

Rapid Onsite Visual Detection of Orf Virus Using a Recombinase-Aided Amplification Assay

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus and Clinical Samples of ORFV
2.2. Primers and Probe Design for RAA
2.3. Pyrolysis (DNA Extraction–Free) of Clinical Samples
2.4. Real-Time PCR and DNA Extraction–Free RAA Assay
2.5. Analytical Specificity
2.6. Analytical Sensitivity
2.7. Comparison of DNA Extraction–Free RAA with qPCR Using Clinical Samples
2.8. Statistical Analysis
3. Results
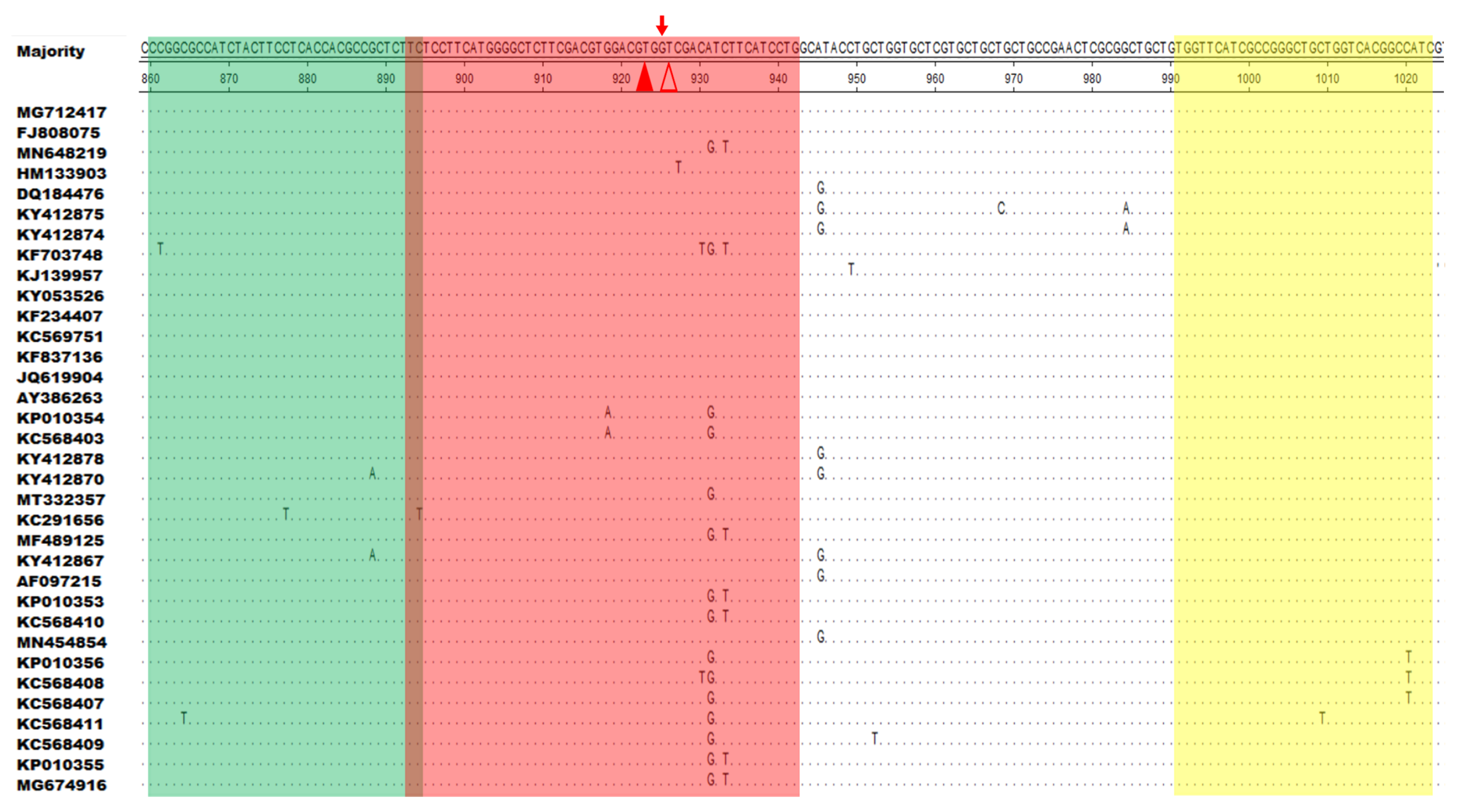
3.1. Positions of the RAA Primers and Probe
3.2. Screening the Optimal Primers for RAA Amplification
3.3. Specificity Analysis
3.4. Sensitivity Analysis
3.5. Diagnostic Sensitivity and Specificity of Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the evolution of host range and virulence. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2014, 21, 15–40. [Google Scholar] [CrossRef]
- Babkin, I.V.; Shchelkunov, S.N. The time scale in poxvirus evolution. Mol. Biol. 2006, 40, 20–24. [Google Scholar] [CrossRef]
- Brennan, G.; Stoian, A.M.M.; Yu, H.; Rahman, M.J.; Banerjee, S.; Stroup, J.N.; Park, C.; Tazi, L.; Rothenburg, S. Molecular Mechanisms of Poxvirus Evolution. mBio 2022, e0152622. [Google Scholar] [CrossRef] [PubMed]
- Spyrou, V.; Valiakos, G. Orf virus infection in sheep or goats. Vet. Microbiol. 2015, 181, 178–182. [Google Scholar] [CrossRef]
- Delhon, G.; Tulman, E.R.; Afonso, C.L.; Lu, Z.; de la Concha-Bermejillo, A.; Lehmkuhl, H.D.; Piccone, M.E.; Kutish, G.F.; Rock, D.L. Genomes of the parapoxviruses ORF virus and bovine papular stomatitis virus. J. Virol. 2004, 78, 168–177. [Google Scholar] [CrossRef]
- Gelaye, E.; Achenbach, J.E.; Jenberie, S.; Ayelet, G.; Belay, A.; Yami, M.; Loitsch, A.; Grabherr, R.; Diallo, A.; Lamien, C.E. Molecular characterization of orf virus from sheep and goats in Ethiopia, 2008–2013. Virol. J. 2016, 13, 34. [Google Scholar] [CrossRef]
- Chi, X.; Zeng, X.; Luo, S. Diagnosis and phylogenetic analysis of a multifocal cutaneous orf virus with mixed bacterial infection outbreak in goats in Fujian province, China. Arch. Virol. 2017, 162, 2997–3006. [Google Scholar] [CrossRef]
- Raele, D.A.; Stoffolano, J.G.; Vasco, I.; Pennuzzi, G.; La Porta, M.C.N.; Cafiero, M.A. Study on the Role of the Common House Fly, Musca domestica, in the Spread of ORF Virus (Poxviridae) DNA under Laboratory Conditions. Microorganisms 2021, 9, 2185. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, H.; Ghorani, M.; Arani, E.B.; Shakeri, A.P. Detection of contagious ovine ecthyma (orf) and risk factors for infection in small ruminants in Iran. Comp. Immunol. Microbiol. Infect. Dis. 2021, 79, 101714. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Yu, Z.; Li, Y.; Bai, C.; Sun, P.; Zhu, W.; Li, Y. Development of a recombinase-aided amplification assay for detection of orf virus. J. Virol. Methods 2020, 280, 113861. [Google Scholar] [CrossRef]
- Hosamani, M.; Scagliarini, A.; Bhanuprakash, V.; McInnes, C.J.; Singh, R.K. Orf: An update on current research and future perspectives. Expert. Rev. Anti Infect. Ther. 2009, 7, 879–893. [Google Scholar] [CrossRef]
- Abrahao, J.S.; Campos, R.K.; Trindade, G.S.; Guedes, M.I.; Lobato, Z.I.; Mazur, C.; Ferreira, P.C.; Bonjardim, C.A.; Kroon, E.G. Detection and phylogenetic analysis of Orf virus from sheep in Brazil: A case report. Virol. J. 2009, 6, 47. [Google Scholar] [CrossRef]
- Oem, J.K.; Roh, I.S.; Lee, K.H.; Lee, K.K.; Kim, H.R.; Jean, Y.H.; Lee, O.S. Phylogenetic analysis and characterization of Korean orf virus from dairy goats: Case report. Virol. J. 2009, 6, 167. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, K.; Wang, Y.A.; Yu, Z.; Zhang, Q.; Zhang, G.; Su, L.; Lu, Z.; Zhang, X.; Liu, Z.; et al. Identification and phylogenetic analysis of an orf virus strain isolated in Anhui Province, East-central China, in 2018. Acta Virol. 2019, 63, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Song, D.; He, W.; Lu, H.; Zhang, B.; Li, C.; Chen, K.; Gao, F. Identification and phylogenetic analysis of an Orf virus isolated from an outbreak in sheep in the Jilin province of China. Vet. Microbiol. 2010, 142, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yang, F.; Zhang, Q.; Xu, Y.; Huang, J.; Fu, M.; Zhang, W. Development of a multiplex TaqMan qPCR assay for simultaneous detection and differentiation of four DNA and RNA viruses from clinical samples of sheep and goats. J. Virol. Methods 2019, 266, 58–64. [Google Scholar] [CrossRef]
- Mullis, K.B.; Faloona, F.A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987, 155, 335–350. [Google Scholar] [CrossRef]
- Shakeeb, N.; Varkey, P.; Ajit, A. Human Saliva as a Diagnostic Specimen for Early Detection of Inflammatory Biomarkers by Real-Time RT-PCR. Inflammation 2021, 44, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Camargo, M.; Soto-De Leon, S.C.; Rojas-Villarraga, A.; Sanchez, R.; Jaimes, C.; Perez-Prados, A.; Patarroyo, M.E.; Patarroyo, M.A. The diagnostic performance of classical molecular tests used for detecting human papillomavirus. J. Virol. Methods 2012, 185, 32–38. [Google Scholar] [CrossRef]
- Nishi, K.; Isobe, S.; Zhu, Y.; Kiyama, R. Fluorescence-based bioassays for the detection and evaluation of food materials. Sensors 2015, 15, 25831–25867. [Google Scholar] [CrossRef]
- Saingam, P.; Li, B.; Yan, T. Use of amplicon sequencing to improve sensitivity in PCR-based detection of microbial pathogen in environmental samples. J. Microbiol. Methods 2018, 149, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, J.; Gai, Z.; Huo, S.; Zhu, J.; Li, J.; Wang, R.; Xing, S.; Shi, G.; Shi, F.; et al. Comparison between digital PCR and real-time PCR in detection of Salmonella typhimurium in milk. Int. J. Food Microbiol. 2018, 266, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yu, M.; Dong, G.; Chen, B.; Zhang, B. Digital PCR as an Emerging Tool for Monitoring of Microbial Biodegradation. Molecules 2020, 25, 706. [Google Scholar] [CrossRef]
- Yan, L.; Zhou, J.; Zheng, Y.; Gamson, A.S.; Roembke, B.T.; Nakayama, S.; Sintim, H.O. Isothermal amplified detection of DNA and RNA. Mol. Biosyst. 2014, 10, 970–1003. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Ge, J.; Guo, Y. Recent advances in methods for the diagnosis of Corona Virus Disease 2019. J. Clin. Lab. Anal. 2022, 36, e24178. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, C.; Tu, F.; Zhao, K.; Kong, Y.; Pu, J.; Zhang, L.; Chen, Z.; Sun, Y.; Wei, Y.; et al. Rapid detection of influenza A viruses using a real-time reverse transcription recombinase-aided amplification assay. Front. Cell. Infect. Microbiol. 2023, 12, 1950. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, F.; Li, Q.; Wang, L.; Fan, C. Isothermal Amplification of Nucleic Acids. Chem. Rev. 2015, 115, 12491–12545. [Google Scholar] [CrossRef]
- Bodulev, O.L.; Sakharov, I.Y. Isothermal Nucleic Acid Amplification Techniques and Their Use in Bioanalysis. Biochemistry. Biokhimiia 2020, 85, 147–166. [Google Scholar] [CrossRef]
- Walker, G.T.; Little, M.C.; Nadeau, J.G.; Shank, D.D. Isothermal in vitro amplification of DNA by a restriction enzyme/DNA polymerase system. Proc. Natl. Acad. Sci. USA 1992, 89, 392–396. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.Q. Rolling replication of short DNA circles. Proc. Natl. Acad. Sci. USA 1995, 92, 4641–4645. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed]
- Vincent, M.; Xu, Y.; Kong, H. Helicase-dependent isothermal DNA amplification. EMBO Rep. 2004, 5, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef]
- James, A.; Macdonald, J. Recombinase polymerase amplification: Emergence as a critical molecular technology for rapid, low-resource diagnostics. Expert Rev. Mol. Diagn. 2015, 15, 1475–1489. [Google Scholar] [CrossRef]
- Yeh, E.C.; Fu, C.C.; Hu, L.; Thakur, R.; Feng, J.; Lee, L.P. Self-powered integrated microfluidic point-of-care low-cost enabling (SIMPLE) chip. Sci. Adv. 2017, 3, e1501645. [Google Scholar] [CrossRef]
- Shen, X.X.; Qiu, F.Z.; Shen, L.P.; Yan, T.F.; Zhao, M.C.; Qi, J.J.; Chen, C.; Zhao, L.; Wang, L.; Feng, Z.S.; et al. A rapid and sensitive recombinase aided amplification assay to detect hepatitis B virus without DNA extraction. BMC Infect. Dis. 2019, 19, 229. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, G.; Bhanuprakash, V.; Balamurugan, V. Development and comparative evaluation of loop mediated isothermal amplification (LAMP) assay for simple visual detection of orf virus in sheep and goats. Mol. Cell. Probes 2015, 29, 193–195. [Google Scholar] [CrossRef]
- Fan, X.; Li, L.; Zhao, Y.; Liu, Y.; Liu, C.; Wang, Q.; Dong, Y.; Wang, S.; Chi, T.; Song, F.; et al. Clinical Validation of Two Recombinase-Based Isothermal Amplification Assays (RPA/RAA) for the Rapid Detection of African Swine Fever Virus. Front. Microbiol. 2020, 11, 1696. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Lin, Z.; Huang, X.; Lu, J.; Zhou, Y.; Zheng, L.; Lou, Y. Rapid and Sensitive Detection of Vibrio vulnificus Using CRISPR/Cas12a Combined with a Recombinase-Aided Amplification Assay. Front. Microbiol. 2021, 12, 767315. [Google Scholar] [CrossRef]
- Wei, N.; Zheng, B.; Niu, J.; Chen, T.; Ye, J.; Si, Y.; Cao, S. Rapid Detection of Genotype II African Swine Fever Virus Using CRISPR Cas13a-Based Lateral Flow Strip. Viruses 2022, 14, 179. [Google Scholar] [CrossRef]
- Wang, Z.H.; Li, P.; Lin, X.; Jia, H.; Jiang, Y.T.; Wang, X.J.; Hou, S.H. Application of portable real-time recombinase-aided amplification (rt-RAA) assay in the clinical diagnosis of ASFV and prospective DIVA diagnosis. Appl. Microbiol. Biotechnol. 2021, 105, 3249–3264. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Zhang, Y.; Zeng, S.; Liu, X.; Li, Y.; Li, X.; Chen, W.; Li, Z.; Qin, Y.; Chen, J.; et al. Development and Application of RAA Nucleic Acid Test Strip Assay and Double RAA Gel Electrophoresis Detection Methods for ASFV and CSFV. Front. Mol. Biosci. 2021, 8, 811824. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Chen, Y.; Zhang, Y.; Yang, H. Development of a VP2-based real-time fluorescent reverse transcription recombinase-aided amplification assay to rapidly detect Senecavirus A. Transbound. Emerg. Dis. 2022, 69, 2828–2839. [Google Scholar] [CrossRef]
- Nie, M.Z.; Zhang, R.Q.; Zhao, M.C.; Tan, H.; Hu, Y.X.; Fan, G.H.; Li, J.Y.; He, A.N.; Tian, F.Y.; Li, F.Y.; et al. Development of a duplex recombinase-aided amplification assay for direct detection of Mycoplasma pneumoniae and Chlamydia trachomatis in clinical samples. J. Microbiol. Methods 2022, 198, 106504. [Google Scholar] [CrossRef]
- Lin, H.; Liang, Y.; Zou, L.; Li, B.; Zhao, J.; Wang, H.; Sun, J.; Deng, X.; Tang, S. Combination of Isothermal Recombinase-Aided Amplification and CRISPR-Cas12a-Mediated Assay for Rapid Detection of Major Severe Acute Respiratory Syndrome Coronavirus 2 Variants of Concern. Front. Microbiol. 2022, 13, 945133. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, W.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Chen, Y.; Zhang, Y.; Yang, H. A nucleic acid detection assay combining reverse transcription recombinase-aided amplification with a lateral flow dipstick for the rapid visual detection of porcine deltacoronavirus. Virulence 2022, 13, 1471–1485. [Google Scholar] [CrossRef]
- Zhong, J.; Guan, J.; Zhou, Y.; Cui, S.; Wang, Z.; Zhou, S.; Xu, M.; Wei, X.; Gao, Y.; Zhai, S.; et al. Genomic characterization of two Orf virus isolates from Jilin province in China. Virus Genes 2019, 55, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Tu, F.; Yang, X.; Xu, S.; Chen, D.; Zhou, L.; Ge, X.; Han, J.; Zhang, Y.; Guo, X.; Yang, H. Development of a fluorescent probe-based real-time reverse transcription recombinase-aided amplification assay for the rapid detection of classical swine fever virus. Transbound. Emerg. Dis. 2021, 68, 2017–2027. [Google Scholar] [CrossRef]
- Gallina, L.; Dal Pozzo, F.; Mc Innes, C.J.; Cardeti, G.; Guercio, A.; Battilani, M.; Ciulli, S.; Scagliarini, A. A real time PCR assay for the detection and quantification of orf virus. J. Virol. Methods 2006, 134, 140–145. [Google Scholar] [CrossRef]
- Tsai, S.M.; Chan, K.W.; Hsu, W.L.; Chang, T.J.; Wong, M.L.; Wang, C.Y. Development of a loop-mediated isothermal amplification for rapid detection of orf virus. J. Virol. Methods 2009, 157, 200–204. [Google Scholar] [CrossRef]
- Tu, F.; Zhang, Y.; Xu, S.; Yang, X.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Yang, H. Detection of pseudorabies virus with a real-time recombinase-aided amplification assay. Transbound. Emerg. Dis. 2022, 69, 2266–2274. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Li, W.J.; Li, X.M.; Fan, Q.L.; Tang, X.D.; Liu, M.J.; Ma, W.T.; Chen, D.K. Pathogenicity of blood orf virus isolates in the development of dairy goat contagious pustular dermatitis. Vet. Microbiol. 2018, 219, 178–182. [Google Scholar] [CrossRef]
- He, Y.; Xie, T.; Tong, Y. Rapid and highly sensitive one-tube colorimetric RT-LAMP assay for visual detection of SARS-CoV-2 RNA. Biosens. Bioelectron. 2021, 187, 113330. [Google Scholar] [CrossRef]
- Cui, H.; Tu, F.; Zhang, C.; Zhang, C.; Zhao, K.; Liu, J.; Dong, S.; Chen, L.; Liu, J.; Guo, Z. Real-Time Reverse Transcription Recombinase-Aided Amplification Assay for Rapid Amplification of the N Gene of SARS-CoV-2. Int. J. Mol. Sci. 2022, 23, 15269. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, S.E.; Bathrick, A.S. Direct PCR amplification of forensic touch and other challenging DNA samples: A review. Forensic Sci. Int. Genet. 2018, 32, 40–49. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, K.; Bai, C.; Yin, D.; Li, G.; Qi, K.; Wang, G.; Li, Y. Development of a SYBR Green I real-time PCR for the detection of the orf virus. AMB Express 2017, 7, 21. [Google Scholar] [CrossRef]
- Du, H.; Li, W.; Hao, W.; Liao, X.; Li, M.; Luo, S. Taqman real-time PCR assay based on ORFV024 gene for rapid detection of orf infection. Toxicol. Mech. Methods 2013, 23, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Shang, Y.; Wang, Y.; Tian, H.; Liu, X. Comparison of a loop-mediated isothermal amplification for orf virus with quantitative real-time PCR. Virol. J. 2013, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qin, X.; Wang, G.; Zhang, Y.; Shang, Y.; Zhang, Z. Development of a fluorescent probe-based recombinase polymerase amplification assay for rapid detection of Orf virus. Virol. J. 2015, 12, 206. [Google Scholar] [CrossRef]
- Yang, Y.; Qin, X.; Wang, G.; Jin, J.; Shang, Y.; Zhang, Z. Development of an isothermoal amplification-based assay for rapid visual detection of an Orf virus. Virol. J. 2016, 13, 46. [Google Scholar] [CrossRef]
- Yan, T.F.; Li, X.N.; Wang, L.; Chen, C.; Duan, S.X.; Qi, J.J.; Li, L.X.; Ma, X.J. Development of a reverse transcription recombinase-aided amplification assay for the detection of coxsackievirus A10 and coxsackievirus A6 RNA. Arch. Virol. 2018, 163, 1455–1461. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences (5′→3′) | Gene | Amplicon Size | Source |
|---|---|---|---|---|
| F860–894 | CCGGCGCCATCTACTTCCTCACCACGCCGCTCTTC | F1L | 164 bp | This study |
| R991–1023 | GATGGCCGTGACCAGCAGCCCGGCGATGAACCA | F1L | This study | |
| P893–942 a | TCTCCTTCATGGGGCTCTTCGACGTGGACG(FAM-dT)G(THF)(BHQ1-dT)CGACGTCTTCATCCTG[C3-spacer] | F1L | This study | |
| F | CAGCAGAGCCGCGTGAA | F1L | 70 bp | [49] |
| R | CATGAACCGCTACAACACCTTCT | F1L | [49] | |
| Probe b | FAM-CACCTTCGGCTCCAC-MGB | F1L | [49] |
| Assay | qPCR | Sensitivity | Specificity | Kappa | ||
|---|---|---|---|---|---|---|
| Positive | Negative | |||||
| RAA (Via real-time fluorescence read-out) | Positive | 39 | 0 | 100%(39/39) | 100%(39/39) | 1 |
| Negative | 0 | 39 | ||||
| RAA (Via visual detection) | Positive | 36 | 0 | 92.31%(36/39) | 100%(39/39) | 0.923 |
| Negative | 3 | 39 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, H.; Guan, J.; Lu, H.; Liu, J.; Tu, F.; Zhang, C.; Su, K.; Guo, Z.; Zhao, K. Rapid Onsite Visual Detection of Orf Virus Using a Recombinase-Aided Amplification Assay. Life 2023, 13, 494. https://doi.org/10.3390/life13020494
Cui H, Guan J, Lu H, Liu J, Tu F, Zhang C, Su K, Guo Z, Zhao K. Rapid Onsite Visual Detection of Orf Virus Using a Recombinase-Aided Amplification Assay. Life. 2023; 13(2):494. https://doi.org/10.3390/life13020494
Chicago/Turabian StyleCui, Huan, Jiyu Guan, Huijun Lu, Jun Liu, Fei Tu, Cheng Zhang, Kai Su, Zhendong Guo, and Kui Zhao. 2023. "Rapid Onsite Visual Detection of Orf Virus Using a Recombinase-Aided Amplification Assay" Life 13, no. 2: 494. https://doi.org/10.3390/life13020494
APA StyleCui, H., Guan, J., Lu, H., Liu, J., Tu, F., Zhang, C., Su, K., Guo, Z., & Zhao, K. (2023). Rapid Onsite Visual Detection of Orf Virus Using a Recombinase-Aided Amplification Assay. Life, 13(2), 494. https://doi.org/10.3390/life13020494