
Agaricus bisporus-Derived Glucosamine Hydrochloride Regulates VEGF through BMP Signaling to Promote Zebrafish Vascular Development and Impairment Repair

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. GAH and Inhibitors
2.2. Transgenic Lines
2.3. Drug Treatment
2.4. BrdU Staining and WISH
2.5. Quantitative PCR and ELISA
2.6. Statistical Analyses
3. Results
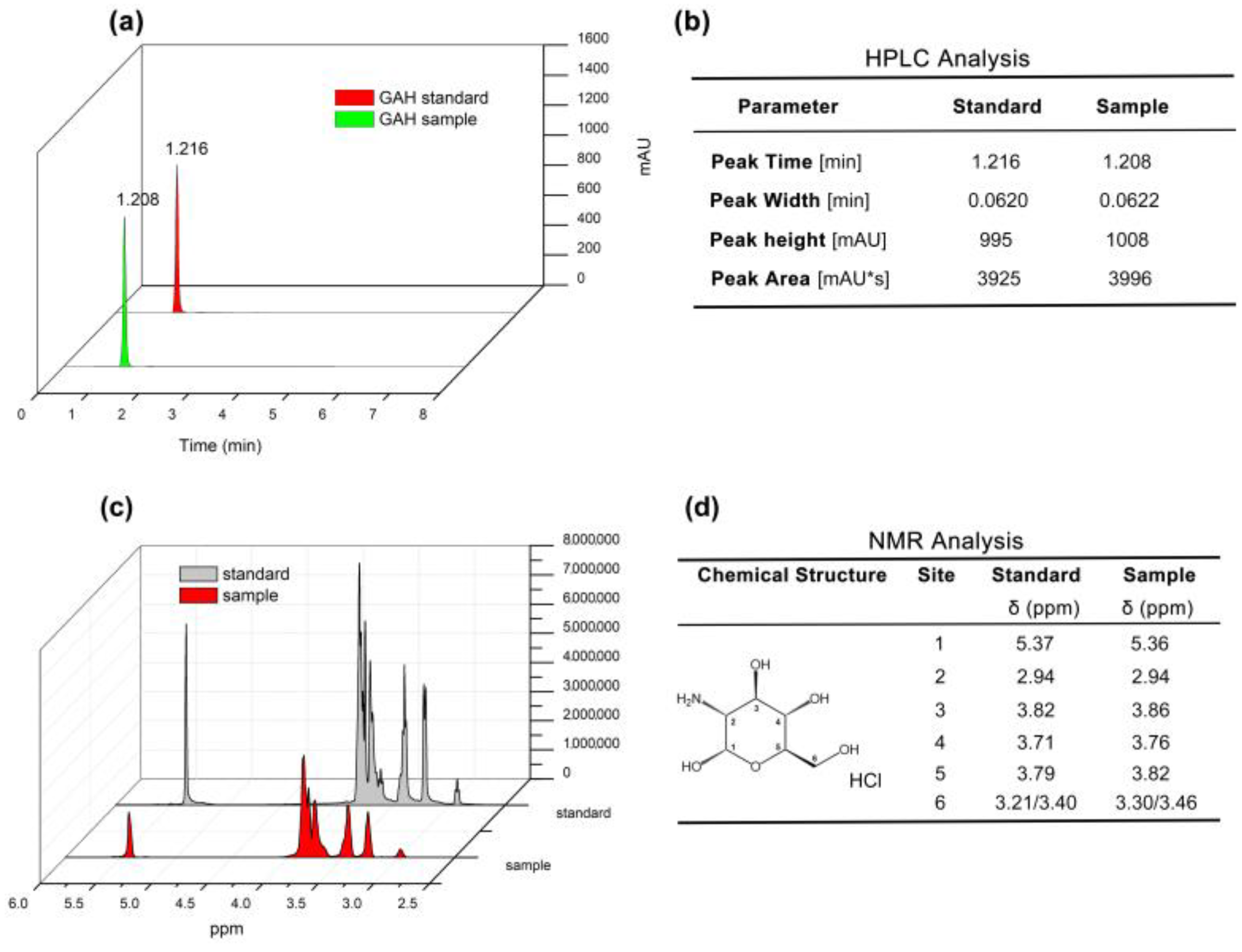
3.1. HPLC and 1HNMR Analysis of Agaricus bisporus GAH
3.2. GAH Promotes Vascular Development in Zebrafish
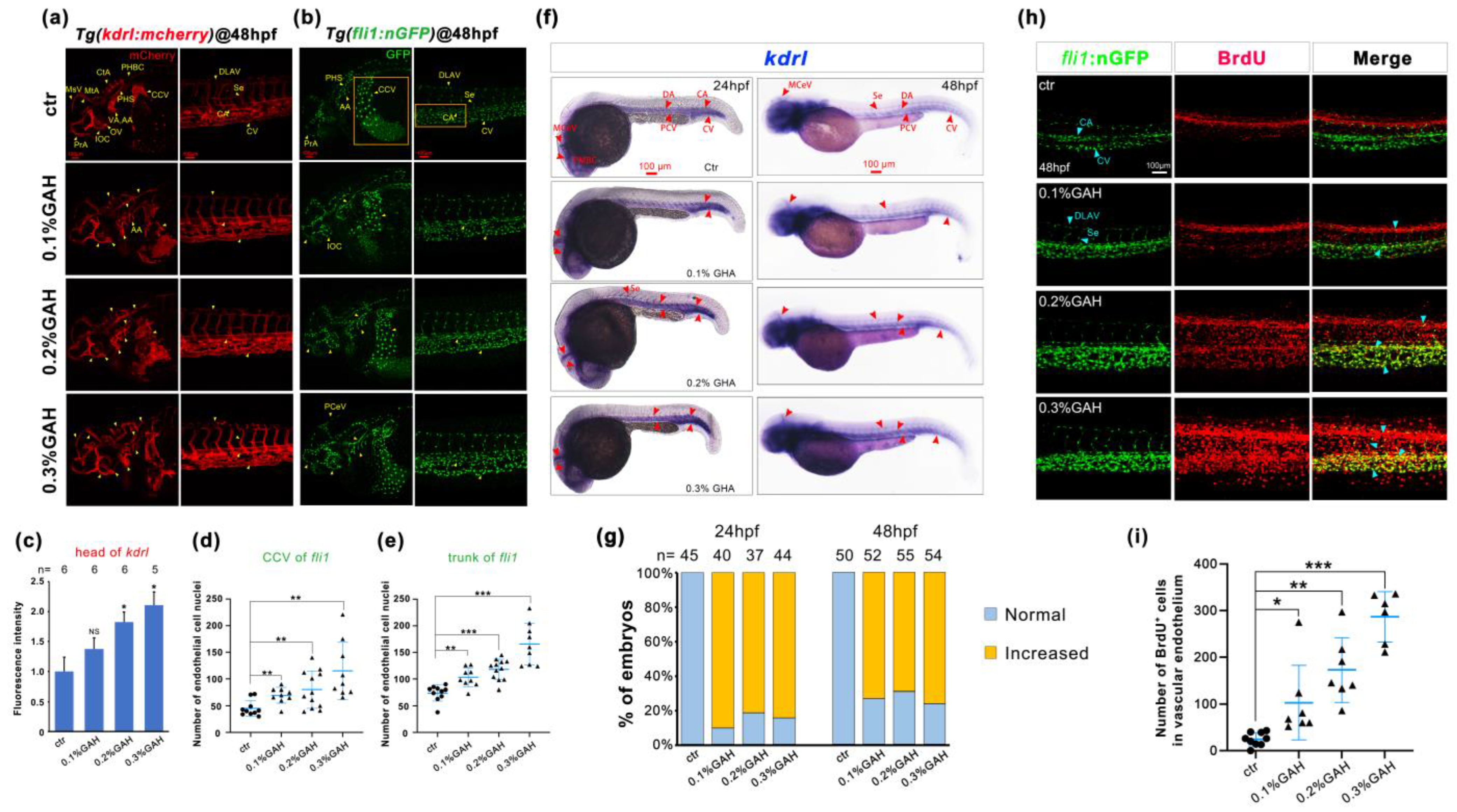
3.3. GAH Promotes Vascular Development through VEGF Signaling
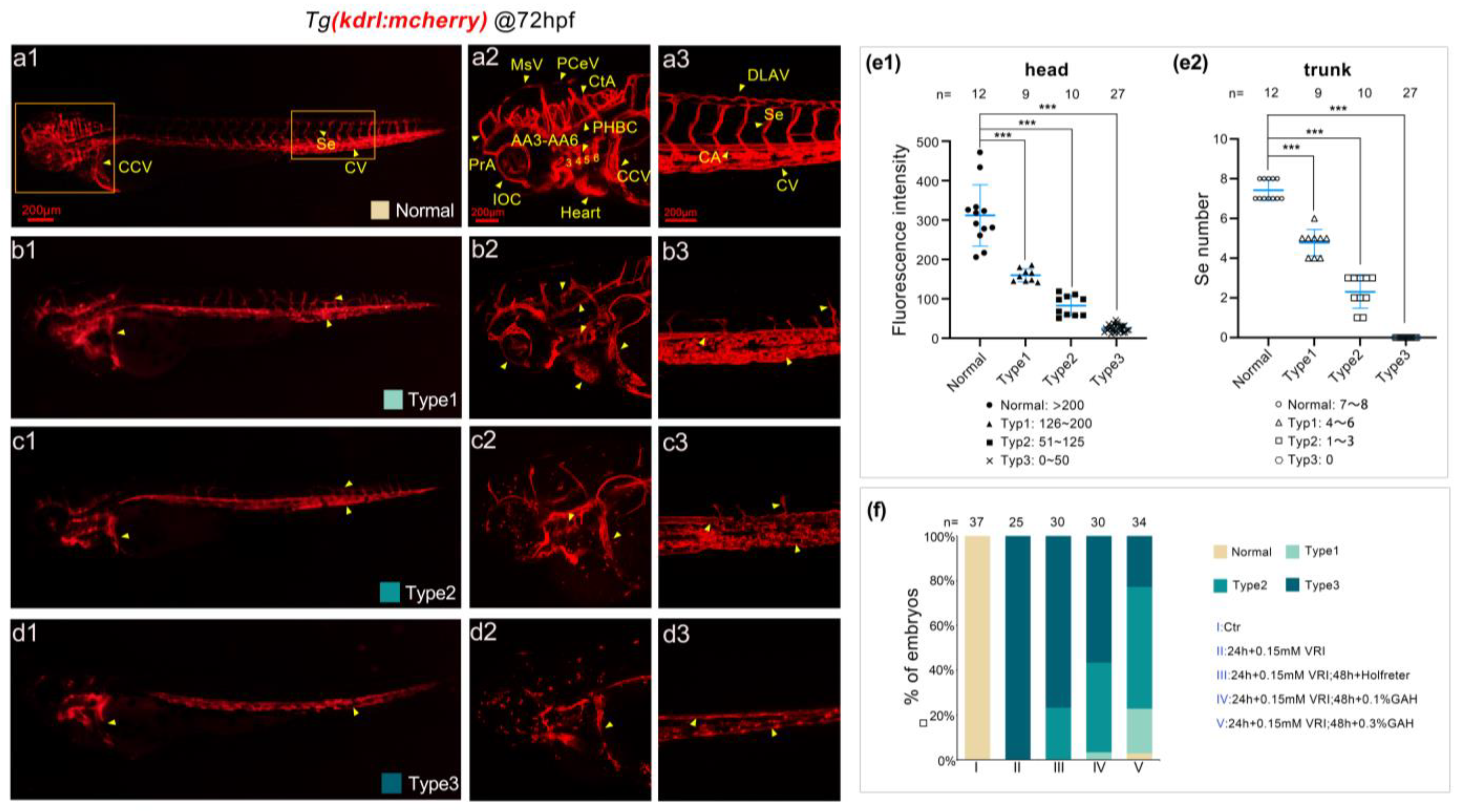
3.4. GAH Repairs Vascular Impairment Caused by DMH1 and DM
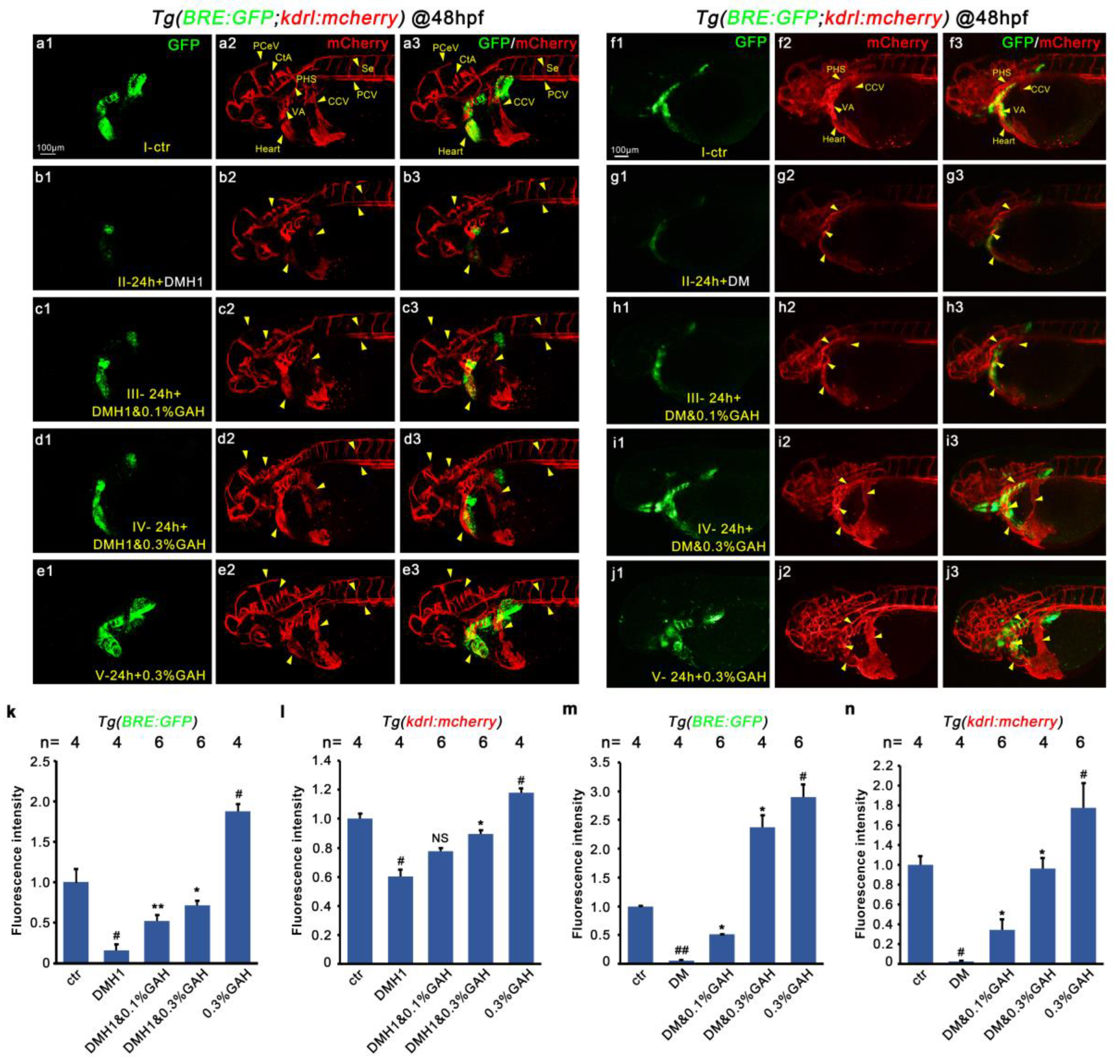
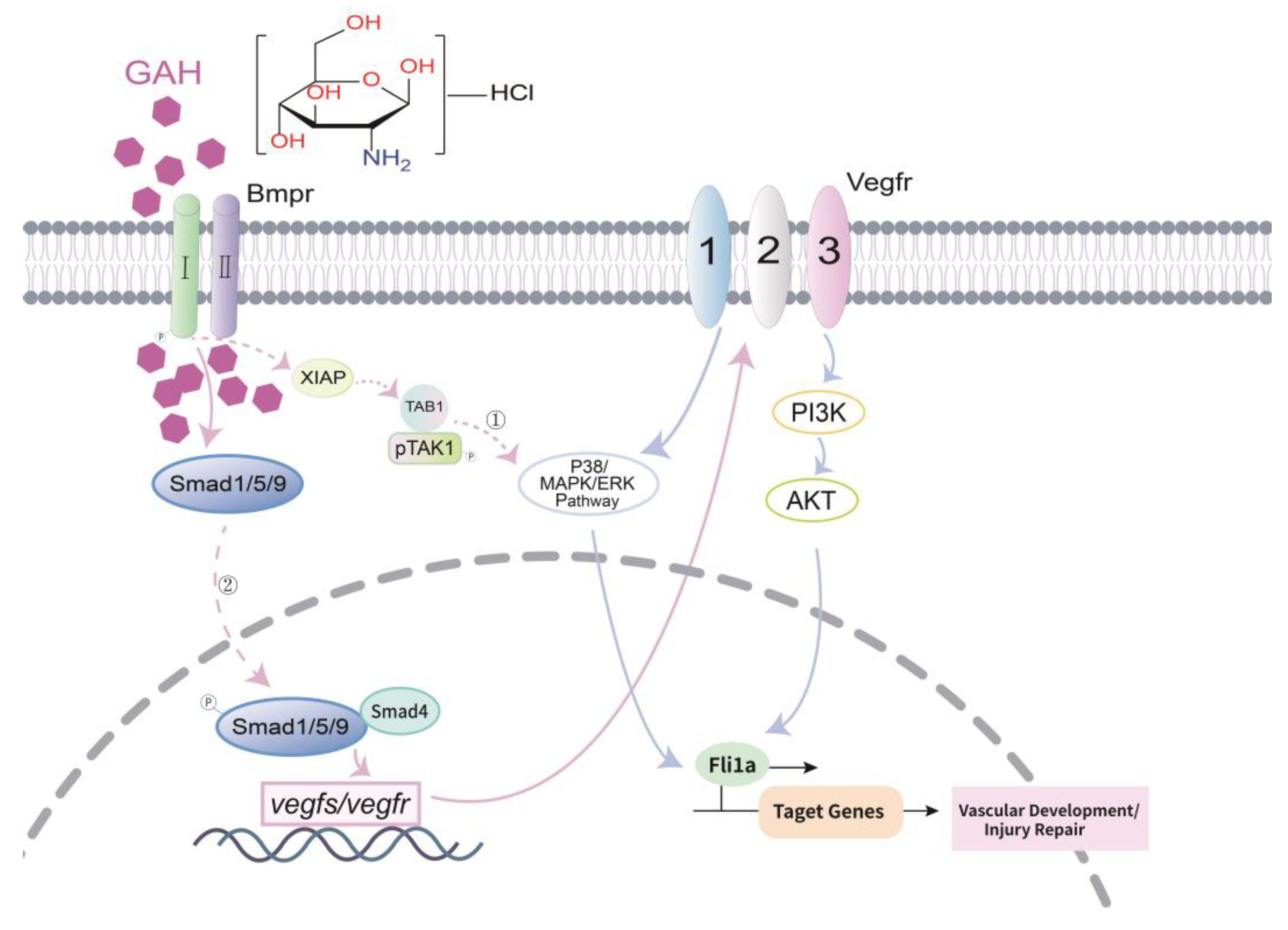
3.5. GAH Regulates VEGF to Repair Vascular Impairment through BMP Signaling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dahmer, S.; Schiller, R.M. Glucosamine. Am. Fam. Physician 2008, 78, 471–476. [Google Scholar]
- Zhang, J.; Ge, R.; Yang, Z. Effect of celecoxib combined with glucosamine hydrochloride in promoting the functional recovery and decreasing the inflammatory factor levels in patients with knee osteoarthritis. Pak. J. Pharm. Sci. 2021, 34, 1277–1282. [Google Scholar]
- Bascoul-Colombo, C.; Garaiova, I.; Plummer, S.F.; Harwood, J.L.; Caterson, B.; Hughes, C.E. Glucosamine Hydrochloride but Not Chondroitin Sulfate Prevents Cartilage Degradation and Inflammation Induced by Interleukin-1α in Bovine Cartilage Explants. Cartilage 2016, 7, 70–81. [Google Scholar] [CrossRef]
- Mel’nikova, N.B.; Sidorova, M.V.; Sorokina, A.R.; Martusevich, A.K.; Peretyagin, S.P.; Kochubeinik, A.V.; Gulenova, M.V.; Bol’shakova, A.E. Complexes of Glucosamine Hydrochloride and Phytic Acid in Aqueous Medium and Their Antioxidant Properties in Human Blood Plasma. Pharm. Chem. J. 2016, 49, 667–671. [Google Scholar] [CrossRef]
- Peng, W.; Zhang, W.; Wu, Q.; Cai, S.; Jia, T.; Sun, J.; Lin, Z.; Alitongbieke, G.; Chen, Y.; Su, Y.; et al. Agaricus bisporus-Derived Glucosamine Hydrochloride Facilitates Skeletal Injury Repair through Bmp Signaling in Zebrafish Osteoporosis Model. J. Nat. Prod. 2021, 84, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Bhathal, A.; Spryszak, M.; Louizos, C.; Frankel, G. Glucosamine and chondroitin use in canines for osteoarthritis: A review. Open Vet. J. 2017, 7, 36–49. [Google Scholar] [CrossRef]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef]
- Deckers, M.M.; van Bezooijen, R.L.; van der Horst, G.; Hoogendam, J.; van Der Bent, C.; Papapoulos, S.E.; Lowik, C.W. Bone morphogenetic proteins stimulate angiogenesis through osteoblast-derived vascular endothelial growth factor A. Endocrinology 2002, 143, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Kaluza, D.; Jakobsson, L. VEGF, Notch and TGFbeta/BMPs in regulation of sprouting angiogenesis and vascular patterning. Biochem. Soc. Trans. 2014, 42, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Kimmel, C.B. Genetics and early development of zebrafish. Trends Genet. 1989, 5, 283–288. [Google Scholar] [CrossRef]
- Gore, A.V.; Pillay, L.M.; Venero Galanternik, M.; Weinstein, B.M. The zebrafish: A fintastic model for hematopoietic development and disease. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e312. [Google Scholar] [CrossRef]
- Dooley, K.; Zon, L.I. Zebrafish: A model system for the study of human disease. Curr. Opin. Genet. Dev. 2000, 10, 252–256. [Google Scholar] [CrossRef]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, Y.; Fukuhara, S.; Ando, K.; Matsuda, M.; Mochizuki, N. Cdc42 mediates Bmp-induced sprouting angiogenesis through Fmnl3-driven assembly of endothelial filopodia in zebrafish. Dev. Cell 2015, 32, 109–122. [Google Scholar] [CrossRef]
- Zeng, X.X.; Zheng, X.; Xiang, Y.; Cho, H.P.; Jessen, J.R.; Zhong, T.P.; Solnica-Krezel, L.; Brown, H.A. Phospholipase D1 is required for angiogenesis of intersegmental blood vessels in zebrafish. Dev. Biol. 2009, 328, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Casie Chetty, S.; Rost, M.S.; Enriquez, J.R.; Schumacher, J.A.; Baltrunaite, K.; Rossi, A.; Stainier, D.Y.; Sumanas, S. Vegf signaling promotes vascular endothelial differentiation by modulating etv2 expression. Dev. Biol. 2017, 424, 147–161. [Google Scholar] [CrossRef]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Kamei, M.; Isogai, S.; Weinstein, B.M. Imaging blood vessels in the zebrafish. Methods Cell Biol. 2004, 76, 51–74. [Google Scholar] [CrossRef]
- Hoareau, M.; El Kholti, N.; Debret, R.; Lambert, E. Zebrafish as a Model to Study Vascular Elastic Fibers and Associated Pathologies. Int. J. Mol. Sci. 2022, 23, 2102. [Google Scholar] [CrossRef]
- Gora-Kupilas, K.; Josko, J. The neuroprotective function of vascular endothelial growth factor (VEGF). Folia Neuropathol. 2005, 43, 31–39. [Google Scholar]
- Matsumoto, K.; Ema, M. Roles of VEGF-A signalling in development, regeneration, and tumours. J. Biochem. 2014, 156, 1–10. [Google Scholar] [CrossRef]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.P.; Liu, K.L.; Li, X.Y.; Lu, G.Q.; Xue, W.H.; Qian, X.H.; Mohamed, O.K.; Meng, F.H. Design, synthesis, and in vitro and in vivo anti-angiogenesis study of a novel vascular endothelial growth factor receptor-2 (VEGFR-2) inhibitor based on 1,2,3-triazole scaffold. Eur. J. Med. Chem. 2021, 211, 113083. [Google Scholar] [CrossRef]
- Nickel, J.; Mueller, T.D. Specification of BMP Signaling. Cells 2019, 8, 1579. [Google Scholar] [CrossRef]
- Lowery, J.W.; Rosen, V. The BMP Pathway and Its Inhibitors in the Skeleton. Physiol. Rev. 2018, 98, 2431–2452. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Boergermann, J.H.; Kopf, J.; Yu, P.B.; Knaus, P. Dorsomorphin and LDN-193189 inhibit BMP-mediated Smad, p38 and Akt signalling in C2C12 cells. Int. J. Biochem. Cell Biol. 2010, 42, 1802–1807. [Google Scholar] [CrossRef]
- Li, L.; Ning, G.; Yang, S.; Yan, Y.; Cao, Y.; Wang, Q. BMP signaling is required for nkx2.3-positive pharyngeal pouch progenitor specification in zebrafish. PLoS Genet. 2019, 15, e1007996. [Google Scholar] [CrossRef]
- de Pater, E.; Ciampricotti, M.; Priller, F.; Veerkamp, J.; Strate, I.; Smith, K.; Lagendijk, A.K.; Schilling, T.F.; Herzog, W.; Abdelilah-Seyfried, S.; et al. Bmp signaling exerts opposite effects on cardiac differentiation. Circ. Res. 2012, 110, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Plaster, N.; Sonntag, C.; Busse, C.E.; Hammerschmidt, M. p53 deficiency rescues apoptosis and differentiation of multiple cell types in zebrafish flathead mutants deficient for zygotic DNA polymerase delta1. Cell Death Differ. 2006, 13, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zheng, X.; Huang, L.; Xu, P.; Ma, Y.; Min, Z.; Tao, Q.; Tao, Y.; Meng, A. Organizer-derived Bmp2 is required for the formation of a correct Bmp activity gradient during embryonic development. Nat. Commun. 2014, 5, 3766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, J.; Qiu, J.; Xing, C.; Li, X.; Leng, B.; Su, Y.; Lin, J.; Lin, J.; Mei, X.; et al. Novel and rapid osteoporosis model established in zebrafish using high iron stress. Biochem. Biophys. Res. Commun. 2018, 496, 654–660. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Chen, X. Transcription regulation of the vegf gene by the BMP/Smad pathway in the angioblast of zebrafish embryos. Biochem. Biophys. Res. Commun. 2005, 329, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Liao, L.; Tian, W. Stem Cell-Based Dental Pulp Regeneration: Insights from Signaling Pathways. Stem Cell Rev. Rep. 2021, 17, 1251–1263. [Google Scholar] [CrossRef]
- Zhang, Y.; Que, J. BMP Signaling in Development, Stem Cells, and Diseases of the Gastrointestinal Tract. Annu. Rev. Physiol. 2020, 82, 251–273. [Google Scholar] [CrossRef]
- Bai, Y.; Leng, Y.; Yin, G.; Pu, X.; Huang, Z.; Liao, X.; Chen, X.; Yao, Y. Effects of combinations of BMP-2 with FGF-2 and/or VEGF on HUVECs angiogenesis in vitro and CAM angiogenesis in vivo. Cell Tissue Res. 2014, 356, 109–121. [Google Scholar] [CrossRef]
- Li, Y.; Sun, R.; Zou, J.; Ying, Y.; Luo, Z. Dual Roles of the AMP-Activated Protein Kinase Pathway in Angiogenesis. Cells 2019, 8, 752. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Wu, Q.; Wei, Y.; Zhao, W.; Lv, H.; Peng, W.; Zheng, J.; Chen, Y.; Wang, Z.; Pan, Y.; et al. Agaricus bisporus-Derived Glucosamine Hydrochloride Regulates VEGF through BMP Signaling to Promote Zebrafish Vascular Development and Impairment Repair. Life 2023, 13, 2330. https://doi.org/10.3390/life13122330
Sun J, Wu Q, Wei Y, Zhao W, Lv H, Peng W, Zheng J, Chen Y, Wang Z, Pan Y, et al. Agaricus bisporus-Derived Glucosamine Hydrochloride Regulates VEGF through BMP Signaling to Promote Zebrafish Vascular Development and Impairment Repair. Life. 2023; 13(12):2330. https://doi.org/10.3390/life13122330
Chicago/Turabian StyleSun, Jiarui, Qici Wu, Yuxin Wei, Wei Zhao, Haokun Lv, Wei Peng, Jiayi Zheng, Yixuan Chen, Zhengsen Wang, Yutian Pan, and et al. 2023. "Agaricus bisporus-Derived Glucosamine Hydrochloride Regulates VEGF through BMP Signaling to Promote Zebrafish Vascular Development and Impairment Repair" Life 13, no. 12: 2330. https://doi.org/10.3390/life13122330
APA StyleSun, J., Wu, Q., Wei, Y., Zhao, W., Lv, H., Peng, W., Zheng, J., Chen, Y., Wang, Z., Pan, Y., & Xue, Y. (2023). Agaricus bisporus-Derived Glucosamine Hydrochloride Regulates VEGF through BMP Signaling to Promote Zebrafish Vascular Development and Impairment Repair. Life, 13(12), 2330. https://doi.org/10.3390/life13122330