

Gut Microbiota-Derived Short-Chain Fatty Acids: Novel Regulators of Intestinal Serotonin Transporter

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. 5-HT Uptake Studies
2.3. RNA Isolation, Reverse Transcription and Real-Time PCR
2.4. Protein Analysis by Western Blotting
2.5. Statistical Analysis
3. Results
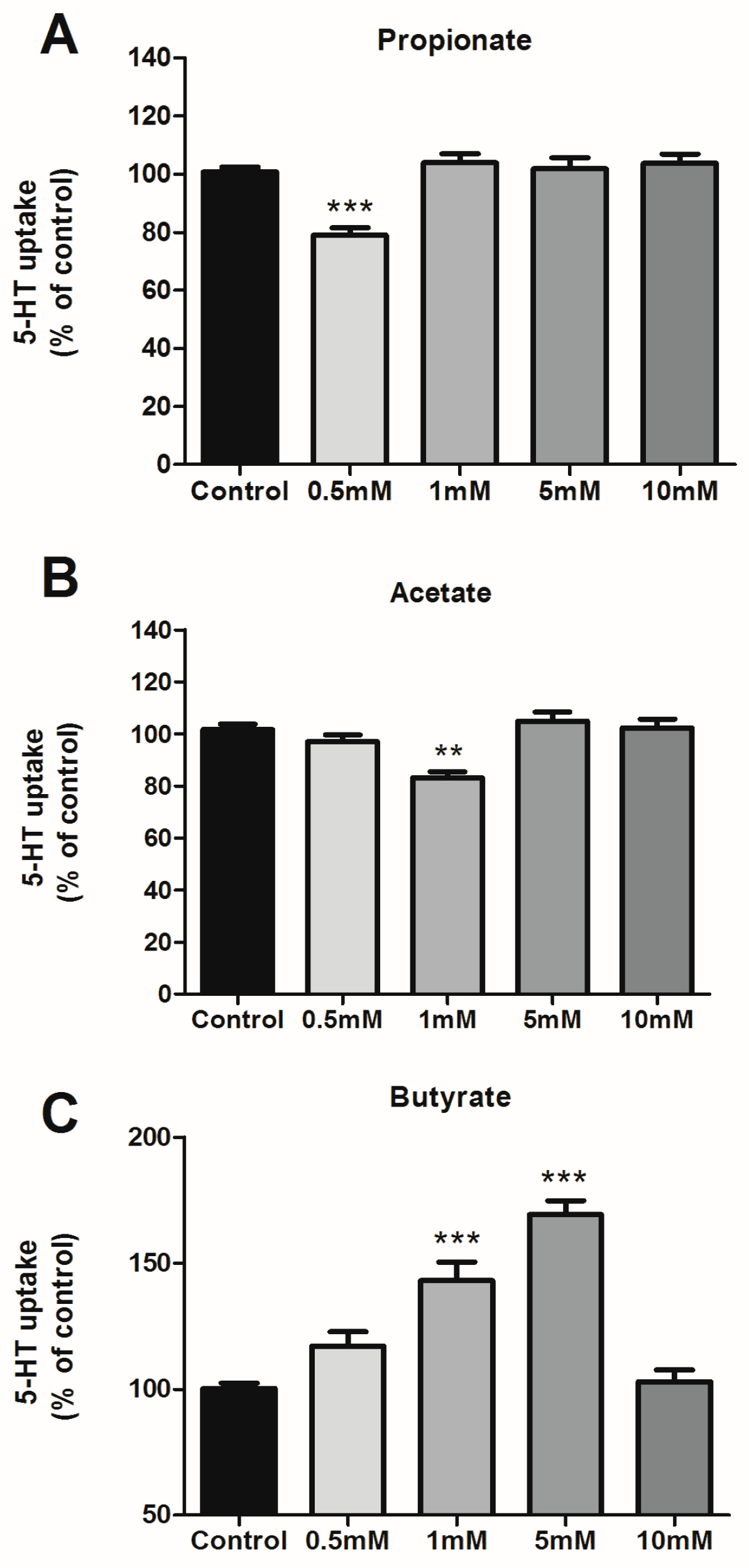
3.1. Effects of SCFAs on 5-HT Uptake
3.2. SERT Expression Is Regulated by SCFAs
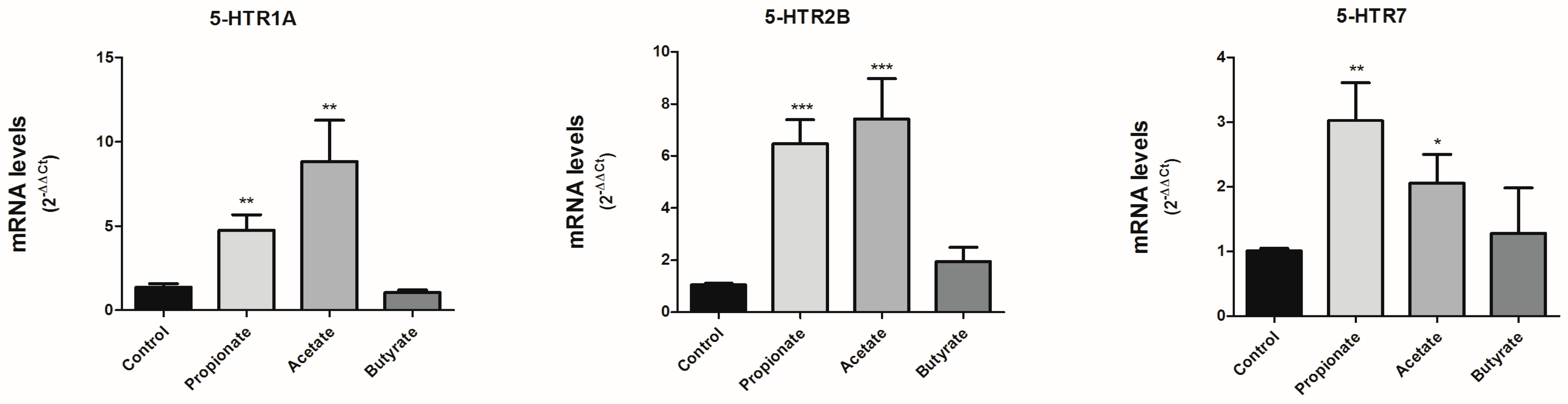
3.3. SCFAs Modulate the Expression of 5-HT Receptors
3.4. Effects of Physiological SCFAs Mix on Serotoninergic System
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.; Fernandes, G.; Tap, J.; Bruls, T.; Batto, J.; et al. Enterotypes of the Human Gut Microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, K.J.; Collins, M.K.; Moloney, G.M.; Knox, E.G.; Aburto, M.R.; Fülling, C.; Morley, S.J.; Clarke, G.; Schellekens, H.; Cryan, J.F. Short Chain Fatty Acids: Microbial Metabolites for Gut-Brain Axis Signalling. Mol. Cell. Endocrinol. 2022, 546, 111572:1–111572:18. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Campos-Perez, W.; Martinez-Lopez, E. Effects of Short Chain Fatty Acids on Metabolic and Inflammatory Processes in Human Health. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2021, 1866, 158900:1–158900:8. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The Role of Short-Chain Fatty Acids in Immunity, Inflammation and Metabolism. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–12. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Microbes Immunity and Behavior: Psychoneuroimmunology Meets the Microbiome. Neuropsychopharmacology 2017, 42, 178–192. [Google Scholar] [CrossRef]
- Latorre, E.; Mesonero, J.E.; Harries, L.W. Alternative Splicing in Serotonergic System: Implications in Neuropsychiatric Disorders. J. Psychopharmacol. 2019, 33, 1352–1363. [Google Scholar] [CrossRef]
- Layunta, E.; Latorre, E.; Forcén, R.; Grasa, L.; Plaza, M.A.; Arias, M.; Alcalde, A.I.; Mesonero, J.E. NOD1 Downregulates Intestinal Serotonin Transporter and Interacts with Other Pattern Recognition Receptors. J. Cell. Physiol. 2018, 233, 4183–4193. [Google Scholar] [CrossRef]
- Layunta, E.; Buey, B.; Mesonero, J.E.; Latorre, E. Crosstalk Between Intestinal Serotonergic System and Pattern Recognition Receptors on the Microbiota–Gut–Brain Axis. Front. Endocrinol. 2021, 12, 748254:1–748254:24. [Google Scholar] [CrossRef]
- Layunta, E.; Latorre, E.; Grasa, L.; Arruebo, M.P.; Buey, B.; Alcalde, A.I.; Mesonero, J.E. Intestinal Serotonergic System Is Modulated by Toll-like Receptor 9. J. Physiol. Biochem. 2022, 78, 689–701. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F.; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut Microbes Promote Colonic Serotonin Production through an Effect of Short-Chain Fatty Acids on Enterochromaffin Cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13:1–13:22. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef]
- Iceta, R.; Mesonero, J.E.; Aramayona, J.J.; Alcalde, A.I. Expression of 5-HT1A and 5-HT7 Receptors in Caco-2 Cells and Their Role in the Regulation of Serotonin Transporter Activity. J. Physiol. Pharmacol. 2009, 60, 157–164. [Google Scholar]
- Iceta, R.; Aramayona, J.J.; Mesonero, J.E.; Alcalde, A.I. Regulation of the Human Serotonin Transporter Mediated by Long-Term Action of Serotonin in Caco-2 Cells. Acta Physiol. 2008, 193, 57–65. [Google Scholar] [CrossRef]
- D’Argenio, G.; Mazzacca, G. Short-Chain Fatty Acid in the Human Colon: Relation to Inflammatory Bowel Diseases and Colon Cancer. In Advances in Experimental Medicine and Biology; Zappia, V., DellaRagione, F., Barbarisi, A., Russo, G., Dellolacovo, R., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; Volume 472, pp. 149–158. ISBN 0-306-46306-46307. [Google Scholar]
- Banskota, S.; Ghia, J.E.; Khan, W.I. Serotonin in the gut: Blessing or a curse. Biochimie 2019, 161, 56–64. [Google Scholar] [CrossRef]
- Yano, J.; Yu, K.; Donaldson, G.; Shastri, G.; Ann, P.; Ma, L.; Nagler, C.; Ismagilov, R.; Mazmanian, S.; Hsiao, E. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Ye, L.; Bae, M.; Cassilly, C.D.; Jabba, S.V.; Thorpe, D.W.; Martin, A.M.; Lu, H.-Y.; Wang, J.; Thompson, J.D.; Lickwar, C.R.; et al. Enteroendocrine Cells Sense Bacterial Tryptophan Catabolites to Activate Enteric and Vagal Neuronal Pathways. Cell Host Microbe 2021, 29, 179–196. [Google Scholar] [CrossRef]
- Latorre, E.; Mendoza, C.; Matheus, N.; Castro, M.; Grasa, L.; Mesonero, J.E.; Alcalde, A.I. IL-10 Modulates Serotonin Transporter Activity and Molecular Expression in Intestinal Epithelial Cells. Cytokine 2013, 61, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, H.; Lin, Y.; Wang, G.; Luo, Y.; Li, X.; Wang, M.; Huai, M.; Li, L.; Barri, A. Butyrate Glycerides Protect against Intestinal Inflammation and Barrier Dysfunction in Mice. Nutrients 2022, 14, 3991. [Google Scholar] [CrossRef] [PubMed]
- Jørandli, J.W.; Thorsvik, S.; Skovdahl, H.K.; Kornfeld, B.; Sæterstad, S.; Gustafsson, B.I.; Sandvik, A.K.; Van Beelen Granlund, A. The Serotonin Reuptake Transporter Is Reduced in the Epithelium of Active Crohn’s Disease and Ulcerative Colitis. Am. J. Physiol.—Gastrointest. Liver Physiol. 2020, 319, G761–G768. [Google Scholar] [CrossRef] [PubMed]
- Sharif, M.; Chauhan, U.; Adeeb, S.; Chetty, Y.; Armstrong, D.; Halder, S.; Marshall, J.; Khan, W. Characterization of Serotonin Signaling Components in Patients with Inflammatory Bowel Disease. J. Can. Assoc. Gastroenterol. 2019, 2, 132–140. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979:1–979:21. [Google Scholar] [CrossRef]
- Gasaly, N.; Hermoso, M.A.; Gotteland, M. Butyrate and the Fine-Tuning of Colonic Homeostasis: Implication for Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 3061. [Google Scholar] [CrossRef]
- Venegas, D.P.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277:1–277:16. [Google Scholar] [CrossRef]
- Engevik, M.; Ruan, W.; Visuthranukul, C.; Shi, Z.; Engevik, K.A.; Engevik, A.C.; Fultz, R.; Schady, D.A.; Spinler, J.K.; Versalovic, J. Limosilactobacillus Reuteri ATCC 6475 Metabolites Upregulate the Serotonin Transporter in the Intestinal Epithelium. Benef. Microbes 2021, 12, 583–599. [Google Scholar] [CrossRef]
- Carrard, A.; Elsayed, M.; Margineanu, M.; Boury-Jamot, B.; Fragnière, L.; Meylan, E.M.; Petit, J.M.; Fiumelli, H.; Magistretti, P.J.; Martin, J.L. Peripheral Administration of Lactate Produces Antidepressant-like Effects. Mol. Psychiatry 2018, 23, 392–399. [Google Scholar] [CrossRef]
- Diaz, S.L.; Doly, S.; Narboux-Nme, N.; Fernández, S.; Mazot, P.; Banas, S.M.; Boutourlinsky, K.; Moutkine, I.; Belmer, A.; Roumier, A.; et al. 5-HT 2B Receptors Are Required for Serotonin-Selective Antidepressant Actions. Mol. Psychiatry 2012, 17, 154–163. [Google Scholar] [CrossRef]
- Chang, W.Y.; Yang, Y.T.; She, M.P.; Tu, C.H.; Lee, T.C.; Wu, M.S.; Sun, C.H.; Hsin, L.W.; Yu, L.C.H. 5-HT7 Receptor-Dependent Intestinal Neurite Outgrowth Contributes to Visceral Hypersensitivity in Irritable Bowel Syndrome. Lab. Investig. 2022, 102, 1023–1037. [Google Scholar] [CrossRef]
- Wang, A.; Si, H.; Liu, D.; Jiang, H. Butyrate Activates the CAMP-Protein Kinase A-CAMP Response Element-Binding Protein Signaling Pathway in Caco-2 Cells. J. Nutr. 2012, 142, 1–6. [Google Scholar] [CrossRef]
- Liu, L.; Fu, C.; Li, F. Acetate Affects the Process of Lipid Metabolism in Rabbit Liver, Skeletal Muscle and Adipose Tissue. Animals 2019, 9, 799. [Google Scholar] [CrossRef]
- Park, B.O.; Kim, S.H.; Kong, G.Y.; Kim, D.H.; Kwon, M.S.; Lee, S.U.; Kim, M.O.; Cho, S.; Lee, S.; Lee, H.J.; et al. Selective Novel Inverse Agonists for Human GPR43 Augment GLP-1 Secretion. Eur. J. Pharmacol. 2016, 771, 1–9. [Google Scholar] [CrossRef]
- Baudry, A.; Pietri, M.; Launay, J.M.; Kellermann, O.; Schneider, B. Multifaceted Regulations of the Serotonin Transporter: Impact on Antidepressant Response. Front. Neurosci. 2019, 13, 91:1–91:13. [Google Scholar] [CrossRef]
- Matheus, N.; Mendoza, C.; Iceta, R.; Mesonero, J.E.; Alcalde, A.I. Regulation of serotonin transporter activity by adenosine in intestinal epithelial cells. Biochem. Pharmacol. 2009, 78, 1198–1204. [Google Scholar] [CrossRef]
- Banskota, S.; Brim, H.; Kwon, Y.H.; Singh, G.; Sinha, S.R.; Wang, H.; Khan, W.I.; Ashktorab, H. Saffron Pre-Treatment Promotes Reduction in Tissue Inflammatory Profiles and Alters Microbiome Composition in Experimental Colitis Mice. Molecules 2021, 26, 3351. [Google Scholar] [CrossRef]
- Kwon, Y.H.; Wang, H.; Denou, E.; Ghia, J.E.; Rossi, L.; Fontes, M.E.; Bernier, S.P.; Shajib, M.S.; Banskota, S.; Collins, S.M.; et al. Modulation of Gut Microbiota Composition by Serotonin Signaling Influences Intestinal Immune Response and Susceptibility to Colitis. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 709–728. [Google Scholar] [CrossRef]
- Ahmed, H.; Leyrolle, Q.; Koistinen, V.; Kärkkäinen, O.; Layé, S.; Delzenne, N.; Hanhineva, K. Microbiota-Derived Metabolites as Drivers of Gut–Brain Communication. Gut Microbes 2022, 14, 2102878:1–2102878:33. [Google Scholar] [CrossRef]
- El-Salhy, M.; Valeur, J.; Hausken, T.; Gunnar Hatlebakk, J. Changes in Fecal Short-Chain Fatty Acids Following Fecal Microbiota Transplantation in Patients with Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2021, 33, e13983:1–e13983:11. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| SERT | AAATCCAAGCACCCAGAGAT | AGACTGTGTCCCTGTGGAGA |
| 5-HTR1A | AACAACAACACATCACCACCGGC | AGATGCTCCATGGCGGGTGT |
| 5-HTR2A | CAACTACGAACTCCCTAATG | AAACAGGAAGAAGACGATGC |
| 5-HTR2B | GAATCACAGAAAACAGCAAATGG | CCCATTTCGAATTCCATGTT |
| 5-HTR3A | GCCCTACTTTCGGGAGTTCAGCAG | TCTTGGTGGCTTGGGAGGTG |
| 5-HTR4 | CCTGTAATGGACAACTTGA | CCATGTTATTCCAGCCTTG |
| 5-HTR7 | AGAGAAGCCAGACGGAGAGAA | TACGGCAGAGTCGAGAAAGTG |
| GAPDH | CATGACCACAGTCCATGCCATCACT | TGAGGTCCACCACCCTGTTGCTGTA |
| HPRT1 | CTGACCTGCTGGATTACA | GCGACCTTGACCATCTTT |
| β-actin | AGCACGGCATCGTCACCAACT | ACATGGCTGGGGGTGTTGAAAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buey, B.; Forcén, A.; Grasa, L.; Layunta, E.; Mesonero, J.E.; Latorre, E. Gut Microbiota-Derived Short-Chain Fatty Acids: Novel Regulators of Intestinal Serotonin Transporter. Life 2023, 13, 1085. https://doi.org/10.3390/life13051085
Buey B, Forcén A, Grasa L, Layunta E, Mesonero JE, Latorre E. Gut Microbiota-Derived Short-Chain Fatty Acids: Novel Regulators of Intestinal Serotonin Transporter. Life. 2023; 13(5):1085. https://doi.org/10.3390/life13051085
Chicago/Turabian StyleBuey, Berta, Ana Forcén, Laura Grasa, Elena Layunta, Jose Emilio Mesonero, and Eva Latorre. 2023. "Gut Microbiota-Derived Short-Chain Fatty Acids: Novel Regulators of Intestinal Serotonin Transporter" Life 13, no. 5: 1085. https://doi.org/10.3390/life13051085
APA StyleBuey, B., Forcén, A., Grasa, L., Layunta, E., Mesonero, J. E., & Latorre, E. (2023). Gut Microbiota-Derived Short-Chain Fatty Acids: Novel Regulators of Intestinal Serotonin Transporter. Life, 13(5), 1085. https://doi.org/10.3390/life13051085