

Suppression of Lipid Accumulation in the Differentiation of 3T3-L1 Preadipocytes and Human Adipose Stem Cells into Adipocytes by TAK-715, a Specific Inhibitor of p38 MAPK
 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. 3T3-L1 Cells or hASCs Culture and Differentiation
2.3. Oil Red O Staining
2.4. Cell Count Analysis
2.5. Measurement of Intracellular TG Content
2.6. Preparation of Whole-Cell Lysates
2.7. Westerm Blotting Analysis
2.8. RT-qPCR
2.9. Statistical Analysis
3. Results
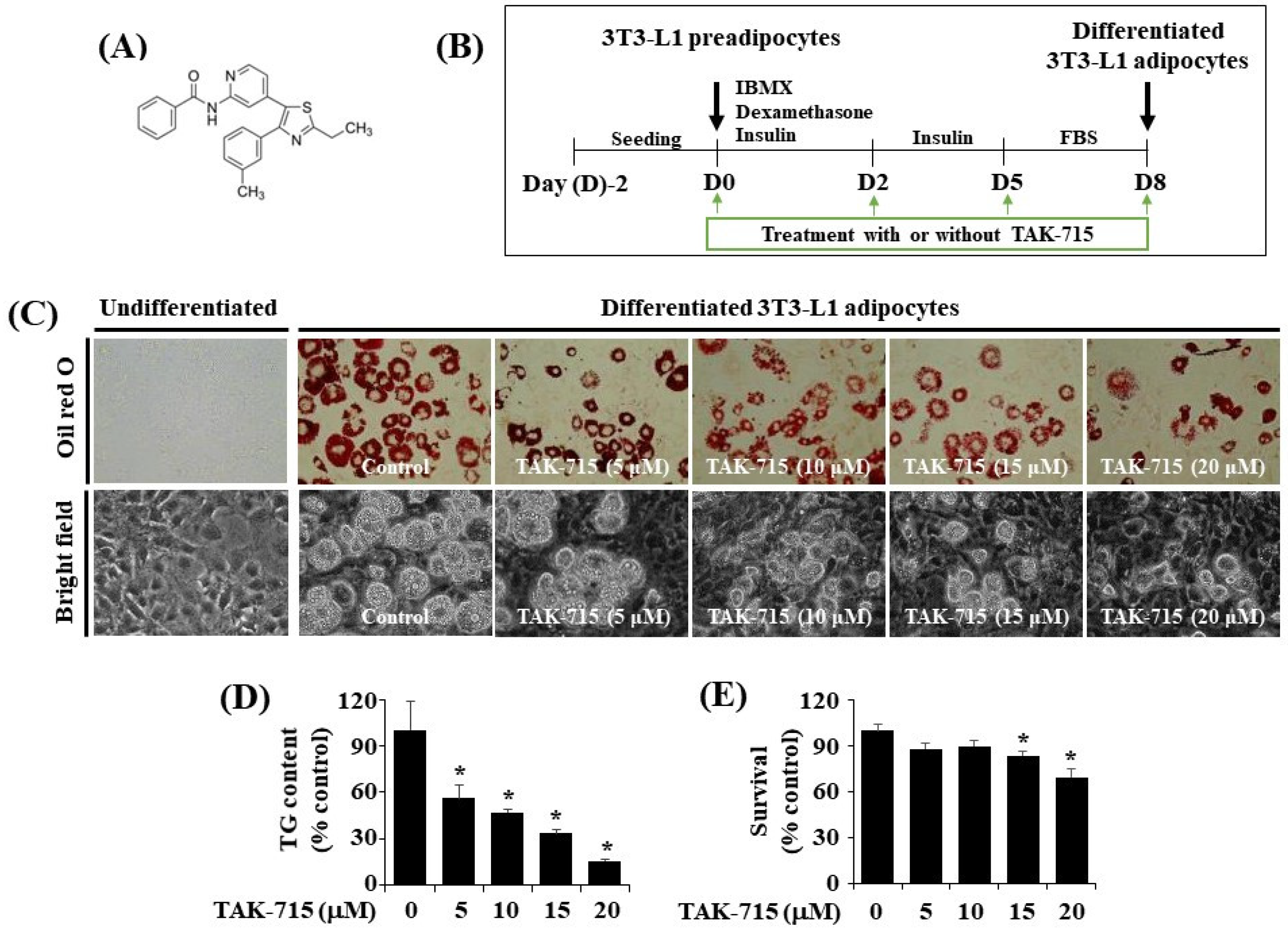
3.1. TAK-715 at 10 μM Markedly Suppresses Intracelullar Lipid Accumulation and TG Content during the Adipogenesis of 3T3-L1 Preadipocyte
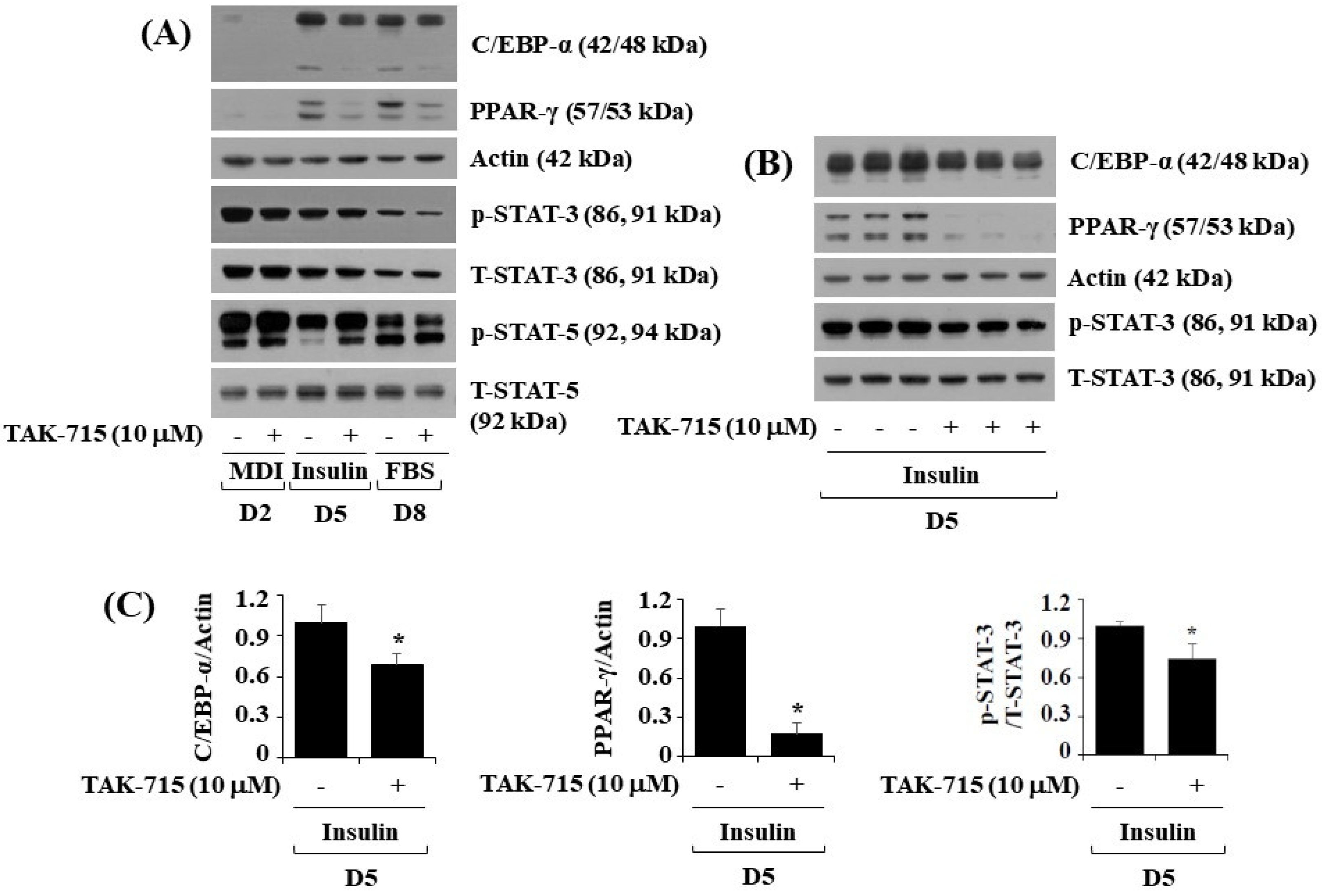
3.2. TAK-715 Significantly Reduces the Expression and Phosphorylation Levels of C/EBP-α, PPAR-γ, and STAT-3 in 3T3-L1 Cells Following Differentiation
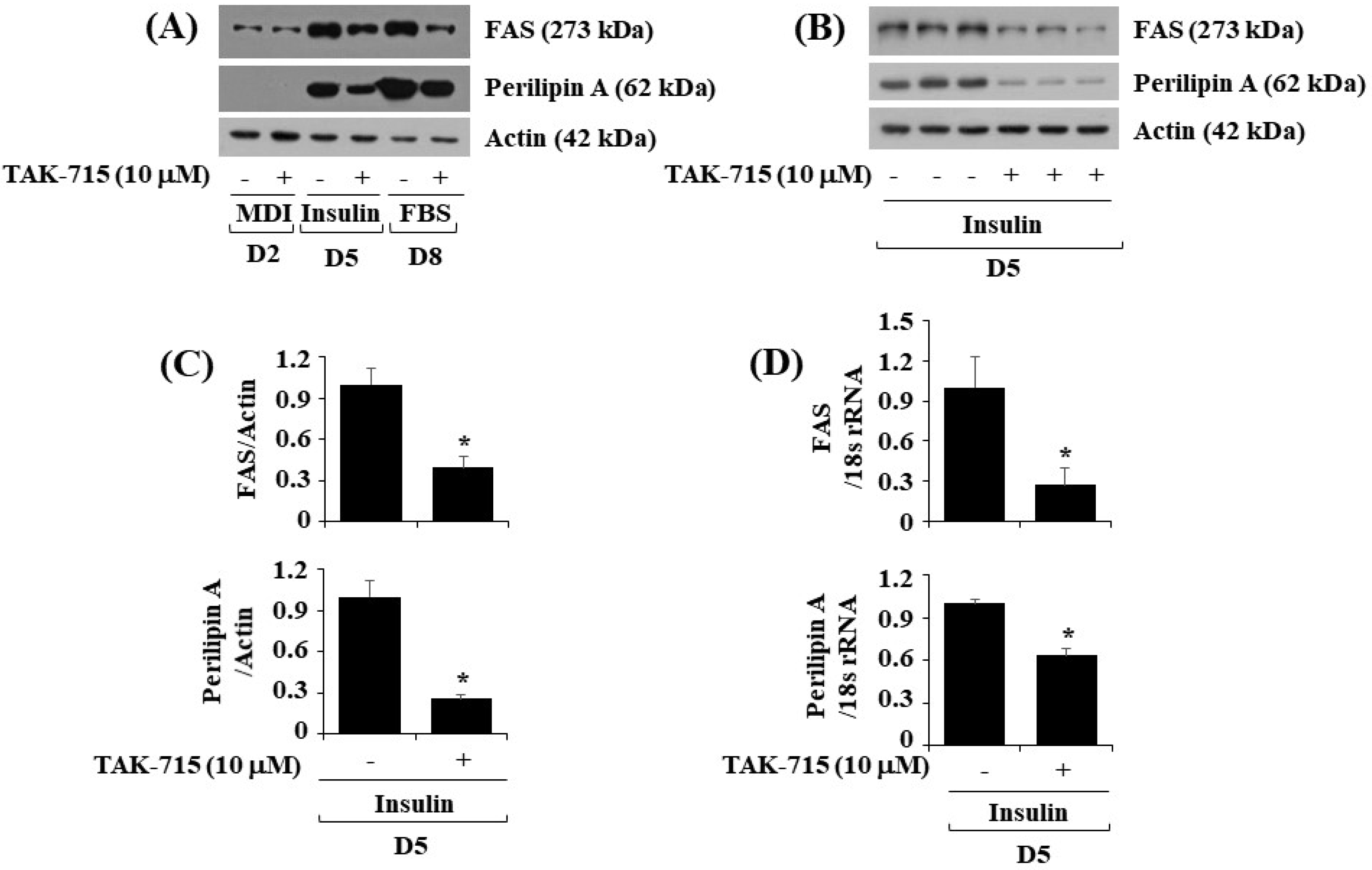
3.3. TAK-715 Significantly Lower the Expressions of FAS and Perilipin A during Preadipocyte Differentiation in 3T3-L1 Cells
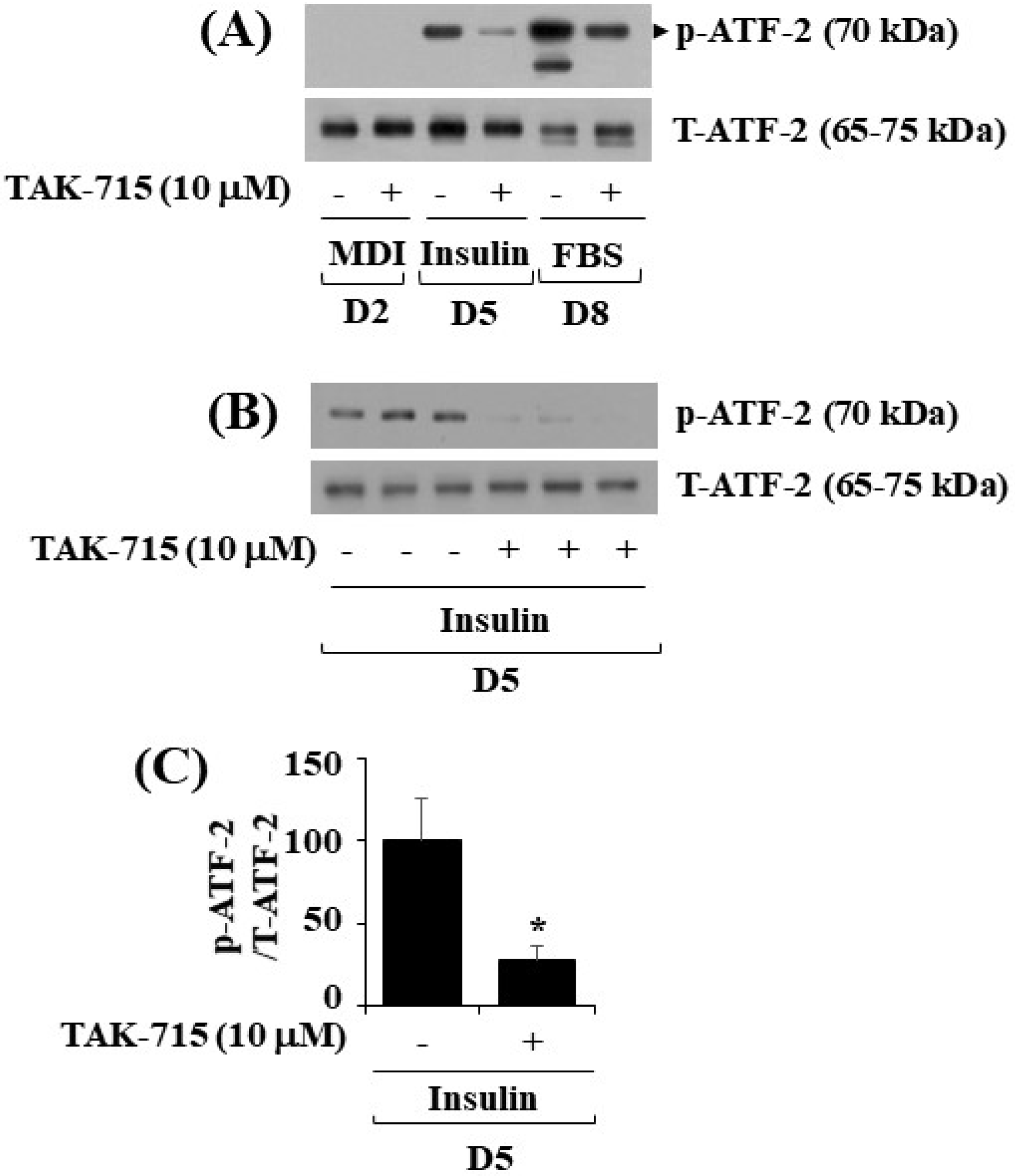
3.4. TAK-715 Significantly Reduces the Phosphorylation Levels of ATF-2, a p38 MAPK Downstream Effector, in Differentiating 3T3-L1 Preadipocyte
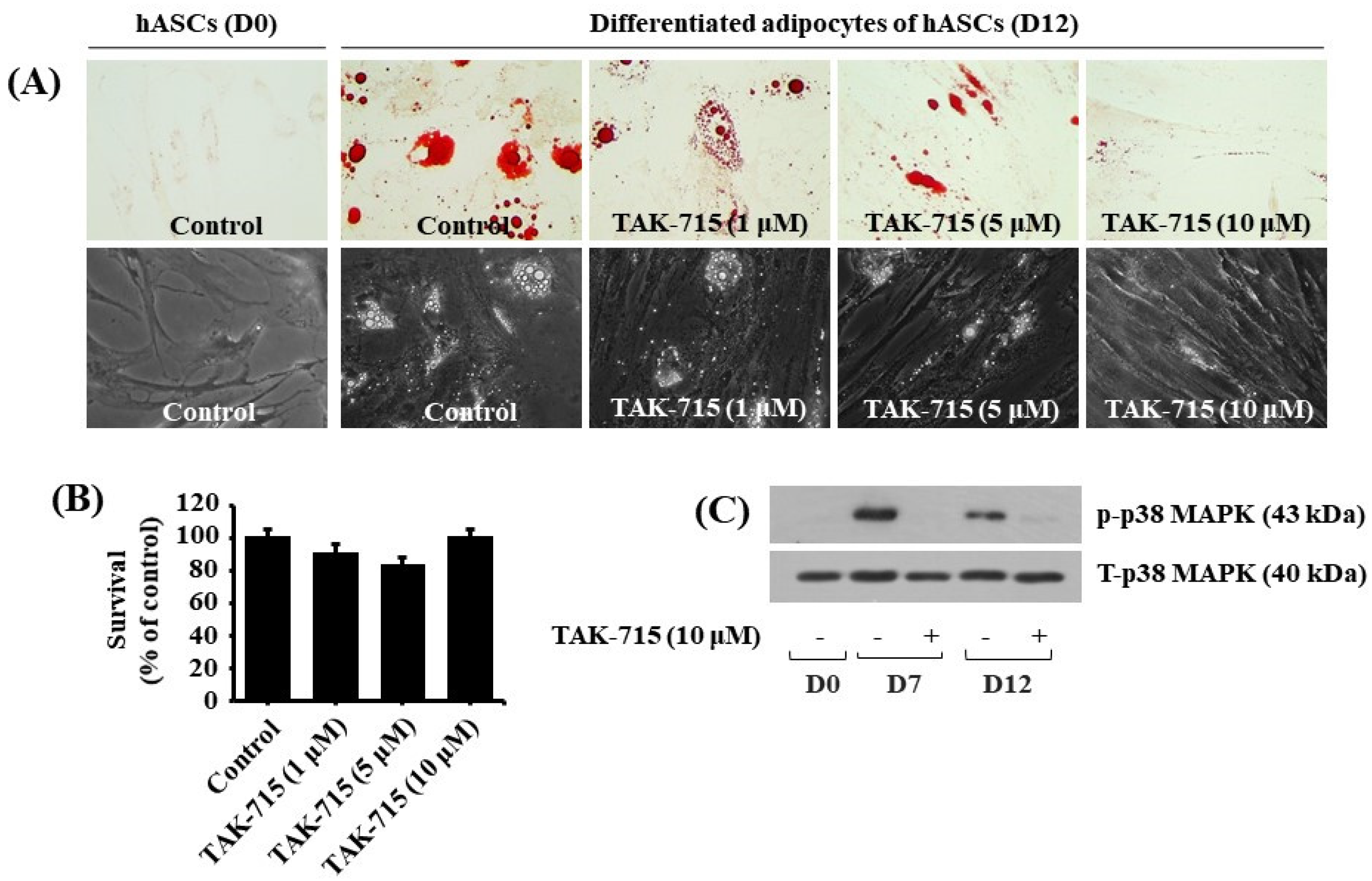
3.5. TAK-715 Markedly Reduces Lipid Formation and p38 MAPK Phosphorylation in hASCs Adipocyte Differentiation
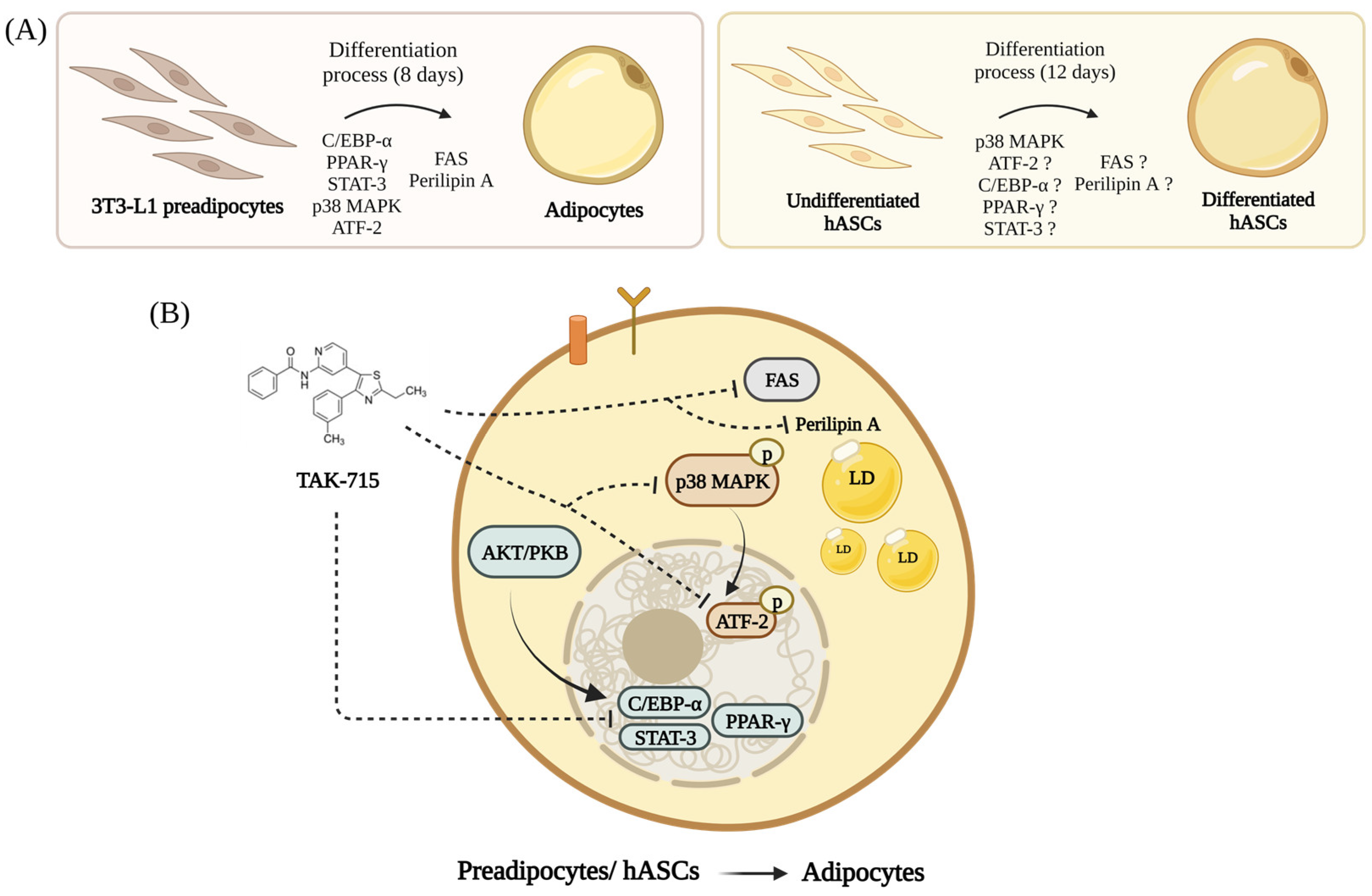
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sam, S.; Mazzone, T. Adipose tissue changes in obesity and impact on metabolic function. Transl. Res. 2014, 164, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Jakab, J.; Miškić, B.; Mikšić, S.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes Metab. Syndr. Obes. 2021, 14, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, S.; Mozaffarian, D. The perfect storm: Obesity, adipocyte dysfunction, and metabolic consequences. Clin. Chem. 2008, 54, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Lee, P.; Chisholm, D.J.; James, D.E. Control of adipocyte differentiation in different fat depots; implications for pathophysiology or therapy. Front. Endocrinol. 2015, 6, 1–8. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef]
- Khalilpourfarshbafi, M.; Gholami, K.; Murugan, D.D.; Sattar, M.Z.A.; Abdullah, N.A. Differential effects of dietary flavonoids on adipogenesis. Eur. J. Nutr. 2019, 58, 5–25. [Google Scholar] [CrossRef]
- Lazar, A.D.; Dinescu, S.; Costache, M. Adipose tissue engineering and adipogenesis—A review. Rev. Biol. Biomed. Sci. 2018, 1, 17–26. [Google Scholar] [CrossRef]
- Stephens, J.M.; Morrison, P.F.; Pilch, P.F. The expression and regulation of STATs during 3T3-L1 adipocyte differentiation. J. Biol. Chem. 1996, 271, 10441–10444. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.; Coligan, J.E.; Shuman, J.D. Activating transcription factor-2 regulates phosphoenolpyruvate carboxykinase transcription through a stress-inducible mitogen-activated protein kinase pathway. J. Biol. Chem. 1998, 273, 22714–22718. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Jung, C.H.; Lee, K.; Choi, Y.H.; Hong, S.H.; Cheong, J.H. Activating transcription factor-2 mediates transcriptional regulation of gluconeogenic gene PEPCK by retinoic acid. Diabetes 2002, 51, 3400–3407. [Google Scholar] [CrossRef]
- Okamura, T.; Shimizu, H.; Nagao, T.; Ueda, R.; Ishii, S. ATF-2 regulates fat metabolism in Drosophila. Mol. Biol. Cell. 2007, 18, 1519–1529. [Google Scholar] [CrossRef]
- Miwatashi, S.; Arikawa, Y.; Kotani, E.; Miyamoto, M.; Naruo, K.I.; Kimura, H.; Tanaka, T.; Asahi, S.; Ohkawa, S. Novel inhibitor of p38 MAP kinase as an anti-TNF-α drug: Discovery of N-[4-[2-Ethyl-4-(3-methylphenyl)-1,3-thiazol-5-yl]-2-pyridyl]benzamide (TAK-715) as a potent and orally active anti-rheumatoid arthritis agent. J. Med. Chem. 2005, 48, 5966–5979. [Google Scholar] [CrossRef]
- Veerkar, F.; van der Doelen, A.A.; Smits, J.F.M.; Blankestejin, W.M.; Zaman, G.J.R. Inhibition of Wnt/β-Catenin signaling by p38 MAP kinase inhibitors is explained by cross-reactivity with casein kinase Iδ/ε. Chem. Biol. 2011, 18, 485–494. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, H.; Liu, J.; Tse, C.A.; Dragunow, M.; Cooper, G.J.S. Activation of activating transcription factor 2 by p38 MAP kinase during apoptosis induced by human amylin in cultured pancreatic β-cells. FEBS J. 2006, 273, 3779–3791. [Google Scholar] [CrossRef]
- Breitwieser, W.; Lyons, S.; Flenniken, A.M.; Ashton, G.; Bruder, G.; Willington, M.; Lacaud, G.; Kouskoff, V.; Jones, N. Feedback regulation of p38 activity ATF2 is essential for survival of embryonic liver cells. Genes Dev. 2007, 21, 2069–2082. [Google Scholar] [CrossRef]
- Zhao, J.; Ma, J.; Lu, J.; Jiang, Y.; Zhang, Y.; Zhang, X.; Zhao, J.; Yang, H.; Huang, Y.; Zhao, M.; et al. Involvement of p38MAPK-ATF2 signaling pathway in alternariol induced DNA polymerase β expression. Oncol. Lett. 2017, 12, 675–679. [Google Scholar] [CrossRef]
- Maekawa, T.; Jin, W.; Ishii, S. The role of ATF-2 family transcription factors in adipocyte differentiation: Antiobesity effects of p38 inhibitors. Mol. Cell Biol. 2010, 30, 613–625. [Google Scholar] [CrossRef]
- Kayali, A.G.; Austin, D.A.; Webster, N.J. Stimulation of MAPK cascades by insulin and osmotic shock: Lack of an involvement of p38 mitogen-activated protein kinase in glucose transport in 3T3-L1 adipocytes. Diabetes 2000, 49, 1783–1793. [Google Scholar] [CrossRef]
- Fujishiro, M.; Gotoh, Y.; Katagiri, H.; Sakoda, H.; Ogihara, T.; Anai, M.; Onishi, Y.; Ono, H.; Funaki, M.; Inukai, K.; et al. MKK6/3 and p38 MAPK pathway activation is not necessary for insulin-induced glucose uptake but regulates glucose transporter expression. J. Biol. Chem. 2001, 276, 19800–19806. [Google Scholar] [CrossRef]
- Engelman, J.A.; Lisanti, M.P.; Scherer, P.E. Specific inhibitors of p38 mitogen-activated protein kinase block 3T3–L1 adipogenesis. J. Biol. Chem. 1998, 273, 32111–32120. [Google Scholar] [CrossRef]
- Takenouchi, T.; Takayama, Y.; Takezawa, T. Co-treatment with dexamethasone and octanoate induces adipogenesis in 3T3–L1 cells. Cell Biol. Int. 2004, 28, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Z.; Ron, D. Stress-induced phosphorylation and activation of the transcription factor CHOP (GADD153) by p38 MAP Kinase. Science 1996, 272, 1347–1349. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Guo, W.; Yang, Y. JAK2/STAT3 pathway is involved in the early stage of adipogenesis through regulating C/EBPβ transcription. J. Cell. Biochem. 2011, 112, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Xi, Y.; Chen, J.; Zhu, P.; Kang, J.; Zou, Z.; Wang, F.; Bu, S. STAT3 stimulates adipogenic stem cell proliferation and cooperates with HMGA2 during the early stage of differentiation to promote adipogenesis. Biochem. Biophys. Res. Commun. 2017, 482, 1360–1366. [Google Scholar] [CrossRef]
- Kadye, R.; Stoffels, M.; Fanucci, S.; Mbanxa, S.; Prinsloo, E. A STAT3 of addiction: Adipose tissue, adipocytokine signalling and STAT3 as mediators of metabolic remodelling in the tumour microenvironment. Cells 2020, 9, 1043. [Google Scholar] [CrossRef]
- Schmid, B.; Rippmann, J.F.; Tadayyon, M.; Hamilton, B.S. Inhibition of fatty acid synthase prevents preadipocyte differentiation. Biochem. Biophys. Res. Commun. 2005, 328, 1073–1082. [Google Scholar] [CrossRef]
- Tansey, J.T.; Sztalryd, C.; Hlavin, E.M.; Kimmel, A.R.; Londos, C. The central role of perilipin A in lipid metabolism and adipocyte lipolysis. IUBMB Life 2004, 56, 379–385. [Google Scholar] [CrossRef]
- Giménez-Andrés, M.; Emeršič, T.; Antoine-Bally, S.; D’Ambrosio, J.M.; Antony, B.; Derganc, J.; Čopič, A. Exceptional stability of a perilipin on lipid droplets depends on its polar residues, suggesting multimeric assembly. eLife 2021, 10, e61401. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kong, H.J.; Cheong, J.H. Regulation of activating transcription factor-2 in early stage of the adipocyte differentiation program. Biochem. Biophys. Res. Commun. 2001, 281, 1241–1247. [Google Scholar] [CrossRef]
- Aouadi, M.; Laurent, K.; Prot, M.; Brustel, Y.L.M.; Binétruy, Y.; Bost, F. Inhibition of p38MAPK increases adipogenesis from embryonic to adult stages. Diabetes 2006, 55, 281–289. [Google Scholar] [CrossRef]
- Ando, Y.; Sato, F.; Fukunaga, H.; Iwasaki, Y.; Chiba, Y.; Tebakari, M.; Daigo, Y.; Kawashima, J.; Kamei, J. Placental extract suppresses differentiation of 3T3-L1 preadipocytes to mature adipocytes via accelerated activation of p38 MAPK during the early phase of adipogenesis. Nutr. Metab. 2019, 16, 1–13. [Google Scholar] [CrossRef]
- de Winter, T.J.J.; Nusse, R. Running against the Wnt/β-catenin suppresses adipogenesis. Front. Cell Dev. Biol. 2021, 9, 627429. [Google Scholar] [CrossRef]
- Leiva, M.; Matesanz, N.; Pulgarn-Alfaro, M.; Nikolic, I.; Sabio, G. Uncovering the role of p38 family members in adipose tissue tissue physiology. Front. Endocrinol. 2020, 11, 572089. [Google Scholar] [CrossRef]
- Bose, C.; Bhuvaneswaran, C.; Udupa, K.B. Age-related alteration in hepatic acyl-CoA: Cholesterol acyltransferase and its relation to LDL receptor and MAPK. Mech. Ageing Dev. 2005, 126, 740–751. [Google Scholar] [CrossRef]
- Hai, Q.; Smith, J.D. Acyl-coenzyme A: Cholesterol Acyltransferase (ACAT) in cholesterol metabolism: From its discovery to clinical trials and the genomics era. Metabolites 2021, 11, 543. [Google Scholar] [CrossRef]
- Xu, Y.; Du, X.; Turner, N.; Brown, A.J.; Yang, H. Enhanced acyl-CoA: Cholesterol acyltransferase activity increases cholesterol levels on the lipid droplet surface and impairs adipocyte function. J. Biol. Chem. 2019, 294, 19306–19321. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Dilution Used | Source | Catalog No. |
|---|---|---|---|
| Primary Antibodies | |||
| C/EBP-α | 1:2000 | Santa Cruz Biotechnology | sc-61 |
| PPAR-γ | 1:2000 | Santa Cruz Biotechnology | sc-7272 |
| p-STAT-3 | 1:2000 | Santa Cruz Biotechnology | sc-8059 |
| T-STAT-3 | 1:2000 | Santa Cruz Biotechnology | sc-8019 |
| p-STAT-5 | 1:2000 | Santa Cruz Biotechnology | sc-10180b |
| T-STAT-5 | 1:2000 | Santa Cruz Biotechnology | sc-835 |
| FAS | 1:2000 | BD Bioscience | #9452 |
| Perilipin A | 1:2000 | BioVision | #3948-200 |
| p-ATF-2 | 1:2000 | Cell Signaling | #9225 |
| T-ATF-2 | 1:2000 | Cell Signaling | #9226 |
| p-p38 MAPK | 1:2000 | Cell Signaling | #9211 |
| T-p38 MAPK | 1:2000 | Cell Signaling | #9212 |
| β-actin | 1:10,000 | Sigma | A5441 |
| Gene | Forward | Reverse |
|---|---|---|
| FAS | TTGCTGGCACTACAGAATGC | AACAGCCTCAGAGCGACAAT |
| Perilipin A | CTTTCTCGACACACCATGGAAAC | CCACGTTATCCGTAACACCCTTCA |
| 18S rRNA | GGTGAAGGTCGGTGTGAACG | GGTAGGAACACGGAAGGCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perumal, N.L.; Mufida, A.; Yadav, A.K.; Son, D.-G.; Ryoo, Y.-W.; Kim, S.-A.; Jang, B.-C. Suppression of Lipid Accumulation in the Differentiation of 3T3-L1 Preadipocytes and Human Adipose Stem Cells into Adipocytes by TAK-715, a Specific Inhibitor of p38 MAPK. Life 2023, 13, 412. https://doi.org/10.3390/life13020412
Perumal NL, Mufida A, Yadav AK, Son D-G, Ryoo Y-W, Kim S-A, Jang B-C. Suppression of Lipid Accumulation in the Differentiation of 3T3-L1 Preadipocytes and Human Adipose Stem Cells into Adipocytes by TAK-715, a Specific Inhibitor of p38 MAPK. Life. 2023; 13(2):412. https://doi.org/10.3390/life13020412
Chicago/Turabian StylePerumal, Nivethasri Lakshmana, Amila Mufida, Anil Kumar Yadav, Dae-Gu Son, Young-Wook Ryoo, Sung-Ae Kim, and Byeong-Churl Jang. 2023. "Suppression of Lipid Accumulation in the Differentiation of 3T3-L1 Preadipocytes and Human Adipose Stem Cells into Adipocytes by TAK-715, a Specific Inhibitor of p38 MAPK" Life 13, no. 2: 412. https://doi.org/10.3390/life13020412
APA StylePerumal, N. L., Mufida, A., Yadav, A. K., Son, D.-G., Ryoo, Y.-W., Kim, S.-A., & Jang, B.-C. (2023). Suppression of Lipid Accumulation in the Differentiation of 3T3-L1 Preadipocytes and Human Adipose Stem Cells into Adipocytes by TAK-715, a Specific Inhibitor of p38 MAPK. Life, 13(2), 412. https://doi.org/10.3390/life13020412