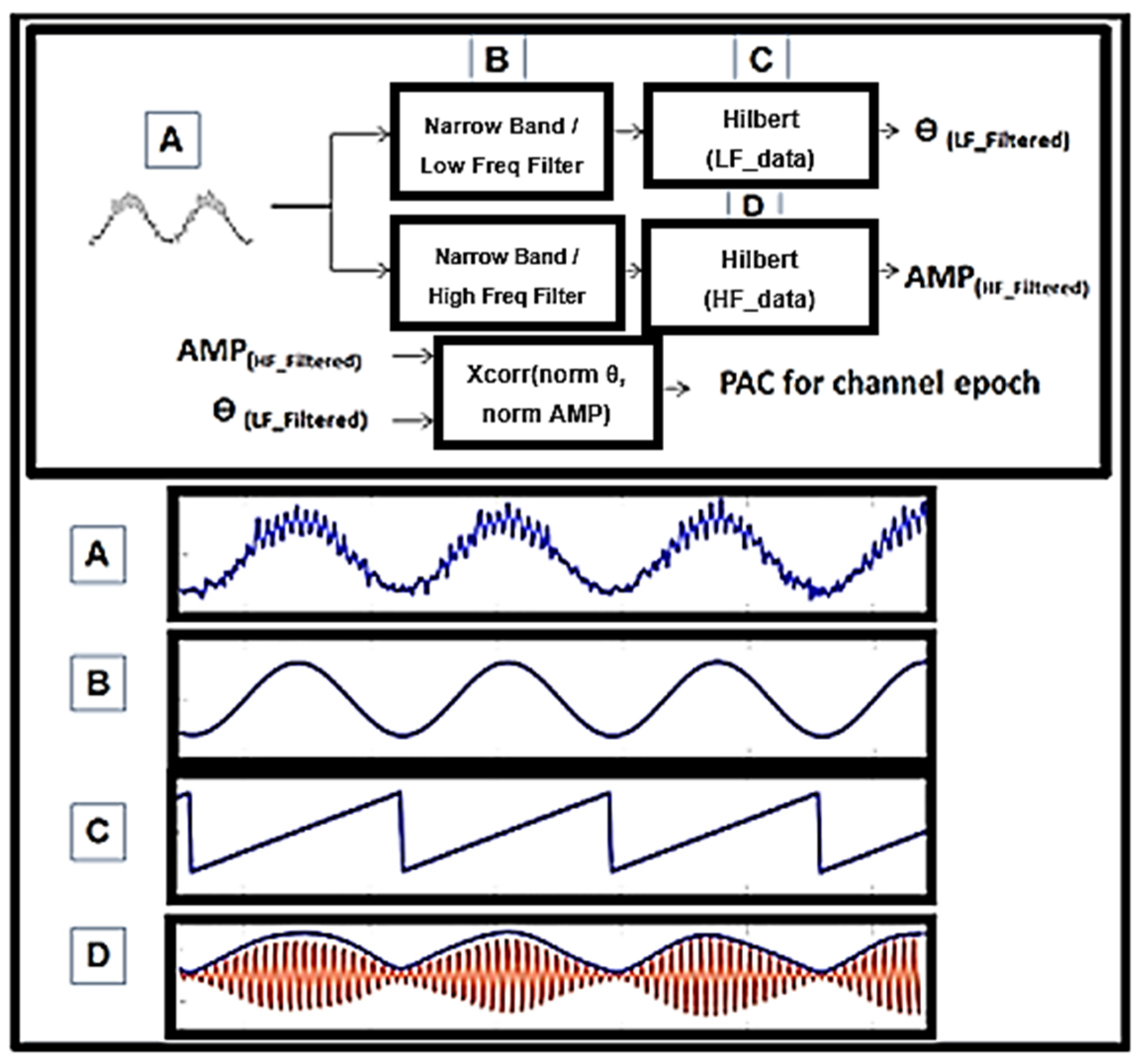{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Data Acquisition
2.3. Contact Placement and Co-Registered Localization
2.3.1. Artifact Identification and Night Segment Data Choice
2.3.2. Interictal Epileptiform Discharge and High Frequency Oscillation, and Phase-Amplitude Coupling Identification
2.3.3. Seizure Onset Zone
2.3.4. Sleep Scoring with Scalp EEG
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A. Phase-Amplitude Coupling
References
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; Engel, J., Jr.; Forsgren, L.; French, J.A.; Glynn, M.; et al. ILAE official report: A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.N.; Filippi, D.; Hauser, W.A. The Descriptive Epidemiology of Epilepsy-a Review. Epilepsy Res. 2009, 85, 31–45. [Google Scholar] [CrossRef]
- Geertsema, E.E.; Visser, G.H.; Velis, D.N.; Claus, S.P.; Zijlmans, M.; Kalitzin, S. Automated Seizure Onset Zone Approximation Based on Nonharmonic High-Frequency Oscillations in Human Interictal Intracranial EEGs. Int. J. Neural Syst. 2015, 25, 1550015. [Google Scholar] [CrossRef] [PubMed]
- Kremen, V.; Duque, J.J.; Brinkmann, B.H.; Berry, B.M.; Kucewicz, M.T.; Khadjevand, F.; Van Gompel, J.; Stead, M.; Louis, E.K.S.; Worrell, G.A. Behavioral State Classification in Epileptic Brain Using Intracranial Electrophysiology. J. Neural Eng. 2017, 14, 26001. [Google Scholar] [CrossRef] [PubMed]
- Novikov, E.; Novikov, A.; Shannahoff-Khalsa, D.; Schwartz, B.; Wright, J. Scale-similar activity in the brain. Phys. Rev. E 1997, 56, R2387–R2389. [Google Scholar] [CrossRef]
- Saboo, K.V.; Varatharajah, Y.; Berry, B.M.; Kremen, V.; Sperling, M.R.; Davis, K.A.; Jobst, B.C.; Gross, R.E.; Lega, B.; Sheth, S.A.; et al. Unsupervised machine-learning classification of electrophysiologically active electrodes during human cognitive task performance. Sci. Rep. 2019, 9, 17390. [Google Scholar] [CrossRef]
- Wagh, N.; Wei, J.; Rawal, S.; Berry, B.M.; Barnard, L.; Brinkmann, B.H.; Worrell, G.A.; Jones, D.T.; Varatharajah, Y. Domain-guided Self-supervision of EEG Data Improves Downstream Classification Performance and Generalizability. In Proceedings of the Machine Learning Research (PMLR), Virtual Event, 6–7 August 2021. [Google Scholar]
- Varatharajah, Y.; Chong, M.J.; Saboo, K.V.; Berry, B.M.; Brinkmann, B.H.; Worrell, G.A.; Iyer, R.K. EEG-GRAPH: A Factor-Graph-Based Model for Capturing Spatial, Temporal, and Observational Relationships in Electroencephalograms; NIPS: Denver, CO, USA, 2017. [Google Scholar]
- Varatharajah, Y.; Berry, B.; Joseph, B.; Balzekas, I.; Attia, T.P.; Kremen, V.; Brinkmann, B.; Iyer, R.; Worrell, G. Characterizing the electrophysiological abnormalities in visually reviewed normal EEGs of drug-resistant focal epilepsy patients. Brain Commun. 2021, 3, fcab102. [Google Scholar] [CrossRef]
- Matsumoto, A.; Brinkmann, B.H.; Stead, S.M.; Matsumoto, J.; Kucewicz, M.T.; Marsh, W.R.; Meyer, F.; Worrell, G. Pathological and Physiological High-Frequency Oscillations in Focal Human Epilepsy. J. Neurophysiol. 2013, 110, 1958–1964. [Google Scholar] [CrossRef]
- Burns, S.P.; Santaniello, S.; Yaffe, R.B.; Jouny, C.C.; Crone, N.E.; Bergey, G.K.; Anderson, W.S.; Sarma, S.V. Network dynamics of the brain and influence of the epileptic seizure onset zone. Proc. Natl. Acad. Sci. USA 2014, 111, E5321–E5330. [Google Scholar] [CrossRef]
- Alvarado-Rojas, C.; Valderrama, M.; Fouad-Ahmed, A.; Feldwisch-Drentrup, H.; Ihle, M.; Teixeira, C.A.; Sales, F.; Schulze-Bonhage, A.; Adam, C.; Dourado, A.; et al. Slow Modulations of High-Frequency Activity (40–140 Hz) Discriminate Preictal Changes in Human Focal Epilepsy. Sci. Rep. 2014, 4, 4545. [Google Scholar] [CrossRef]
- Edakawa, K.; Yanagisawa, T.; Kishima, H.; Fukuma, R.; Oshino, S.; Khoo, H.M.; Kobayashi, M.; Tanaka, M.; Yoshimine, T. Detection of Epileptic Seizures Using Phase–Amplitude Coupling in Intracranial Electroencephalography. Sci. Rep. 2016, 6, 25422. [Google Scholar] [CrossRef]
- Kucewicz, M.T.; Berry, B.M.; Miller, L.R.; Khadjevand, F.; Ezzyat, Y.; Stein, J.M.; Kremen, V.; Brinkmann, B.H.; Wanda, P.; Sperling, M.R.; et al. Dissecting Gamma Frequency Activity during Human Memory Processing. Brain 2017, 140, 1337–1350. [Google Scholar] [CrossRef]
- Kucewicz, M.T.; Cimbalnik, J.; Matsumoto, J.Y.; Brinkmann, B.H.; Bower, M.R.; Vasoli, V.; Sulc, V.; Meyer, F.; Marsh, W.R.; Stead, S.M.; et al. High Frequency Oscillations Are Associated with Cognitive Processing in Human Recognition Memory. Brain A J. Neurol. 2014, 137 Pt 8, 2231–2244. [Google Scholar] [CrossRef]
- Kucewicz, M.T.; Berry, B.M.; Bower, M.R.; Cimbalnik, J.; Svehlik, V.; Stead, S.M.; Worrell, G.A. Combined Single Neuron Unit Activity and Local Field Potential Oscillations in a Human Visual Recognition Memory Task. IEEE Trans. Biomed. Eng. 2016, 63, 67–75. [Google Scholar] [CrossRef]
- Weiss, S.A.; Lemesiou, A.; Connors, R.; Banks, G.P.; McKhann, G.M.; Goodman, R.R.; Zhao, B.; Filippi, C.G.; Nowell, M.; Rodionov, R.; et al. Seizure Localization Using Ictal Phase-Locked High Gamma: A Retrospective Surgical Outcome Study. Neurology 2015, 84, 2320–2328. [Google Scholar] [CrossRef]
- Weiss, S.A.; Orosz, I.; Salamon, N.; Moy, S.; Wei, L.; Klooster, M.A.V.; Knight, R.T.; Harper, R.M.; Bragin, A.; Fried, I.; et al. Ripples on Spikes Show Increased Phase-Amplitude Coupling in Mesial Temporal Lobe Epilepsy Seizure-Onset Zones. Epilepsia 2016, 57, 1916–1930. [Google Scholar] [CrossRef]
- Canolty, R.T.; Edwards, E.; Dalal, S.S.; Soltani, M.; Nagarajan, S.S.; Kirsch, H.E.; Berger, M.S.; Barbaro, N.M.; Knight, R.T. High Gamma Power Is Phase-Locked to Theta Oscillations in Human Neocortex. Science 2006, 313, 1626–1628. [Google Scholar] [CrossRef]
- Jacobs, J.; Vogt, C.; LeVan, P.; Zelmann, R.; Gotman, J.; Kobayashi, K. The Identification of Distinct High-Frequency Oscillations during Spikes Delineates the Seizure Onset Zone Better than High-Frequency Spectral Power Changes. Clin. Neurophysiol. 2016, 127, 129–142. [Google Scholar] [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Linkenkaer-Hansen, K.; Nikouline, V.V.; Palva, J.M.; Ilmoniemi, R.J. Long-range temporal correlations and scaling behavior in human brain oscillations. J. Neurosci. 2001, 21, 1370–1377. [Google Scholar] [CrossRef]
- Vanhatalo, S.; Palva, J.M.; Holmes, M.D.; Miller, J.W.; Voipio, J.; Kaila, K. Infraslow Oscillations Modulate Excitability and Interictal Epileptic Activity in the Human Cortex during Sleep. Proc. Natl. Acad. Sci. USA 2004, 101, 5053–5057. [Google Scholar] [CrossRef] [PubMed]
- Monto, S.; Palva, S.; Voipio, J.; Palva, J.M. Very slow EEG fluctuations predict the dynamics of stimulus detection and oscillation amplitudes in humans. J. Neurosci. 2008, 28, 8268–8272. [Google Scholar] [CrossRef] [PubMed]
- Canolty, R.T.; Knight, R.T. The Functional Role of Cross-Frequency Coupling. Trends Cogn. Sci. 2010, 14, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Osipova, D.; Hermes, D.; Jensen, O. Gamma Power Is Phase-Locked to Posterior Alpha Activity. PLoS ONE 2008, 3, e3990. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Frauscher, B.; Gotman, J. Phase-Amplitude Coupling Is Elevated in Deep Sleep and in the Onset Zone of Focal Epileptic Seizures. Front. Hum. Neurosci. 2016, 10, 387. [Google Scholar] [CrossRef]
- Mihály, I.; Orbán-Kis, K.; Gáll, Z.; Berki, Á.-J.; Bod, R.-B.; Szilágyi, T. Amygdala Low-Frequency Stimulation Reduces Pathological Phase-Amplitude Coupling in the Pilocarpine Model of Epilepsy. Brain Sci. 2020, 10, 856. [Google Scholar] [CrossRef]
- Pasquetti, M.V.; Meier, L.; Marafiga, J.R.; Caus, L.B.; Tort, A.B.L.; Calcagnotto, M.E. Hippocampal CA1 and cortical interictal oscillations in the pilocarpine model of epilepsy. Brain Res. 2019, 1722, 146351. [Google Scholar] [CrossRef]
- Samiee, S.; Lévesque, M.; Avoli, M.; Baillet, S. Phase-amplitude coupling and epileptogenesis in an animal model of mesial temporal lobe epilepsy. Neurobiol. Dis. 2018, 114, 111–119. [Google Scholar] [CrossRef]
- Guirgis, M.; Chinvarun, Y.; del Campo, M.; Carlen, P.L.; Bardakjian, B.L. Defining Regions of Interest Using Cross-Frequency Coupling in Extratemporal Lobe Epilepsy Patients. J. Neural Eng. 2015, 12, 26011. [Google Scholar] [CrossRef]
- Ibrahim, G.M.; Wong, S.M.; Anderson, R.A.; Singh-Cadieux, G.; Akiyama, T.; Ochi, A.; Otsubo, H.; Okanishi, T.; Valiante, T.A.; Donner, E.; et al. Dynamic Modulation of Epileptic High Frequency Oscillations by the Phase of Slower Cortical Rhythms. Exp. Neurol. 2014, 251, 30–38. [Google Scholar] [CrossRef]
- Maris, E.; van Vugt, M.; Kahana, M. Spatially Distributed Patterns of Oscillatory Coupling between High-Frequency Amplitudes and Low-Frequency Phases in Human iEEG. NeuroImage 2011, 54, 836–850. [Google Scholar] [CrossRef]
- Staresina, B.P.; Bergmann, T.O.; Bonnefond, M.; Van Der Meij, R.; Jensen, O.; Deuker, L.; Elger, C.E.; Axmacher, N.; Fell, J. Hierarchical nesting of slow oscillations, spindles and ripples in the human hippocampus during sleep. Nat. Neurosci. 2015, 18, 1679–1686. [Google Scholar] [CrossRef]
- Achermann, P.; Borbély, A.A. Low-frequency (< 1 Hz) oscillations in the human sleep electroencephalogram. Neuroscience 1997, 81, 213–222. [Google Scholar]
- Steriade, M.; Nunez, A. Oscillatory Processes in the Brain; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Sirota, A.; Csicsvari, J.; Buhl, D.; Buzsáki, G. Communication between neocortex and hippocampus during sleep in rodents. Proc. Natl. Acad. Sci. USA 2003, 100, 2065–2069. [Google Scholar] [CrossRef]
- Andrillon, T.; Nir, Y.; Staba, R.J.; Ferrarelli, F.; Cirelli, C.; Tononi, G.; Fried, I. Sleep spindles in humans: Insights from intracranial EEG and unit recordings. J. Neurosci. 2011, 31, 17821–17834. [Google Scholar] [CrossRef]
- Roebber, J.K.; Lewis, P.A.; Crunelli, V.; Navarrete, M.; Hamandi, K. Effects of Anti-Seizure Medication on Sleep Spindles and Slow Waves in Drug-Resistant Epilepsy. Brain Sci. 2022, 12, 1288. [Google Scholar] [CrossRef]
- Gouveris, H.; Koirala, N.; Anwar, A.R.; Ding, H.; Ludwig, K.; Huppertz, T.; Matthias, C.; Groppa, S.; Muthuraman, M. Reduced Cross-Frequency Coupling and Daytime Sleepiness in Obstructive Sleep Apnea Patients. Biology 2022, 11, 700. [Google Scholar] [CrossRef]
- Diekelmann, S.; Born, J. The Memory Function of Sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef]
- Varatharajah, Y.; Berry, B.M.; Kalbarczyk, Z.T.; Brinkmann, B.H.; Worrell, G.A.; Iyer, R.K. Inter-ictal Seizure Onset Zone localization using unsupervised clustering and Bayesian Filtering. In Proceedings of the 2017 8th International IEEE/EMBS Conference on Neural Engineering (NER), Shanghai, China, 25–28 May 2017; pp. 533–539. [Google Scholar]
- Thompson, G.J.; Pan, W.-J.; Billings, J.C.W.; Grooms, J.K.; Shakil, S.; Jaeger, D.; Keilholz, S.D. Phase-Amplitude Coupling and Infraslow. Front. Integr. Neurosci. 2014, 8, 41. [Google Scholar] [CrossRef]
- Thompson, S.A.; Krishnan, B.; Gonzalez-Martinez, J.; Bulacio, J.; Jehi, L.; Mosher, J.; Alexopoulos, A.; Burgess, R.C. Interictal Infraslow Activity in Stereoelectroencephalography. J. Clin. Neurophysiol. 2016, 33, 141–148. [Google Scholar] [CrossRef]
- Ren, L.; Kucewicz, M.T.; Cimbalnik, J.; Matsumoto, J.Y.; Brinkmann, B.H.; Hu, W.; Marsh, W.R.; Meyer, F.B.; Stead, S.M.; Worrell, G.A. Gamma Oscillations Precede Interictal Epileptiform Spikes in the Seizure Onset Zone. Neurology 2015, 84, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Bower, M.R.; Stead, M.; Bower, R.S.; Kucewicz, M.T.; Sulc, V.; Cimbalnik, J.; Brinkmann, B.H.; Vasoli, V.M.; Louis, E.K.S.; Meyer, F.B.; et al. Evidence for Consolidation of Neuronal Assemblies after Seizures in Humans. J. Neurosci. 2015, 35, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, B.H.; Bower, M.R.; Stengel, K.A.; Worrell, G.A.; Stead, M. Large-scale electrophysiology: Acquisition, compression, encryption, and storage of big data. J. Neurosci. Methods 2009, 180, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Barkmeier, D.T.; Shah, A.K.; Flanagan, D.; Atkinson, M.D.; Agarwal, R.; Fuerst, D.R.; Jafari-Khouzani, K.; Loeb, J.A. High Inter-Reviewer Variability of Spike Detection on Intracranial EEG Addressed by an Automated Multi-Channel Algorithm. Clin. Neurophysiol. 2012, 123, 1088–1095. [Google Scholar] [CrossRef]
- Cimbalnik, J.; Kucewicz, M.; Worrell, G. Interictal High-Frequency Oscillations in Focal Human Epilepsy. Curr. Opin. Neurol. 2016, 29, 175–181. [Google Scholar] [CrossRef]
- Jordan, D.; Ilg, R.; Riedl, V.; Schorer, A.; Grimberg, S.; Neufang, S.; Omerovic, A.; Berger, S.; Untergehrer, G.; Preibisch, C.; et al. Simultaneous electroencephalographic and functional magnetic resonance imaging indicate impaired cortical top-down processing in association with anesthetic-induced unconsciousness. Anesthesiology 2013, 119, 1031–1042. [Google Scholar] [CrossRef]
- Bagshaw, A.P.; Jacobs, J.; LeVan, P.; Dubeau, F.; Gotman, J. Effect of sleep stage on interictal high-frequency oscillations recorded from depth macroelectrodes in patients with focal epilepsy. Epilepsia 2009, 50, 617–628. [Google Scholar] [CrossRef]
- Valencia, M.; Artieda, J.; Bolam, J.P.; Mena-Segovia, J. Dynamic interaction of spindles and gamma activity during cortical slow oscillations and its modulation by subcortical afferents. PLoS ONE 2013, 8, e67540. [Google Scholar] [CrossRef]
- Demuru, M.; Kalitzin, S.; Zweiphenning, W.; van Blooijs, D.; Klooster, M.V.; Van Eijsden, P.; Leijten,, F.; Zijlmans, M.; Braun, K.; Ferrier, C.; et al. The value of intra-operative electrographic biomarkers for tailoring during epilepsy surgery: From group-level to patient-level analysis. Sci. Rep. 2020, 10, 14654. [Google Scholar] [CrossRef]
- Cámpora, N.E.; Mininni, C.J.; Kochen, S.; Lew, S.E. Seizure localization using pre ictal phase-amplitude coupling in intracranial electroencephalography. Sci. Rep. 2019, 9, 20022. [Google Scholar] [CrossRef]
- Bandarabadi, M.; Gast, H.; Rummel, C.; Bassetti, C.; Adamantidis, A.; Schindler, K.; Zubler, F. Assessing Epileptogenicity Using Phase-Locked High Frequency Oscillations: A Systematic Comparison of Methods. Front. Neurol. 2019, 10, 1132. [Google Scholar] [CrossRef]
- Mukamel, E.A.; Wong, K.F.; Prerau, M.J.; Brown, E.N.; Purdon, P.L. Phase-Based Measures of Cross-Frequency Coupling in Brain Electrical Dynamics under General Anesthesia. In Proceedings of the 2011 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Boston, MA, USA, 30 August–3 September 2011; pp. 1981–1984. [Google Scholar] [CrossRef]
- Mukamel, E.A.; Pirondini, E.; Babadi, B.; Wong, K.F.K.; Pierce, E.T.; Harrell, P.G.; Walsh, J.L.; Salazar-Gomez, A.F.; Cash, S.S.; Eskandar, E.N.; et al. A Transition in Brain State during Propofol-Induced Unconsciousness. J. Neurosci. 2014, 34, 839–845. [Google Scholar] [CrossRef]
- Geertsema, E.E.; Klooster, M.A.V.; van Klink, N.E.; Leijten, F.S.; van Rijen, P.C.; Visser, G.H.; Kalitzin, S.N.; Zijlmans, M. Non-Harmonicity in High-Frequency Components of the Intra-Operative Corticogram to Delineate Epileptogenic Tissue during Surgery. Clin. Neurophysiol. 2017, 128, 153–164. [Google Scholar] [CrossRef]
- Guragain, H.; Cimbalnik, J.; Stead, M.; Groppe, D.M.; Berry, B.M.; Kremen, V.; Kenney-Jung, D.; Britton, J.; Worrell, G.A.; Brinkmann, B.H. Spatial variation in high-frequency oscillation rates and amplitudes in intracranial EEG. Neurology 2018, 90, e639–e646. [Google Scholar] [CrossRef]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.; Quan, S.F. For the American Academy of Sleep Medicine. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications; American Academy of Sleep Medicine: Westchester, IL, USA, 2007. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berry, B.; Varatharajah, Y.; Kremen, V.; Kucewicz, M.; Guragain, H.; Brinkmann, B.; Duque, J.; Carvalho, D.Z.; Stead, M.; Sieck, G.; et al. Phase-Amplitude Coupling Localizes Pathologic Brain with Aid of Behavioral Staging in Sleep. Life 2023, 13, 1186. https://doi.org/10.3390/life13051186
Berry B, Varatharajah Y, Kremen V, Kucewicz M, Guragain H, Brinkmann B, Duque J, Carvalho DZ, Stead M, Sieck G, et al. Phase-Amplitude Coupling Localizes Pathologic Brain with Aid of Behavioral Staging in Sleep. Life. 2023; 13(5):1186. https://doi.org/10.3390/life13051186
Chicago/Turabian StyleBerry, Brent, Yogatheesan Varatharajah, Vaclav Kremen, Michal Kucewicz, Hari Guragain, Benjamin Brinkmann, Juliano Duque, Diego Z. Carvalho, Matt Stead, Gary Sieck, and et al. 2023. "Phase-Amplitude Coupling Localizes Pathologic Brain with Aid of Behavioral Staging in Sleep" Life 13, no. 5: 1186. https://doi.org/10.3390/life13051186
APA StyleBerry, B., Varatharajah, Y., Kremen, V., Kucewicz, M., Guragain, H., Brinkmann, B., Duque, J., Carvalho, D. Z., Stead, M., Sieck, G., & Worrell, G. (2023). Phase-Amplitude Coupling Localizes Pathologic Brain with Aid of Behavioral Staging in Sleep. Life, 13(5), 1186. https://doi.org/10.3390/life13051186