

Numerical Response of Owls to the Dampening of Small Mammal Population Cycles in Latvia


Abstract
1. Introduction
2. Materials and Methods
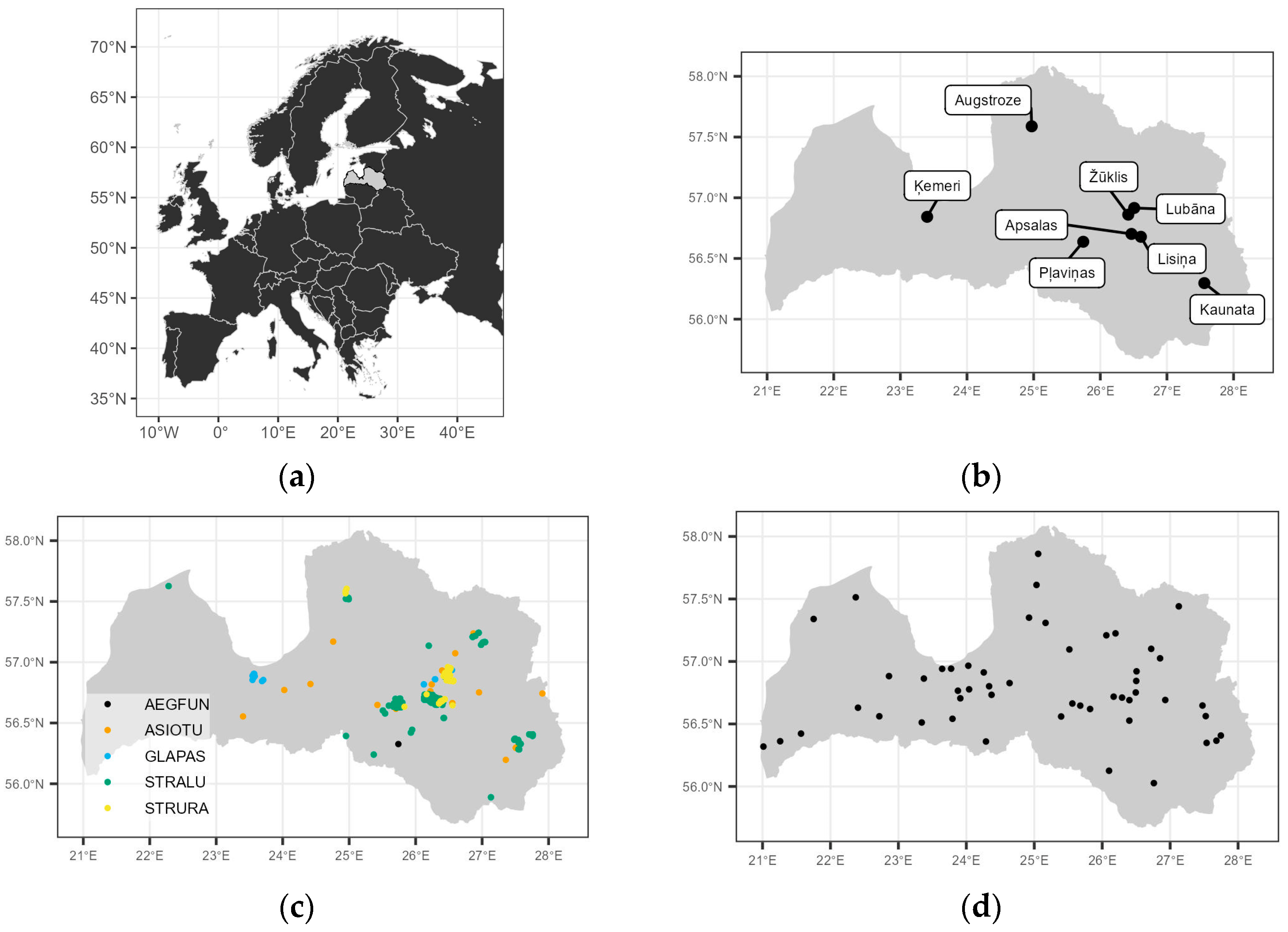
2.1. Location and Field Methods
2.1.1. Small Mammal Monitoring
- YP—young (clearcuts and stands <7 years old) stands on poor soils;
- YF—young (clearcuts and stands <7 years old) stands on fertile soils;
- MPU—medium-aged (between 8 years and 80% of rotation age) stands on poor soils without drainage;
- MFU—medium-aged (between 8 years and 80% of rotation age) stands on fertile soils without drainage;
- MPD—medium-aged (between 8 years and 80% of rotation age) stands on poor drained soils;
- MFD—medium-aged (between 8 years and 80% of rotation age) stands on fertile drained soils;
- OPU—older (≥80% of rotation age) stands on poor soils without drainage;
- OFU—older (≥80% of rotation age) stands on fertile soils without drainage;
- OPD—older (≥80% of rotation age) stands on poor drained soils;
- OFD—older (≥80% of rotation age) stands on fertile drained soils.
2.1.2. Owl Diet during the Breeding Season
2.1.3. Owl Population Change Monitoring
2.1.4. Owl Breeding Performance
2.2. Data Analysis
2.2.1. Small Mammal Monitoring
- Random intercept per transect and the comparable variable in the fixed part;
- Random intercept per transect and the comparable variable and year as a factor in the fixed part.
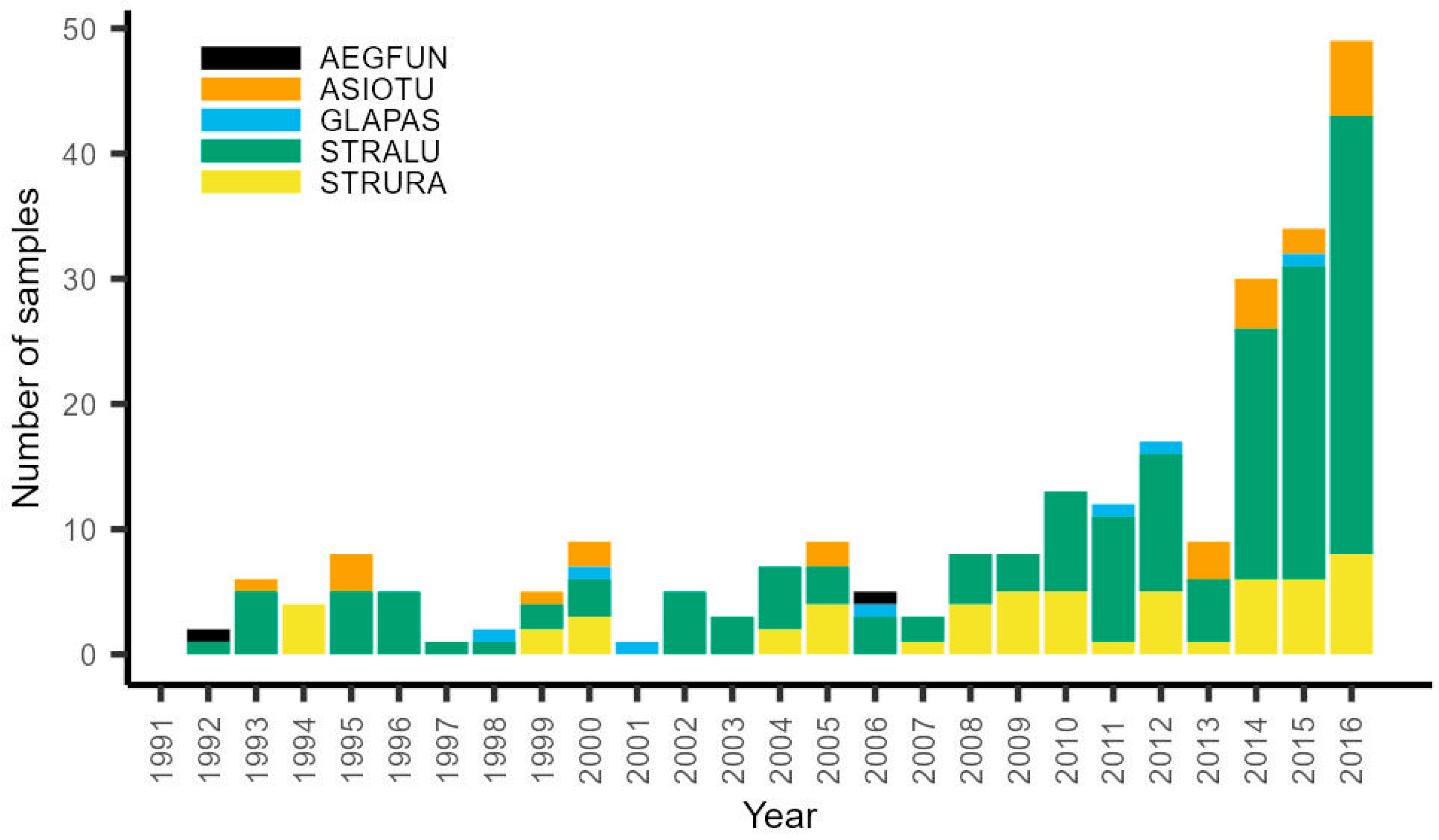
2.2.2. Owl Diet during the Breeding Season
2.2.3. Owl Population Change Monitoring
- The approximate time since when the small mammal populations did not recover to previous peaks;
- The approximate midpoint of small mammal monitoring;
- The approximate midpoint of STRURA monitoring;
- The beginning of GLAPAS monitoring.
2.2.4. Owl Breeding Performance
3. Results
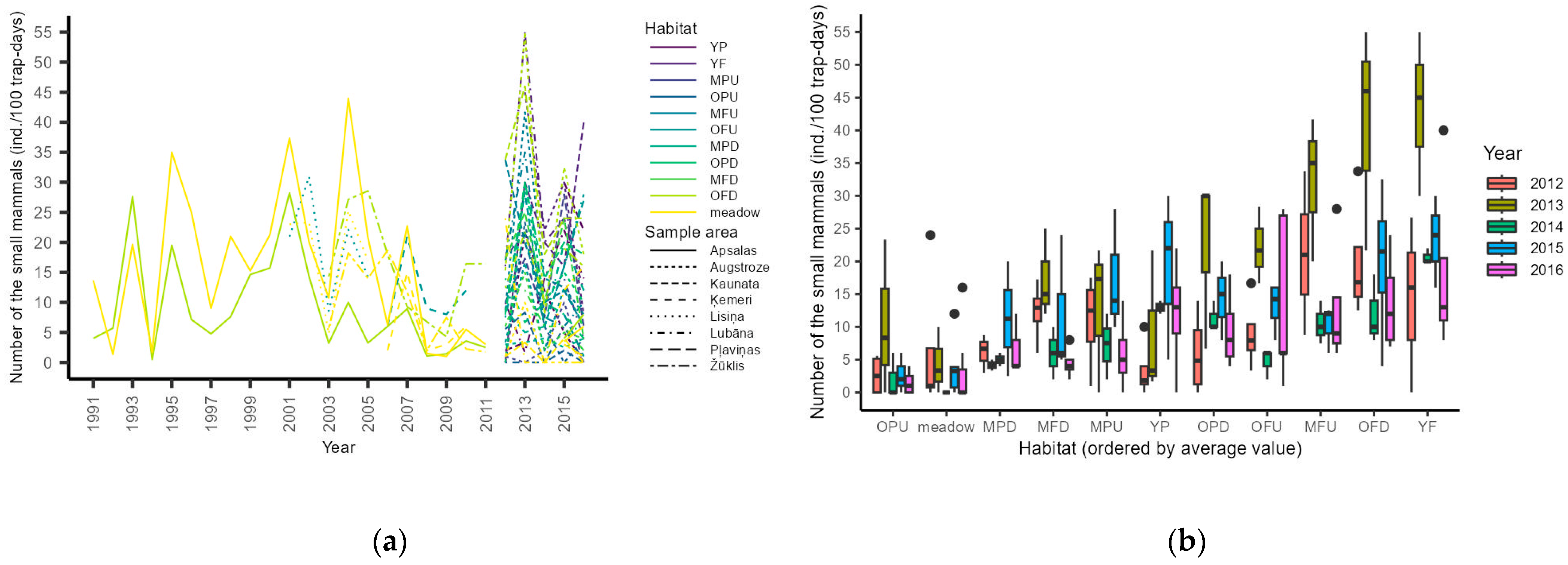
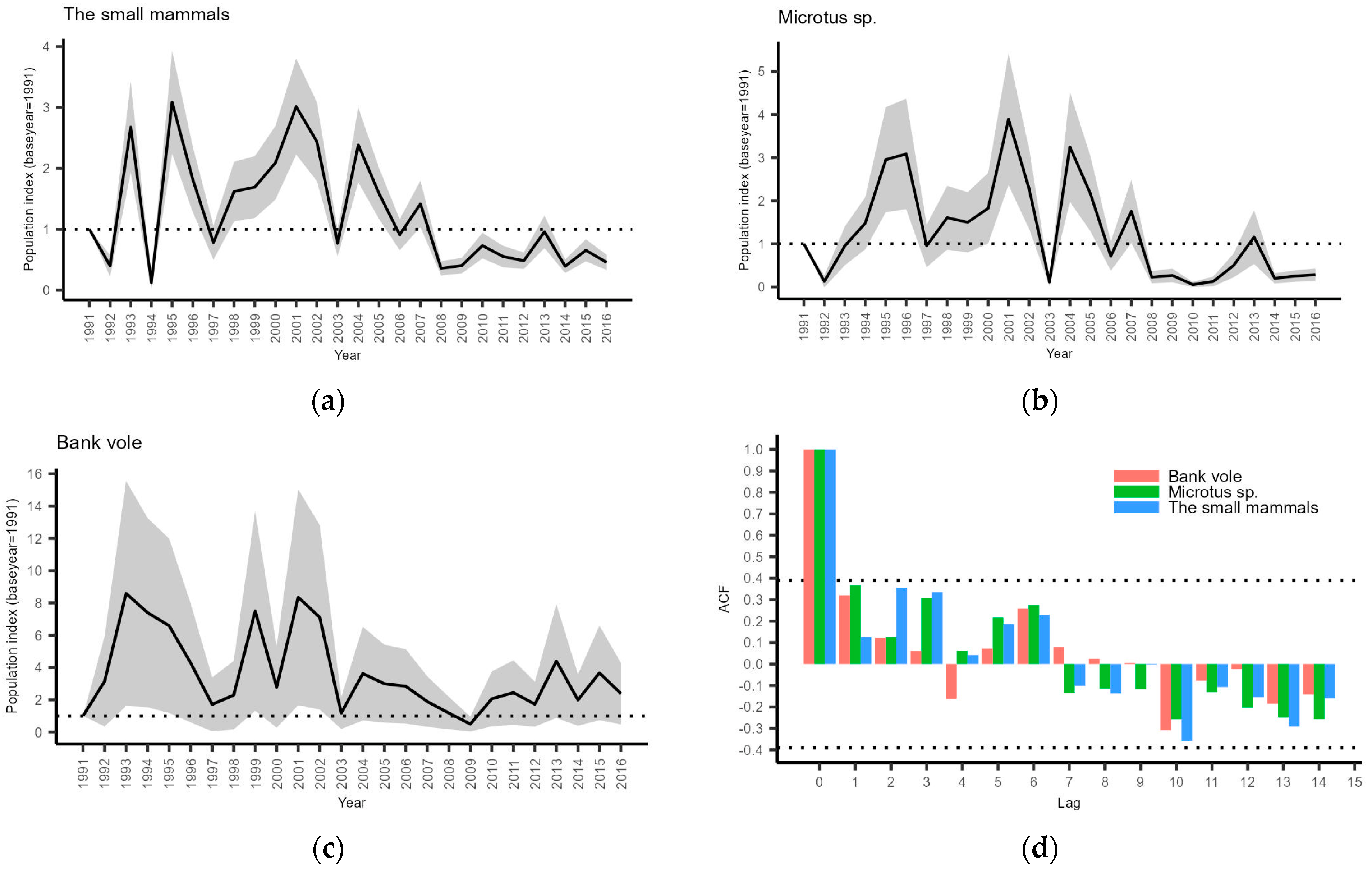
3.1. Small Mammal Monitoring
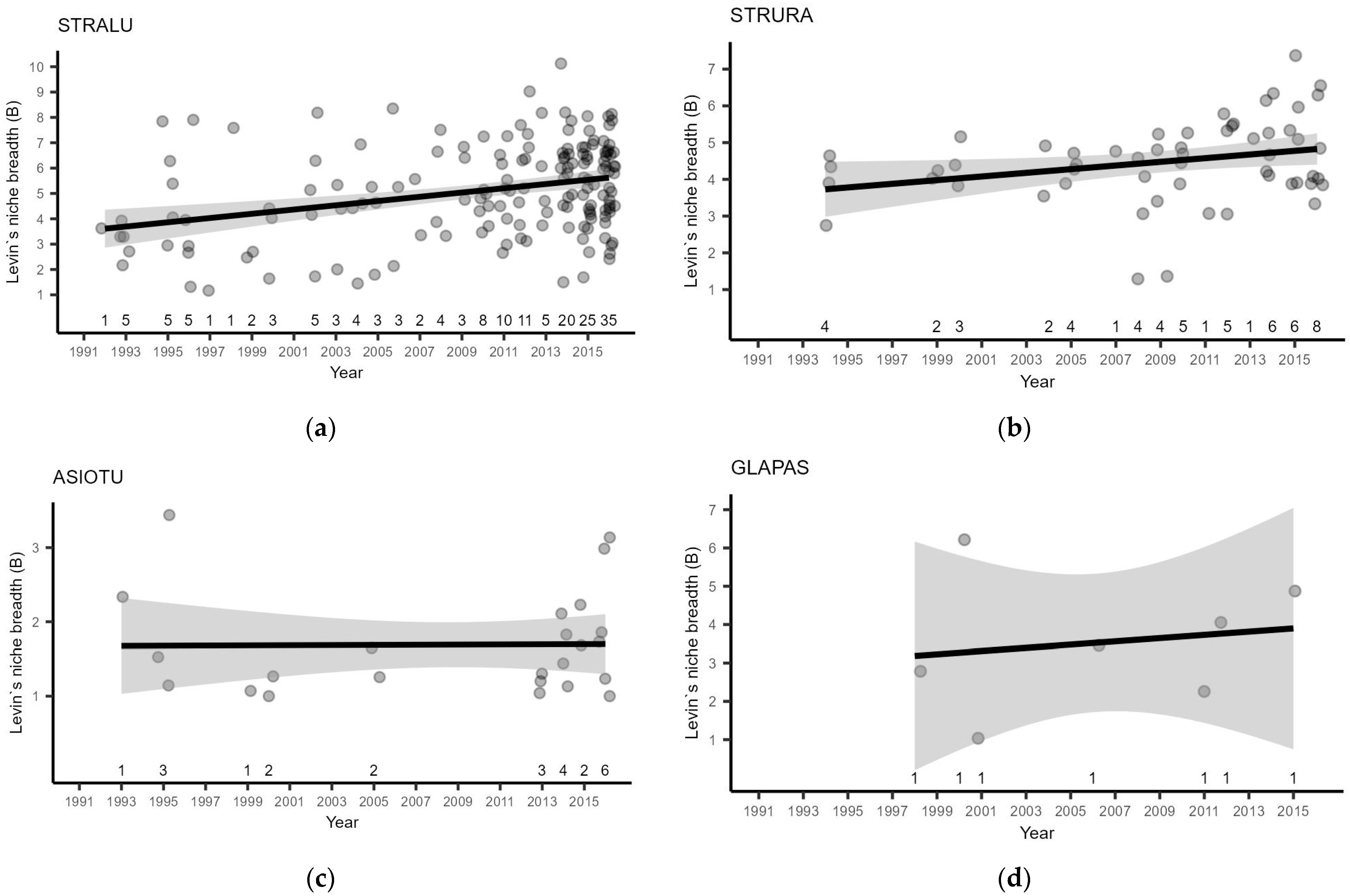
3.2. Owl Breeding Season Diet
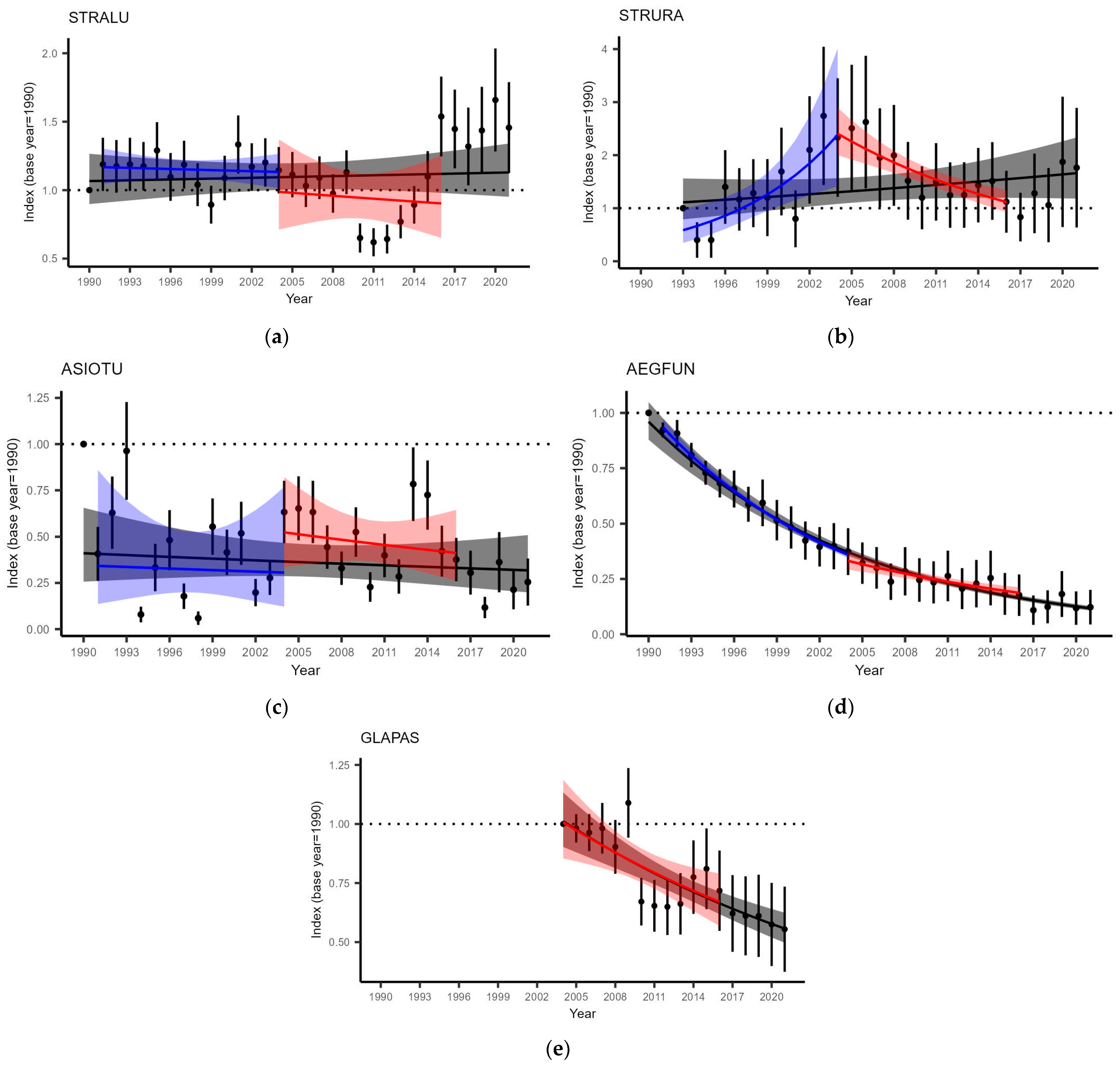
3.3. Owl Population Change
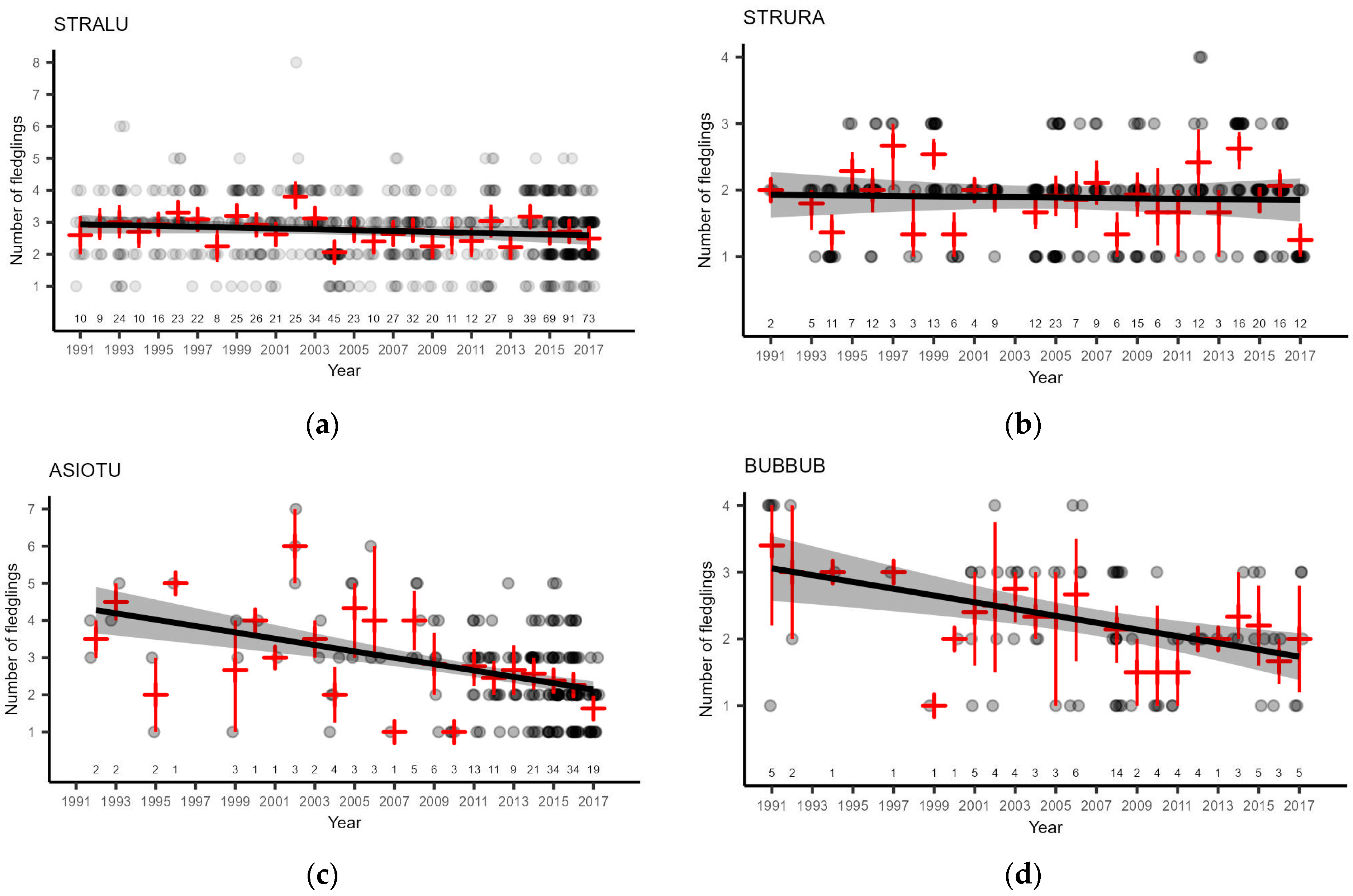
3.4. Owl Breeding Performance
4. Discussion
4.1. Small Mammal Monitoring
4.2. Numerical Response of Owls
4.2.1. Long-Eared Owl
4.2.2. Tengmalm’s Owl
4.2.3. Eurasian Pygmy Owl
4.2.4. Ural Owl
4.2.5. Tawny Owl
4.2.6. Eagle Owl
5. Conclusions
- Small mammal relative abundance indices have shown depleted population cycles since approx. 2004. This has impacted the breeding performance, food niche breadth, and population trends of owl species to various degrees depending on the particular species;
- The number of ASIOTU fledglings has declined since the depletion of small mammal populations. The population size of the species declined later and was significant for the period from 2004 to 2021. ASIOTU is the most specialized of the analyzed owl species in terms of the proportion of voles in the diet;
- The breeding performance of the three forest specialist species AEGFUN, GLAPAS, and STRURA in Latvia was similar to vole depression years in the boreal and boreonemoral regions;
- Populations of GLAPAS and AEGFUN declined in Latvia and showed no difference compared to periods with pronounced or depleted population dynamics of small mammals. In contrast, the population of STRURA has shown a significant decline since rodent depression. We suggest the depletion of the small mammal population dynamics to be an important negative contributing factor to more important effects of forestry, although the impact of forestry needs to be investigated further;
- Neither the breeding performance nor population size of STRALU changed between the compared periods with pronounced and depleted population dynamics of small mammals. This suggests a strong plasticity of the species, as food niche breadth was temporarily increased;
- We found evidence that suggests the dependency of BUBBUB on voles via a carry-over effect. The breeding performance of BUBBUB was significantly correlated with the abundance indices of small mammals in nature in the previous autumn.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Tree Species | Highest Quality | Medium Quality | Lowest Quality |
|---|---|---|---|
| Oaks | 101 | 121 | 121 |
| Pines and larches | 101 | 101 | 121 |
| Spruces, ashes, limes, elms, and maples | 81 | 81 | 81 |
| Birches | 71 | 71 | 51 |
| Black alders | 71 | 71 | 71 |
| Aspens | 41 | 41 | 41 |
| Sample Areas Contrasted | Ratio ± SE | z-Ratio | p-Value |
|---|---|---|---|
| Apsalas/Augstroze | 0.704 ± 0.459 | −0.538 | 0.9983 |
| Apsalas/Kaunata | 0.468 ± 0.307 | −1.157 | 0.9099 |
| Apsalas/Ķemeri | 1.816 ± 1.587 | 0.682 | 0.9936 |
| Apsalas/Lubāna | 0.538 ± 0.35 | −0.951 | 0.964 |
| Apsalas/Pļaviņas | 0.566 ± 0.371 | −0.868 | 0.9772 |
| Apsalas/Žūklis | 1.329 ± 1.176 | 0.322 | 0.9999 |
| Augstroze/Kaunata | 0.665 ± 0.236 | −1.148 | 0.9132 |
| Augstroze/Ķemeri | 2.578 ± 1.752 | 1.393 | 0.8057 |
| Augstroze/Lubāna | 0.764 ± 0.265 | −0.774 | 0.9874 |
| Augstroze/Pļaviņas | 0.804 ± 0.286 | −0.614 | 0.9964 |
| Augstroze/Žūklis | 1.887 ± 1.308 | 0.917 | 0.9701 |
| Kaunata/Ķemeri | 3.877 ± 2.65 | 1.983 | 0.4257 |
| Kaunata/Lubāna | 1.149 ± 0.408 | 0.393 | 0.9997 |
| Kaunata/Pļaviņas | 1.209 ± 0.439 | 0.522 | 0.9985 |
| Kaunata/Žūklis | 2.839 ± 1.978 | 1.498 | 0.7465 |
| Ķemeri/Lubāna | 0.296 ± 0.201 | −1.79 | 0.5547 |
| Ķemeri/Pļaviņas | 0.312 ± 0.213 | −1.705 | 0.6123 |
| Ķemeri/Žūklis | 0.732 ± 0.663 | −0.344 | 0.9999 |
| Lubāna/Pļaviņas | 1.052 ± 0.373 | 0.142 | 1 |
| Lubāna/Žūklis | 2.47 ± 1.711 | 1.305 | 0.8498 |
| Pļaviņas/Žūklis | 2.349 ± 1.633 | 1.228 | 0.8834 |
| Forest Age Groups Contrasted | Ratio ± SE | z-Ratio | p-Value |
|---|---|---|---|
| Young/Meadow | 7.078 ± 2.772 | 4.997 | <0.0001 |
| Young/Medium | 1.458 ± 0.442 | 1.244 | 0.599 |
| Young/Old | 1.627 ± 0.477 | 1.661 | 0.3446 |
| Meadow/Medium | 0.206 ± 0.073 | −4.451 | 0.0001 |
| Meadow/Old | 0.23 ± 0.079 | −4.257 | 0.0001 |
| Medium/Old | 1.116 ± 0.27 | 0.452 | 0.9692 |
| Soil Fertility Classes Contrasted | Ratio ± SE | z-Ratio | p-Value |
|---|---|---|---|
| Meadow/Fertile | 0.153 ± 0.047 | −6.086 | <0.0001 |
| Meadow/Poor | 0.309 ± 0.098 | −3.717 | 0.0006 |
| Fertile/Poor | 2.026 ± 0.397 | 3.604 | 0.0009 |
References
- McCaffery, R.; Jenkins, K.J.; Cendejas-Zarelli, S.; Happe, P.J.; Sager-Fradkin, K.A. Small Mammals and Ungulates Respond to and Interact with Revegetation Processes Following Dam Removal. Food Webs 2020, 25, e00159. [Google Scholar] [CrossRef]
- Moorhead, L.C.; Souza, L.; Habeck, C.W.; Lindroth, R.L.; Classen, A.T. Small Mammal Activity Alters Plant Community Composition and Microbial Activity in an Old-Field Ecosystem. Ecosphere 2017, 8, e01777. [Google Scholar] [CrossRef]
- Saurola, P.; Francis, C. Towards Integrated Population Monitoring Based on the Fieldwork of Volunteer Ringers: Productivity, Survival and Population Change of Tawny Owls Strix aluco and Ural Owls Strix uralensis in Finland. Bird Study 2018, 65, S63–S76. [Google Scholar] [CrossRef]
- Karell, P.; Ahola, K.; Karstinen, T.; Zolei, A.; Brommer, J.E. Population Dynamics in a Cyclic Environment: Consequences of Cyclic Food Abundance on Tawny Owl Reproduction and Survival. J. Anim. Ecol. 2009, 78, 1050–1062. [Google Scholar] [CrossRef]
- Solonen, T. Breeding of the Tawny Owl Strix aluco in Finland: Responses of a Southern Colonist to the Highly Variable Environment of the North. Ornis Fenn. 2005, 82, 97–106. [Google Scholar]
- Lehikoinen, A.; Ranta, E.; Pietiäinen, H.; Byholm, P.; Saurola, P.; Valkama, J.; Huitu, O.; Henttonen, H.; Korpimäki, E. The Impact of Climate and Cyclic Food Abundance on the Timing of Breeding and Brood Size in Four Boreal Owl Species. Oecologia 2011, 165, 349–355. [Google Scholar] [CrossRef]
- Blomqvist, S.; Holmgren, N.; Åkesson, S.; Hedenström, A.; Pettersson, J. Indirect Effects of Lemming Cycles on Sandpiper Dynamics: 50 Years of Counts from Southern Sweden. Oecologia 2002, 133, 146–158. [Google Scholar] [CrossRef]
- Summers, R.W. Breeding Production of Dark-Billed Brent Geese Branta bernicla bernicla in Relation to Lemming Cycles. Bird Study 1986, 33, 105–108. [Google Scholar] [CrossRef]
- Root-Bernstein, M.; Fierro, A.; Armesto, J.; Ebensperger, L.A. Avian Ecosystem Functions Are Influenced by Small Mammal Ecosystem Engineering. BMC Res. Notes 2013, 6, 549. [Google Scholar] [CrossRef]
- Sundell, J.; Huitu, O.; Henttonen, H.; Kaikusalo, A.; Korpimäki, E.; Pietiäinen, H.; Saurola, P.; Hanski, I. Large-Scale Spatial Dynamics of Vole Populations in Finland Revealed by the Breeding Success of Vole-Eating Avian Predators. J. Anim. Ecol. 2004, 73, 167–178. [Google Scholar] [CrossRef]
- Lindén, H. Latitudinal Gradients in Predator-Prey Interactions, Cyclicity and Synchronism in Voles and Small Game Populations in Finland. Oikos 1988, 52, 341–349. [Google Scholar] [CrossRef]
- Hansson, L.; Henttonen, H. Gradients in Density Variations of Small Rodents: The Importance of Latitude and Snow Cover. Oecologia 1985, 67, 394–402. [Google Scholar] [CrossRef]
- Lambin, X.; Bretagnolle, V.; Yoccoz, N.G. Vole Population Cycles in Northern and Southern Europe: Is There a Need for Different Explanations for Single Pattern? J. Anim. Ecol. 2006, 75, 340–349. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. Long-Term Changes in a Small Mammal Community in a Temperate Zone Meadow Subject to Seasonal Floods and Habitat Transformation. Integr. Zool. 2022, 17, 443–455. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. Small Mammal Diversity Changes in a Baltic Country, 1975–2021: A Review. Life 2022, 12, 1887. [Google Scholar] [CrossRef]
- Väli, Ü.; Tõnisalu, G. Community- and Species-Level Habitat Associations of Small Mammals in a Hemiboreal Forest-Farmland Landscape. Ann. Zool. Fenn. 2020, 58, 1–11. [Google Scholar] [CrossRef]
- Hanski, I.; Hansson, L.; Henttonen, H. Specialist Predator, Generalist Predator, and the Microtine Rodent Cycle. J. Anim. Ecol. 1991, 60, 353–367. [Google Scholar] [CrossRef]
- Henttonen, H.; Oksanen, T.; Jortikka, A.; Haukisalmi, V. How Much Do Weasels Shape Microtine Cycles in the Northern Fennoscandian Taiga? Oikos 1987, 50, 353. [Google Scholar] [CrossRef]
- Hanski, I.; Henttonen, H. Predation on Competing Rodent Species: A Simple Explanation of Complex Patterns. J. Anim. Ecol. 1996, 65, 220. [Google Scholar] [CrossRef]
- Hörnfeldt, B. Long-Term Decline in Numbers of Cyclic Voles in Boreal Sweden: Analysis and Presentation of Hypotheses. Oikos 2004, 107, 376–392. [Google Scholar] [CrossRef]
- Hörnfeldt, B.; Hipkiss, T.; Eklund, U. Fading out of Vole and Predator Cycles? Proc. R. Soc. B Biol. Sci. 2005, 272, 2045–2049. [Google Scholar] [CrossRef] [PubMed]
- Steen, H.; Ims, R.A.; Sonerud, G.A.; Ecology, S.; Dec, N. Spatial and Temporal Patterns of Small-Rodent Population Dynamics at a Regional Scale. Ecology 1996, 77, 2365–2372. [Google Scholar] [CrossRef]
- Kausrud, K.L.; Mysterud, A.; Steen, H.; Vik, J.O.; Østbye, E.; Cazelles, B.; Framstad, E.; Eikeset, A.M.; Mysterud, I.; Solhøy, T.; et al. Linking Climate Change to Lemming Cycles. Nature 2008, 456, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Ims, R.A.; Henden, J.A.; Killengreen, S.T. Collapsing Population Cycles. Trends Ecol. Evol. 2008, 23, 79–86. [Google Scholar] [CrossRef]
- Cornulier, T.; Yoccoz, N.G.; Bretagnolle, V.; Brommer, J.E.; Butet, A.; Ecke, F.; Elston, D.A.; Framstad, E.; Henttonen, H.; Hörnfeldt, B.; et al. Europe-Wide Dampening of Population Cycles in Keystone Herbivores. Science 2013, 340, 63–66. [Google Scholar] [CrossRef]
- Brommer, J.E.; Pietiäinen, H.; Ahola, K.; Karell, P.; Karstinenz, T.; Kolunen, H. The Return of the Vole Cycle in Southern Finland Refutes the Generality of the Loss of Cycles through “Climatic Forcing”. Glob. Change Biol. 2010, 16, 577–586. [Google Scholar] [CrossRef]
- Birrer, S. Synthesis of 312 Studies on the Diet of the Long-Eared Owl Asio otus. Ardea 2009, 97, 615–624. [Google Scholar] [CrossRef]
- Balčiauskienė, L.; Jovaišas, A.; Naruševičius, V.; Petraška, A.; Skuja, S. Diet of Tawny Owl (Strix aluco) and Long-Eared Owl (Asio otus) in Lithuania as Found from Pellets. Acta Zool. Lith. 2006, 16, 37–45. [Google Scholar] [CrossRef]
- Village, A. The Diet and Breeding of Long-Eared Owls in Relation to Vole Numbers. Bird Study 1981, 28, 215–225. [Google Scholar] [CrossRef]
- Tome, D. Functional Response of the Long-Eared Owl (Asio otus) to Changing Prey Numbers: A 20-Year Study. Ornis Fenn. 2003, 80, 63–70. [Google Scholar]
- Tome, D. Changes in the Diet of Long-Eared Owl Asio otus: Seasonal Patterns of Dependence on Vole Abundance. Ardeola 2009, 56, 49–56. [Google Scholar]
- Korpimäki, E.; Hakkarainen, H. The Boreal Owl: Ecology, Behaviour, and Conservation of a Forest-Dwelling Predator; Cambridge University Press: Cambridge, UK, 2012; ISBN 9781107425323. [Google Scholar]
- Masoero, G.; Laaksonen, T.; Morosinotto, C.; Korpimäki, E. Age and Sex Differences in Numerical Responses, Dietary Shifts, and Total Responses of a Generalist Predator to Population Dynamics of Main Prey. Oecologia 2020, 192, 699–711. [Google Scholar] [CrossRef]
- Mikkola, H. Owls of Europe; A.D. & T. Poyser: Calton, UK, 1983. [Google Scholar]
- Vrezec, A.; Saurola, P.; Avotins, A.; Kocijančič, S.; Sulkava, S. A Comparative Study of Ural Owl Strix uralensis Breeding Season Diet within Its European Breeding Range, Derived from Nest Box Monitoring Schemes. Bird Study 2018, 65, S85–S95. [Google Scholar] [CrossRef]
- Grašytė, G.; Rumbutis, S.; Dagys, M.; Treinys, R. Breeding Performance, Apparent Survival, Nesting Location and Diet in a Local Population of the Tawny Owl Strix aluco in Central Lithuania Over the Long-Term. Acta Ornithol. 2016, 51, 163–174. [Google Scholar] [CrossRef]
- Penteriani, V.; del Mar Delgado, M. The Eagle Owl; T & A.D. Poyser: London, UK, 2019; ISBN 978-1-4729-0066-1. [Google Scholar]
- Korpimäki, E.; Hakkarainen, H. Fluctuating Food Supply Affects the Clutch Size of Tengmalm’s Owl Independent of Laying Date. Oecologia 1991, 85, 543–552. [Google Scholar] [CrossRef]
- Saurola, P.; Francis, C.M. Estimating Population Parameters of Owls from Nationally Coordinated Ringing Data in Finland. Anim. Biodivers. Conserv. 2004, 27, 403–415. [Google Scholar]
- Solheim, R. Breeding Biology of the Pygmy Owl Glaucidium passerinum in Two Biogeographical Zones in Southeastern Norway. Ann. Zool. Fenn. 1984, 21, 295–300. [Google Scholar]
- Brommer, J.E.; Pietiäinen, H.; Kolunen, H. Reproduction and Survival in a Variable Environment: Ural Owls (Strix uralensis) and the Three-Year Vole Cycle. Auk 2002, 119, 544–550. [Google Scholar] [CrossRef]
- Tome, D. Post-Fledging Survival and Dynamics of Dispersal in Long-Eared Owls Asio otus. Bird Study 2011, 58, 193–199. [Google Scholar] [CrossRef]
- Hakkarainen, H.; Korpimäki, E.; Koivunen, V.; Ydenberg, R. Survival of Male Tengmalm’s Owls under Temporally Varying Food Conditions. Oecologia 2002, 131, 83–88. [Google Scholar] [CrossRef]
- Masoero, G.; Laaksonen, T.; Morosinotto, C.; Korpimäki, E. Climate Change and Perishable Food Hoards of an Avian Predator: Is the Freezer Still Working? Glob. Change Biol. 2020, 26, 5414–5430. [Google Scholar] [CrossRef] [PubMed]
- Pavón-Jordán, D.; Karell, P.; Ahola, K.; Kolunen, H.; Pietiäinen, H.; Karstinen, T.; Brommer, J.E. Environmental Correlates of Annual Survival Differ between Two Ecologically Similar and Congeneric Owls. Ibis 2013, 155, 823–834. [Google Scholar] [CrossRef]
- Kontiainen, P.; Pietiäinen, H.; Huttunen, K.; Karell, P.; Kolunen, H.; Brommer, J.E. Aggressive Ural Owl Mothers Recruit More Offspring. Behav. Ecol. 2009, 20, 789–796. [Google Scholar] [CrossRef]
- Santangeli, A.; Hakkarainen, H.; Laaksonen, T.; Korpimäki, E. Home Range Size Is Determined by Habitat Composition but Feeding Rate by Food Availability in Male Tengmalm’s Owls. Anim. Behav. 2012, 83, 1115–1123. [Google Scholar] [CrossRef]
- Brommer, J.E.; Karell, P.; Pietiäinen, H. Supplementary Fed Ural Owls Increase Their Reproductive Output with a One Year Time Lag. Oecologia 2004, 139, 354–358. [Google Scholar] [CrossRef]
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation Zones and Their Sections in Northwestern Europe. Ann. Bot. Fenn. 1968, 5, 169–211. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated World Map of the Köppen-Geiger Climate Classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; et al. European Breeding Bird Atlas 2: Distribution, Abundance and Change; European Bird Census Council & Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- Pupila, A.; Bergmanis, U. Species Diversity, Abundance and Dynamics of Small Mammals in the Eastern Latvia. Acta Univ. Latv. 2006, 710, 93–101. [Google Scholar]
- Demongin, L. Identification Guide to Birds in the Hand; Laurent Demongin: Beauregard-Vendon, France, 2016; ISBN 9782955501900. [Google Scholar]
- Tauriņš, E. Latvijas Zīdītājdzīvnieki; Zvaigzne: Rīga, Latvia, 1982. [Google Scholar]
- Avotiņš, A.; Ķemlers, A. Monitoring of Owls in Latvia. Ring 1993, 15, 104–114. [Google Scholar]
- Avotiņš, A.; Graubics, G.; Ķemlers, A.; Krēsliņš, V.; Ķuze, J.; Ļoļāns, U. Number and Breeding Densities of Owls in Latvia: Studies in Sample Plots. Vogelwelt 1999, 120, 333–337. [Google Scholar]
- Avotiņš, A. Owl Census in Sample Plots near Metriena and Laudona, Eastern Latvia. Bird Census News 1999, 12, 52–62. [Google Scholar]
- Avotiņš, A. Changes of Number and Structure in Population of Tawny Owl (Strix aluco) in Sample Plots at Eastern Latvia (1990–1994). Popul. Greifvogel Eulenarten 1996, 3, 377–386. [Google Scholar]
- Avotiņš, A. Tawny Owl’s Territory Occupancy in Eastern Latvia. Bird Census News 2004, 13, 167–173. [Google Scholar]
- Avotiņš, A.; Reihmanis, J. Plēsīgo Putnu Monitorings; Latvijas Ornitoloģijas Biedrība: Riga, Latvia, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: Berling/Heidelberg, Germany, 2002; ISBN 978-0-387-22456-5. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 26 July 2022).
- Bogaart, P.; van der Loo, M.; Pannekoek, J. Rtrim: Trends and Indices for Monitoring Data. Available online: https://CRAN.R-project.org/package=rtrim (accessed on 26 July 2022).
- Pannekoek, J.; Bogaart, P.; van der Loo, M. Models and Statistical Methods in Rtrim; Statistics Netherlands: Heerlen, The Netherlands, 2018; pp. 1–34.
- Pucek, Z. Keys to Vertebrates of Poland. Mammals; PWN—Polish Scientific Publishers: Warsaw, Poland, 1981. [Google Scholar]
- Smith, E.P. Niche Breadth, Resource Availability, and Inference. Ecology 1982, 63, 1675–1681. [Google Scholar] [CrossRef]
- Oberpriller, J.; de Souza Leite, M.; Pichler, M. Fixed or Random? On the Reliability of Mixed-Effects Models for a Small Number of Levels in Grouping Variables. Ecol. Evol. 2022, 12, e9062. [Google Scholar] [CrossRef]
- Gomes, D.G.E. Should I Use Fixed Effects or Random Effects When I Have Fewer than Five Levels of a Grouping Factor in a Mixed-Effects Model? PeerJ 2022, 10, e12794. [Google Scholar] [CrossRef]
- Pannekoek, J.; Strien, A. Van TRIM 3 Manual (TRends & Indices for Monitoring Data); Statistics Netherlands: Heerlen, The Netherlands, 2005; pp. 1–49. [Google Scholar]
- Van Strien, A.J.; Pannekoek, J.; Gibbons, D.W. Indexing European Bird Population Trends Using Results of National Monitoring Schemes: A Trial of a New Method. Bird Study 2001, 48, 200–213. [Google Scholar] [CrossRef]
- Harrell, F.E., Jr. Hmisc: Harrell Miscellaneous. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 26 July 2022).
- Scott, D.M.; Joyce, C.B.; Burnside, N.G. The Influence of Habitat and Landscape on Small Mammals in Estonian Coastal Wetlands. Est. J. Ecol. 2008, 57, 279–295. [Google Scholar] [CrossRef]
- Balčiauskas, L. Small Mammal Communities in the Fragmented Landscape in Lithuania. Acta Zool. Litu. 2006, 16, 130–136. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Čepukienė, A.; Balčiauskienė, L. Small Mammal Community Response to Early Meadow–Forest Succession. For. Ecosyst. 2017, 4, 11. [Google Scholar] [CrossRef]
- Mažeikytė, R. Small Mammals in the Mosaic Landscape of Eastern Lithuania: Species Composition, Distribution and Abundance. Acta Zool. Litu. 2002, 12, 381–391. [Google Scholar] [CrossRef]
- Savola, S.; Henttonen, H.; Lindén, H. Vole Population Dynamics during the Succession of a Commercial Forest in Northern Finland. Ann. Zool. Fenn. 2013, 50, 79–88. [Google Scholar] [CrossRef]
- Wegge, P.; Rolstad, J. Cyclic Small Rodents in Boreal Forests and the Effects of Even-Aged Forest Management: Patterns and Predictions from a Long-Term Study in Southeastern Norway. For. Ecol. Manag. 2018, 422, 79–86. [Google Scholar] [CrossRef]
- Ecke, F.; Löfgren, O.; Sörlin, D. Population Dynamics of Small Mammals in Relation to Forest Age and Structural Habitat Factors in Northern Sweden. J. Appl. Ecol. 2002, 39, 781–792. [Google Scholar] [CrossRef]
- Suchomel, J.; Šipoš, J.; Košulič, O. Management Intensity and Forest Successional Stages as Significant Determinants of Small Mammal Communities in a Lowland Floodplain Forest. Forests 2020, 11, 1320. [Google Scholar] [CrossRef]
- Panzacchi, M.; Linnell, J.D.C.; Melis, C.; Odden, M.; Odden, J.; Gorini, L.; Andersen, R. Effect of Land-Use on Small Mammal Abundance and Diversity in a Forest-Farmland Mosaic Landscape in South-Eastern Norway. For. Ecol. Manag. 2010, 259, 1536–1545. [Google Scholar] [CrossRef]
- Carey, A.B.; Harrington, C.A. Small Mammals in Young Forests: Implications for Management for Sustainability. For. Ecol. Manag. 2001, 154, 289–309. [Google Scholar] [CrossRef]
- Bowman, J.C.; Sleep, D.; Forbes, G.J.; Edwards, M. The Association of Small Mammals with Coarse Woody Debris at Log and Stand Scales. For. Ecol. Manag. 2000, 129, 119–124. [Google Scholar] [CrossRef]
- Henttonen, H.; Gilg, O.; Ims, R.A.; Korpimäki, E.; Yoccoz, N.G. Ilkka Hanski and Small Mammals: From Shrew Metapopulations to Vole and Lemming Cycles. Ann. Zool. Fenn. 2017, 54, 153–162. [Google Scholar] [CrossRef]
- Hanski, I.; Korpimäki, E. Microtine Rodent Dynamics in Northern Europe: Parameterized Models for the Predator-Prey Interaction. Ecology 1995, 76, 840–850. [Google Scholar] [CrossRef]
- Tome, D. Diet Composition of the Long-Eared Owl in Central Slovenia—Seasonal-Variation in Prey Use. J. Raptor Res. 1994, 28, 253–258. [Google Scholar]
- Sergio, F.; Marchesi, L.; Pedrini, P. Density, Diet and Productivity of Long-Eared Owls Asio otus in the Italian Alps: The Importance of Microtus Voles. Bird Study 2008, 55, 321–328. [Google Scholar] [CrossRef]
- Wijnandts, H. Ecological Energetics of the Long-Eared Owl (Asio otus). Ardea 1984, 72, 1–92. [Google Scholar] [CrossRef]
- Glue, D.E.; Hammond, G.J. Feeding Ecology of the Long-Eared Owl in Britain and Ireland. Br. Birds 1974, 67, 361–369. [Google Scholar]
- Romanowski, J.; Zmihorski, M. Effect of Season, Weather and Habitat on Diet Variation of a Feeding-Specialist: A Case Study of the Long-Eared Owl, Asio otus in Central Poland. Folia Zool. 2008, 57, 411–419. [Google Scholar]
- Heisler, L.M.; Somers, C.M.; Poulin, R.G. Owl Pellets: A More Effective Alternative to Conventional Trapping for Broad-Scale Studies of Small Mammal Communities. Methods Ecol. Evol. 2016, 7, 96–103. [Google Scholar] [CrossRef]
- Glue, D. Breeding Biology of Long-Eared Owls. Br. Birds 1977, 70, 318–331. [Google Scholar]
- Glue, D.; Nilsson, I.N. Long-Eared Owl Asio Otus. In The EBCC Atlas of European Breeding Birds: Their Distribution and Abundance; Hagemeijer, E.J.M., Blair, M.J., Eds.; T & A.D. Poyser: London, UK, 1997. [Google Scholar]
- Meller, K.; Björklund, H.; Saurola, P.; Valkama, J. Petolintuvuosi 2016, Pesimistulokset Ja Kannankehitykset. Linnut-vuosikirja 2017, 2016, 16–31. [Google Scholar]
- BirdLife International. Asio Otus. Available online: https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T22689507A201150685.en (accessed on 9 December 2022).
- Korpimäki, E. Diet Composition, Prey Choice, and Breeding Success of Long-Eared Owls—Effects of Multiannual Fluctuations in Food Abundance. Can. J. Zool. Rev. Can. Zool. 1992, 70, 2373–2381. [Google Scholar] [CrossRef]
- Korpimäki, E. Population Dynamics of Fennoscandian Owls in Relation to Wintering Conditions and Between-Year Fluctuations of Food. In The Ecology and Conservation of European Owls; Joint Nature Conservation Committee: Peterborough, UK, 1992; pp. 1–10. [Google Scholar]
- Avotiņš Jun, A. Apodziņa Glaucidium Passerinum, Bikšainā Apoga Aegolius Funereus, Meža Pūces Strix Aluco, Urālpūces Strix Uralensis, Ausainās Pūces Asio Otus Un Ūpja Bubo Bubo Aizsardzības Plāns Plāns; LOB: Riga, Latvia, 2019. [Google Scholar]
- Martínez, J.A.; Zuberogoitia, I. Habitat Preferences for Long-Eared Owls Asio otus and Little Owls Athene noctua in Semi-Arid Environments at Three Spatial Scales. Bird Study 2004, 51, 163–169. [Google Scholar] [CrossRef]
- Aschwanden, J.; Birrer, S.; Jenni, L. Are Ecological Compensation Areas Attractive Hunting Sites for Common Kestrels (Falco tinnunculus) and Long-Eared Owls (Asio otus)? J. Ornithol. 2005, 146, 279–286. [Google Scholar] [CrossRef]
- Henrioux, F. Home Range and Habitat Use by the Long-Eared Owl in Northwestern Switzerland. J. Raptor Res. 2000, 34, 93–101. [Google Scholar]
- Galeotti, P.; Tavecchia, G.; Bonetti, A. Home-Range and Habitat Use of Long-Eared Owls in Open Farmalend (Po Plain, Northern Italy), in Relation to Prey Availability. J. Wildl. Res. 1997, 2, 137–145. [Google Scholar]
- Korpimäki, E. On the Ecology and Biology of Tengmalm’s Owl (Aegolius funereus) in Southern Ostrobothnia and Suomenselkä, Western Finland. Acta Univ. Oul. Ser. A Sci. Rer. Nat. 1981, 118, 1–84. [Google Scholar]
- Solheim, R. Breeding Frequency of Tengmalm’s Owl Aegolius funereus in Three Localities in 1974–1978. In Proceedings of the Third NOK, Ribe, Denmark, 3–8 August 1981; pp. 79–84. [Google Scholar]
- Hakkarainen, H.; Koivunen, V.; Korpimäki, E. Reproductive Success and Parental Effort of Tengmalm’s Owls: Effects of Spatial and Temporal Variation in Habitat Quality. Écoscience 1997, 4, 35–42. [Google Scholar] [CrossRef]
- Korpimäki, E. Clutch Size, Breeding Success and Brood Size Experiments in Tengmalm’ s Owl Aegolius funereus: A Test of Hypotheses. Ornis Scand. 1987, 18, 277–284. [Google Scholar] [CrossRef]
- Vrezec, A. Breeding Density and Altitudinal Distribution of the Ural, Tawny, and Boreal Owls in North Dinaric Alps (Central Slovenia). J. Raptor Res. 2003, 37, 55–62. [Google Scholar]
- Vrezec, A.; Tome, D. Habitat Selection and Patterns of Distribution in a Hierarchic Forest Owl Guild. Ornis Fenn. 2004, 81, 109–118. [Google Scholar]
- Kajtoch, Ł.; Matysek, M.; Figarski, T. Spatio-Temporal Patterns of Owl Territories in Fragmented Forests Are Affected by a Top Predator (Ural Owl). Ann. Zool. Fenn. 2016, 53, 165–174. [Google Scholar] [CrossRef]
- Eionet. Bird Population Status and Trends at the EU and Member State Levels 2013–2018. Available online: https://nature-art12.eionet.europa.eu/article12/ (accessed on 5 December 2022).
- Hakkarainen, H.; Korpimäki, E.; Laaksonen, T.; Nikula, A.; Suorsa, P. Survival of Male Tengmalm’s Owls Increases with Cover of Old Forest in Their Territory. Oecologia 2008, 155, 479–486. [Google Scholar] [CrossRef]
- Laaksonen, T.; Hakkarainen, H.; Korpimäki, E. Lifetime Reproduction of a Forest-Dwelling Owl Increases with Age and Area of Forests. Proc. R. Soc. B Biol. Sci. 2004, 271, 461–464. [Google Scholar] [CrossRef]
- Ravussin, P.-A.; Trolliet, D.; Willenegger, L.; Béguin, D.; Matalon, G. Choix Du Site de Nidification Chez La Chouette de Tengmalm Aegolius Funereus: Influence Des Nichoirs. Nos Oiseaux 2001, 5, 41–51. [Google Scholar]
- Sorbi, S. La Chouette de Tengmalm (Aegolius funereus) En Belgique. Synthèse et Mise à Jour Du Statut. Aves 1995, 32, 101–132. [Google Scholar]
- Shurulinkov, P.; Stoyanov, G. Some New Findings of Pigmy Owls Glaucidium passerinum and Tengmalm’s Owl Aegolius funerus in Western and Southern Bulgaria. Acrocephalus 2006, 27, 65–68. [Google Scholar]
- Brambilla, M.; Bassi, E.; Bergero, V.; Casale, F.; Chemollo, M.; Falco, R.; Longoni, V.; Saporetti, F.; Viganò, E. Modelling Distribution and Potential Overlap between Boreal Owl Aegolius funereus and Black Woodpecker Dryocopus martius Implications for Management and Monitoring Plans. Bird Conserv. Int. 2013, 23, 502–511. [Google Scholar] [CrossRef]
- Avotins, A. Modelling Owl and Woodpecker Habitat Suitability to Evaluate Forest Conservation in Latvia. In Proceedings of the 11th International Conference on Biodiversity Research, Daugavpils, Latvia, 20–22 October 2022; p. 17. [Google Scholar]
- Mikusek, R.; Kloubec, B.; Obuch, J. Diet of the Pygmy Owl (Glaucidium passerinum) in Eastern Central Europe. Buteo 2001, 12, 47–60. [Google Scholar]
- Morosinotto, C.; Villers, A.; Thomson, R.L.; Varjonen, R.; Korpimäki, E. Competitors and Predators Alter Settlement Patterns and Reproductive Success of an Intraguild Prey. Ecol. Monogr. 2017, 87, 4–20. [Google Scholar] [CrossRef]
- Lehikoinen, A.; Hokkanen, T.; Lokki, H. Young and Female-Biased Irruptions in Pygmy Owls Glaucidium passerinum in Southern Finland. J. Avian Biol. 2011, 42, 564–569. [Google Scholar] [CrossRef]
- Barbaro, L.; Blache, S.; Trochard, G.; Arlaud, C.; de Lacoste, N.; Kayser, Y. Hierarchical Habitat Selection by Eurasian Pygmy Owls Glaucidium passerinum in Old-Growth Forests of the Southern French Prealps. J. Ornithol. 2016, 157, 333–342. [Google Scholar] [CrossRef]
- Baroni, D.; Korpimäki, E.; Selonen, V.; Laaksonen, T. Tree Cavity Abundance and beyond: Nesting and Food Storing Sites of the Pygmy Owl in Managed Boreal Forests. For. Ecol. Manag. 2020, 460, 117818. [Google Scholar] [CrossRef]
- Baroni, D.; Masoero, G.; Korpimäki, E.; Morosinotto, C.; Laaksonen, T. Habitat Choice of a Secondary Cavity User Indicates Higher Avoidance of Disturbed Habitat during Breeding than during Food-Hoarding. For. Ecol. Manag. 2021, 483, 118925. [Google Scholar] [CrossRef]
- Pakkala, T.; Lindén, A.; Tiainen, J.; Tomppo, E.; Kouki, J. Indicators of Forest Biodiversity: Which Bird Species Predict High Breeding Bird Assemblage Diversity in Boreal Forests at Multiple Spatial Scales? Ann. Zool. Fenn. 2014, 51, 457–476. [Google Scholar] [CrossRef]
- Shurulinkov, P.; Ralev, A.; Daskalova, G.; Chakarov, N. Distribution, Numbers and Habitat of Pigmy Owl Glaucidium passerinum in Rhodopes Mts (S Bulgaria). Acrocephalus 2007, 28, 161–165. [Google Scholar]
- Sonerud, G.A. Risk of Nest Predation in Three Species of Hole Nesting Owls: Influence on Choice of Nesting Habitat and Incubation Behaviour. Ornis Scand. 1985, 16, 261. [Google Scholar] [CrossRef]
- Strom, H.; Sonerud, G.A. Home Range and Habitat Selection in the Pygmy Owl Glaucidium passerinum. Ornis Fenn. 2001, 78, 145–158. [Google Scholar]
- Avotins, A.; Kerus, V.; Aunins, A. National Scale Habitat Suitability Analysis to Evaluate and Improve Conservation Areas for a Mature Forest Specialist Species. Glob. Ecol. Conserv. 2022, 38, e02218. [Google Scholar] [CrossRef]
- Terraube, J.; Villers, A.; Poudré, L.; Varjonen, R.; Korpimäki, E. Increased Autumn Rainfall Disrupts Predator–Prey Interactions in Fragmented Boreal Forests. Glob. Change Biol. 2017, 23, 1361–1373. [Google Scholar] [CrossRef]
- Carey, A.B.; Johnson, M.L. Small Mammals in Managed, Naturally Young, and Old-Growth Forests. Ecol. Appl. 1995, 5, 336–352. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Apostolopoulou, E.; Mazaris, A.D.; Kallimanis, A.S.; Drakou, E.G.; Pantis, J.D. Human Activities in Natura 2000 Sites: A Highly Diversified Conservation Network. Environ. Manag. 2013, 51, 1025–1033. [Google Scholar] [CrossRef]
- Jäderholm, K. Diets of the Tengalm’s Owl Aegolius funereus and the Ural Owl Strix uralensis in Central Finland. Ornis Fenn. 1987, 64, 149–153. [Google Scholar]
- Korpimäki, E.; Sulkava, S. Diet and Breeding Performance of Ural Owl Strix uralensis under Fluctuating Food Conditions. Ornis Fenn. 1987, 64, 57–66. [Google Scholar]
- Shamovich, D.I.; Shamovich, I.Y. Spatial and Temporal Differences in the Association of Forest Owls in Conditions of Different Landscape Types of Northern Belarus. In Owls of the Northern Eurasia; Volkov, S.V., Morozov, V.V., Sharikov, A.V., Eds.; MPGU: Moscow, Russia, 2005; pp. 121–136. [Google Scholar]
- Sidorovich, V.E.; Shamovich, D.I.; Solovey, I.A.; Lauzhel, G.O. Dietary Variations of the Ural Owl Strix uralensis in the Transitional Mixed Forest of Northern Belarus with Implications for the Distribution Differences. Ornis Fenn. 2003, 80, 145–158. [Google Scholar]
- Tishechkin, A.K. Comparative Food Niche Analysis of Strix Owls in Belarus. In Biology and Conservation of Owls of the Northern Hemisphere; USDA Forest Service: Washington, DC, USA, 1997; pp. 456–460. [Google Scholar]
- Pietiäinen, H. Breeding Season Quality, Age, and the Effect of Experience on the Reproductive Success of the Ural Owl (Strix uralensis). Auk 1988, 105, 316–324. [Google Scholar]
- Brommer, J.E.; Pietiäinen, H.; Kolunen, H. The Effect of Age at First Breeding on Ural Owl Lifetime Reproductive Success and Fitness under Cyclic Food Conditions. J. Anim. Ecol. 1998, 67, 248–258. [Google Scholar] [CrossRef]
- Karell, P.; Pietiäinen, H.; Siitari, H.; Pihlaja, T.; Kontiainen, P.; Brommer, J.E. Parental Allocation of Additional Food to Own Health and Offspring Growth in a Variable Environment. Can. J. Zool. 2008, 87, 8–19. [Google Scholar] [CrossRef]
- Pietiäinen, H.; Saurola, P.; Väisänen, R.A.; Ornis, S.; Scandinavian, S.; Dec, N. Parental Investment in Clutch Size and Egg Size in the Ural Owl Strix uralensis. Ornis Scand. 1986, 17, 309–325. [Google Scholar] [CrossRef]
- Brommer, J.E.; Kontiainen, P.; Pietiäinen, H. Selection on Plasticity of Seasonal Life-History Traits Using: Random Regression Mixed Model Analysis. Ecol. Evol. 2012, 2, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Bylicka, M.; Kajtoch, Ł.; Figarski, T. Habitat and Landscape Characteristics Affecting the Occurrence of Ural Owls Strix uralensis in an Agroforestry Mosaic. Acta Ornithol. 2010, 45, 33–42. [Google Scholar] [CrossRef]
- Tutiš, V.; Radović, D.; Ćiković, D.; Barišić, S.; Kralj, J. Distribution, Density and Habitat Relationships of the Ural Owl Strix uralensis Macroura in Croatia. Ardea 2009, 97, 563–570. [Google Scholar] [CrossRef]
- Vrezec, A.; Mihelič, T. The Ural Owl, Strix Uralensis Macroura, in Slovenia: An Overview of Current Knowledge on Species Ecology. Riv. Ital. Ornitol. 2012, 82, 30–37. [Google Scholar] [CrossRef]
- Bashta, A.-T. Ural Owl Strix Uralensis Population Dynamics and Range Expansion in Western Ukraine. Ardea 2009, 97, 483–487. [Google Scholar] [CrossRef]
- Sonerud, G.A. Effect of Snow Cover on Seasonal Changes in Diet, Habitat and Regional Distribution of Raptors That Prey on Small Mammals in Boreal Zones of Fennoscandia. Holarct. Ecol. 1986, 9, 33–47. [Google Scholar] [CrossRef]
- Lõhmus, A. Do Ural Owls (Strix uralensis) Suffer from the Lack of Nest Sites in Managed Forests? Biol. Conserv. 2003, 110, 1–9. [Google Scholar] [CrossRef]
- Ķerus, V.; Dekants, A.; Auniņš, A.; Mārdega, I. Latvijas Ligzdojošo Putnu Atlanti 1980–2017; Latvijas Ornitoloģijas Biedrība: Riga, Latvia, 2021. [Google Scholar]
- Solonen, T.; Karhunen, J.; Kekkonen, J.A.; Kolunen, H.; Pietiäinen, H. Tawny Owl Prey Remains Indicate Differences in the Dynamics of Coastal and Inland Vole Populations in Southern Finland. Popul. Ecol. 2016, 58, 557–565. [Google Scholar] [CrossRef]
- Jędrzejewski, W.; Jędrzejewska, B.; Zub, K.; Andrzej, L.; Bystrowski, C. Resource Use by Tawny Owls Strix aluco in Relation to Rodent Fluctuations in Białowieża National Park, Poland. J. Avian Biol. 1994, 25, 308–318. [Google Scholar] [CrossRef]
- Goszczynski, J.; Jablonski, P.; Lesiński, G.; Romanowski, J. Variation in Diet of Tawny Owl Strix aluco L. along an Urbanization Gradient. Acta Ornithol. 1993, 27, 113–123. [Google Scholar]
- Lesiński, G.; Gryz, J.; Kowalski, M. Bat Predation by Tawny Owls Strix aluco in Differently Human Transformed Habitats. Ital. J. Zool. 2009, 76, 415–421. [Google Scholar] [CrossRef]
- Petty, S.J. Diet of Tawny Owls (Strix aluco) in Relation to Field Vole (Microtus agrestis) Abundance in a Conifer Forest in Northern England. J. Zool. 1999, 248, 451–465. [Google Scholar] [CrossRef]
- Romanowski, J.; Zmihorski, M. Seasonal and Habitat Variation in the Diet of the Tawny Owl (Strix aluco) in Central Poland during Unusually Warm Years. Biologia 2009, 64, 365–369. [Google Scholar] [CrossRef]
- Solonen, T. Are Vole-Eating Owls Affected by Mild Winters in Southern Finland? Ornis Fenn. 2004, 81, 65–74. [Google Scholar]
- Solonen, T.; Karhunen, J.; Kekkonen, J.; Kolunen, H.; Pietiäinen, H. Diet and Reproduction in Coastal and Inland Populations of the Tawny Owl Strix aluco in Southern Finland. J. Ornithol. 2017, 158, 541–548. [Google Scholar] [CrossRef]
- Solonen, T. Timing of Breeding in Rural and Urban Tawny Owls Strix aluco in Southern Finland: Effects of Vole Abundance and Winter Weather. J. Ornithol. 2014, 155, 27–36. [Google Scholar] [CrossRef]
- Solonen, T.; Ahola, K.; Karstinen, T. Clutch Size of a Vole-Eating Bird of Prey as an Indicator of Vole Abundance. Environ. Monit. Assess. 2015, 187, 588. [Google Scholar] [CrossRef]
- Karell, P.; Ahola, K.; Karstinen, T.; Valkama, J.; Brommer, J.E. Climate Change Drives Microevolution in a Wild Bird. Nat. Commun. 2011, 2, 208. [Google Scholar] [CrossRef]
- Solonen, T. Significance of Plumage Colour for Winter Survival in the Tawny Owl (Strix aluco): Revisiting the Camouflage Hypothesis. Ibis 2021, 163, 1437–1442. [Google Scholar] [CrossRef]
- Rumbutis, S.; Vaitkuvienė, D.; Grašytė, G.; Dagys, M.; Dementavičius, D.; Treinys, R. Adaptive Habitat Preferences in the Tawny Owl Strix aluco. Bird Study 2017, 64, 421–430. [Google Scholar] [CrossRef]
- Mitropolskiy, O.V.; Rustamov, A.K. Eagle Owl—Bubo Bubo Linnaeus, 1758. In The Birds of Middle Asia; Union of Protection of Birds of Kazakhstan: Almaty, Russia, 2007; pp. 423–431. [Google Scholar]
- Korpimäki, E.; Huhtala, K.; Sulkava, S. Does the Year-To-Year Variation in the Diet of Eagle and Ural Owls Support the Alternative Prey Hypothesis? Oikos 1990, 58, 47–54. [Google Scholar] [CrossRef]
- Randla, T. Eesti Röövlinnud [Estonian Birds of Prey]; Valgus: Tallinn, Estinia, 1976. [Google Scholar]
- Schweiger, A. Bubo Bubo Während Der Jungenaufzucht in Bayern. Ornitol. Anz. 2011, 50, 1–25. [Google Scholar]
- Frafjord, K. Population Dynamics of an Island Population of Water Voles Arvicola amphibius (Linnaeus, 1758) with One Major Predator, the Eagle Owl Bubo bubo (Linnaeus, 1758), in Northern Norway. Polar Biol. 2022, 45, 1–12. [Google Scholar] [CrossRef]
- BirdLife International. Species Factsheet: Bubo bubo. Available online: http://datazone.birdlife.org/species/factsheet/eurasian-eagle-owl-bubo-bubo (accessed on 8 December 2022).
- Penteriani, V.; Gallardo, M.; Roche, P. Landscape Structure and Food Supply Affect Eagle Owl (Bubo bubo) Density and Breeding Performance: A Case of Intra-Population Heterogeneity. J. Zool. 2002, 257, S0952836902000961. [Google Scholar] [CrossRef]
- Campioni, L.; Delgado, M.D.M.; Lourenço, R.; Bastianelli, G.; Fernández, N.; Penteriani, V. Individual and Spatio-Temporal Variations in the Home Range Behaviour of a Long-Lived, Territorial Species. Oecologia 2013, 172, 371–385. [Google Scholar] [CrossRef]
- Marchesi, L.; Sergio, F.; Pedrini, P. Costs and Benefits of Breeding in Human-Altered Landscapes for the Eagle Owl Bubo bubo. Ibis 2002, 144, E144–E177. [Google Scholar] [CrossRef]
- Karell, P.; Kontiainen, P.; Pietiäinen, H.; Siitari, H.; Brommer, J.E. Maternal Effects on Offspring Igs and Egg Size in Relation to Natural and Experimentally Improved Food Supply. Funct. Ecol. 2008, 22, 682–690. [Google Scholar] [CrossRef]
- Pietiäinen, H.; Kolunen, H. Female Body Condition and Breeding of the Ural Owl Strix uralensis. Funct. Ecol. 1993, 7, 726–735. [Google Scholar] [CrossRef]







| Sample Area | Period | Description |
|---|---|---|
| Apsalas | 1991–2011; 2015–2016 | 2 habitats: meadow and forest (OFD); 100 traps per transect |
| Lisiņa | 2001–2005 | 2 habitats: meadow and forest (OFU); 100 traps per transect |
| Žūklis | 2003–2011; 2015–2016 | 2 habitats: meadow and forest (OFD); 100 traps per transect |
| Ķemeri | 2006–2010; 2015–2016 | 2 habitats: meadow and forest (OFU); 100 traps per transect |
| Kaunata | 2012–2016 | 11 habitats: 1 meadow and 10 forest classes; 20–25 traps per transect |
| Lubāna | 2012–2016 | 11 habitats: 1 meadow and 10 forest classes; 20–25 traps per transect |
| Pļaviņas | 2012; 2016 | 11 habitats: 1 meadow and 10 forest classes; 20–25 traps per transect |
| Augstroze | 2012–2016 | 11 habitats: 1 meadow and 10 forest classes; 20–25 traps per transect |
| Owl Species | Prey (Index) | β ± SE | Test Statistic | df * | p-Value | AICc | R2adj./R2marg. ** | R2cond. | ICC |
|---|---|---|---|---|---|---|---|---|---|
| STRALU | Small mammals | −0.6127 ± 0.1850 | −3.312 | 144.778 | 0.0012 | 662.308 | 0.063 | 0.068 | 0.005 |
| Microtus voles | −0.3886 ± 0.1551 | −2.506 | 156.303 | 0.0132 | 667.001 | 0.037 | 0.049 | 0.013 | |
| Bank voles | −0.2268 ± 0.0812 | −2.795 | 161.449 | 0.0058 | 666.913 | 0.046 | 0.052 | 0.007 | |
| STRURA | Small mammals | −0.0039 ± 0.2514 | 0.015 | 54 | 0.9880 | 177.536 | −0.019 | ||
| Microtus voles | −0.1304 ± 0.1820 | −0.716 | 54 | 0.4770 | 177.007 | −0.009 | |||
| Bank voles | −0.0050 ± 0.0837 | −0. 06 | 54 | 0.9520 | 177.532 | −0.019 | |||
| GLAPAS | Small mammals | −0.4948 ± 0.7738 | −0.639 | 5 | 0.5507 | 39.803 | −0.109 | ||
| Microtus voles | −0.5290 ± 0.5286 | −1.001 | 5 | 0.3628 | 39.075 | 0.0003 | |||
| Bank voles | −0.4110 ± 0.2885 | −1.425 | 5 | 0.2136 | 37.967 | 0.147 | |||
| ASIOTU | Small mammals | 0.0171 ± 0.1531 | 0.112 | 22 | 0.9120 | 56.975 | −0.045 | ||
| Microtus voles | −0.0672 ± 0.1497 | −0.449 | 22 | 0.6580 | 56.769 | −0.036 | |||
| Bank voles | 0.0236 ± 0.0752 | 0.314 | 22 | 0.7570 | 56.881 | −0.041 |
| Owl Species | Prey (Index) * | β ± SE | Test Statistic | p-Value | AICc | R2MF/R2marg. ** | R2cond. | ICC |
|---|---|---|---|---|---|---|---|---|
| STRALU | Bank voles | 0.0248 ± 0.0057 | 4.359 | <0.0001 | 5693.822 | 0.0005 | 0.134 | 0.133 |
| Microtus voles | 0.1302 ± 0.0061 | 21.520 | <0.0001 | 14934.823 | 0.004 | 0.191 | 0.189 | |
| Bank~Microtus | −0.0138 ± 0.0116 | −1.189 | 0.2340 | 5711.112 | <0.0001 | 0.134 | 0.134 | |
| STRURA | Bank voles | −0.1116 ± 0.0072 | −15.570 | <0.0001 | 4693.187 | 0.0581 | ||
| Microtus voles | 0.1275 ± 0.0100 | 12.720 | <0.0001 | 6132.077 | 0.0268 | |||
| Bank~Microtus | −0.1658 ± 0.0175 | −9.497 | <0.0001 | 4865.001 | 0.0208 | |||
| GLAPAS | Bank voles | −0.4168 ± 0.0717 | −5.812 | <0.0001 | 261.048 | 0.178 | ||
| Microtus voles | 0.2136 ± 0.0556 | 3.839 | 0.0001 | 220.591 | 0.073 | |||
| Bank~Microtus | −0.9097 ± 0.0811 | −11.220 | <0.0001 | 145.446 | 0.613 | |||
| ASIOTU | Bank voles | −0.5294 ± 0.0497 | −10.640 | <0.0001 | 761.318 | 0.215 | ||
| Microtus voles | 0.0611 ± 0.0138 | 4.419 | <0.0001 | 599.700 | 0.044 | |||
| Bank~Microtus | −1.2718 ± 0.1208 | −10.530 | <0.0001 | 705.794 | 0.276 |
| Owl Species | Parameter | β ± SE | Test Statistic | p-Value | df * | R2adj. ** |
|---|---|---|---|---|---|---|
| STRALU | Intercept | 0.1178 ± 0.0396 | 2.978 | 0.0062 | 26 | −0.1096 |
| Time | 0.0016 ± 0.0043 | 0.368 | 0.7159 | |||
| Before | reference | |||||
| After | <0.0001 ± 0.0059 | <0.0001 | 1 | |||
| Time:After | <0.0001 ± 0.0006 | <0.0001 | 1 | |||
| STRURA | Intercept | 0.1389 ± 0.1361 | 1.020 | 0.319 | 21 | 0.5837 |
| Time | 0.0319 ± 0.0179 | 1.783 | 0.089 | |||
| Before | reference | |||||
| After | 1.1350 ± 0.1925 | 5.898 | <0.0001 | |||
| Time:After | −0.1354 ± 0.0253 | −5.361 | <0.0001 | |||
| GLAPAS *** | Intercept | 0.0069 ± 0.07470 | 0.093 | 0.9279 | 11 | 0.4368 |
| Time | −0.0339 ± 0.0106 | −3.210 | 0.0083 | |||
| ASIOTU | Intercept | −0.8450 ± 0.2688 | −3.144 | 0.0046 | 23 | −0.0302 |
| Time | −0.0143 ± 0.0328 | −0.436 | 0.6666 | |||
| Before | reference | |||||
| After | 0.3087 ± 0.3801 | 0.812 | 0.4250 | |||
| Time:After | −0.0080 ± 0.0464 | −0.172 | 0.8651 | |||
| AEGFUN | Intercept | −0.5246 ± 0.0331 | −15.855 | <0.0001 | 23 | 0.9740 |
| Time | −0.0607 ± 0.0040 | −15.033 | <0.0001 | |||
| Before | reference | |||||
| After | −0.7508 ± 0.0468 | −16.043 | <0.0001 | |||
| Time:After | 0.0183 ± 0.0057 | 3.211 | 0.0039 | |||
| Owl Species | Parameter | β ± SE | Test Statistic | p-Value | df * | R2adj. ** |
|---|---|---|---|---|---|---|
| STRALU | Intercept | 2.6221 ± 0.1594 | 16.452 | <0.0001 | 23 | 0.0765 |
| Time | 0.0174 ± 0.0194 | 0.896 | 0.380 | |||
| Before | reference | |||||
| After | −0.3597 ± 0.2254 | −1.596 | 0.124 | |||
| Time:After | 0.0216 ± 0.0275 | 0.784 | 0.441 | |||
| STRURA | Intercept | 1.8291 ± 0.1709 | 10.705 | <0.0001 | 21 | −0.0812 |
| Time | 0.0106 ± 0.0224 | 0.473 | 0.641 | |||
| Before | reference | |||||
| After | −0.2022 ± 0.2416 | −0.837 | 0.412 | |||
| Time:After | 0.0293 ± 0.0317 | 0.923 | 0.366 | |||
| Owl Species | Prey (Index) | rs | p-Value | Number of Years | S |
|---|---|---|---|---|---|
| STRALU | Small mammals | −0.1152 | 0.5737 | 26 | 3262 |
| Microtus voles | −0.1391 | 0.4962 | 26 | 3332 | |
| Bank voles | 0.0338 | 0.8700 | 26 | 2826 | |
| Small mammals−1 | 0.1300 | 0.5341 | 25 | 2262 | |
| Microtus voles−1 | 0.1377 | 0.5100 | 25 | 2242 | |
| Bank voles−1 | 0.1946 | 0.3496 | 25 | 2094 | |
| STRURA | Small mammals | −0.0179 | 0.9340 | 24 | 2341.1 |
| Microtus voles | 0.0545 | 0.8005 | 24 | 2174.8 | |
| Bank voles | −0.0863 | 0.6886 | 24 | 2498.4 | |
| Small mammals−1 | −0.1788 | 0.4145 | 23 | 2385.8 | |
| Microtus voles−1 | −0.2104 | 0.3351 | 23 | 2449.9 | |
| Bank voles−1 | 0.0218 | 0.9214 | 23 | 1979.9 | |
| ASIOTU | Small mammals | 0.0805 | 0.7755 | 15 | 514.92 |
| Microtus voles | 0.1252 | 0.6566 | 15 | 489.87 | |
| Bank voles | 0.0787 | 0.7804 | 15 | 515.92 | |
| Small mammals−1 | 0.3062 | 0.2871 | 14 | 315.69 | |
| Microtus voles−1 | 0.0529 | 0.8576 | 14 | 430.95 | |
| Bank voles−1 | −0.2643 | 0.3612 | 14 | 575.26 | |
| BUBBUB | Small mammals | 0.5329 | 0.0408 | 15 | 261.59 |
| Microtus voles | 0.3817 | 0.1604 | 15 | 346.28 | |
| Bank voles | 0.3402 | 0.2146 | 15 | 369.49 | |
| Small mammals−1 | 0.6438 | 0.0130 | 14 | 162.09 | |
| Microtus voles−1 | 0.5527 | 0.0404 | 14 | 203.50 | |
| Bank voles−1 | 0.2020 | 0.4886 | 14 | 363.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avotins, A.; Avotins, A., Sr.; Ķerus, V.; Aunins, A. Numerical Response of Owls to the Dampening of Small Mammal Population Cycles in Latvia. Life 2023, 13, 572. https://doi.org/10.3390/life13020572
Avotins A, Avotins A Sr., Ķerus V, Aunins A. Numerical Response of Owls to the Dampening of Small Mammal Population Cycles in Latvia. Life. 2023; 13(2):572. https://doi.org/10.3390/life13020572
Chicago/Turabian StyleAvotins, Andris, Andris Avotins, Sr., Viesturs Ķerus, and Ainars Aunins. 2023. "Numerical Response of Owls to the Dampening of Small Mammal Population Cycles in Latvia" Life 13, no. 2: 572. https://doi.org/10.3390/life13020572
APA StyleAvotins, A., Avotins, A., Sr., Ķerus, V., & Aunins, A. (2023). Numerical Response of Owls to the Dampening of Small Mammal Population Cycles in Latvia. Life, 13(2), 572. https://doi.org/10.3390/life13020572