
Ivermectin: A Controversial Focal Point during the COVID-19 Pandemic

 , , , , , , , , and
, , , , , , , , and
Abstract
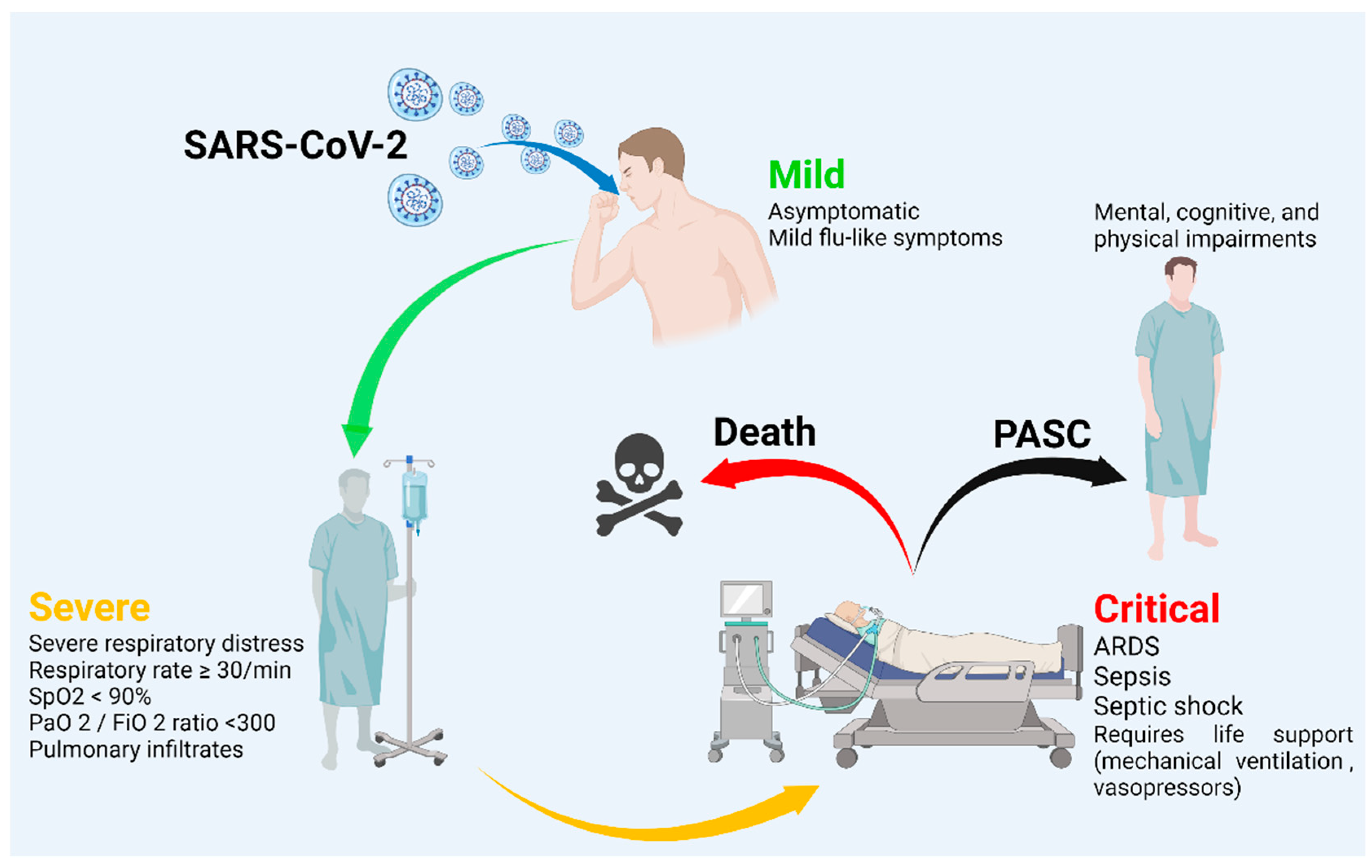
1. Introduction
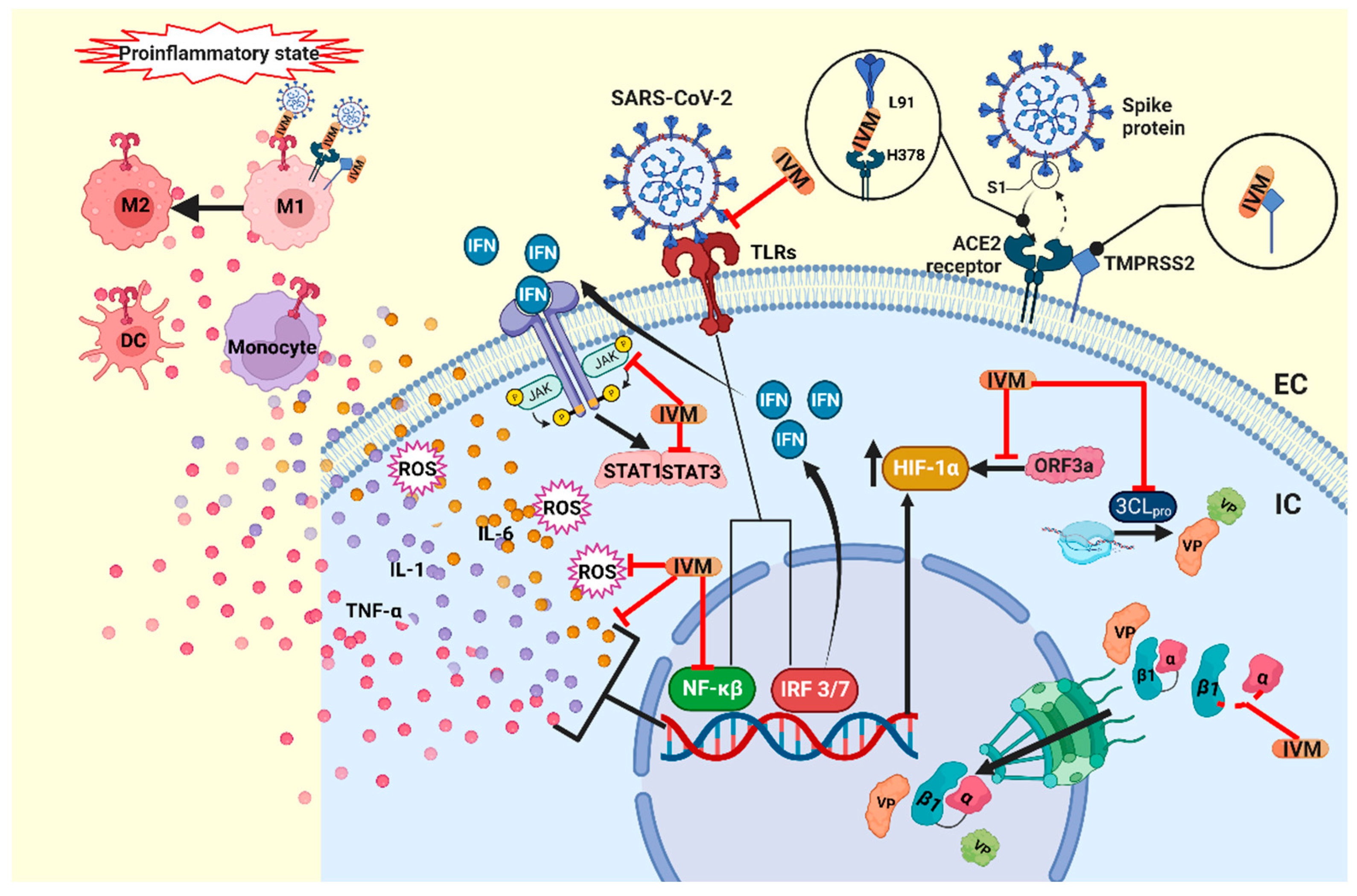
2. Mechanisms of Action of Ivermectin
3. Possible Benefits of Ivermectin in SARS-CoV-2
3.1. Antiviral Activity
3.2. Immunomodulatory Effects
3.3. Antitumoral Mechanisms
4. Systematic Review of Ivermectin in COVID-19
5. Ivermectin in COVID-19 Comorbidities
5.1. Nosocomial Pneumonia
5.2. Wound Healing
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lindley, D. Merck’s new drug free to WHO for river blindness programme. Nature 1987, 329, 752. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Canga, A.G.; Prieto, A.M.S.; Diez Liébana, M.J.; Martínez, N.F.; Sierra Vega, M.; García Vieitez, J.J. The pharmacokinetics and interac-tions of ivermectin in humans—A mini-review. AAPS J. 2008, 10, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Alqahtani, A.; Ilesanmi, O.; Saati, A.; El-Mleeh, A.; Hetta, H.; Beshbishy, A.M. Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects. Pharmaceuticals 2020, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization [WHO]. World Health Organization Model List of Essential Medicines. Ment Holist Health Some Int. Perspect. 2019, 21, 23–24. [Google Scholar]
- Yan, S.; Ci, X.; Chen, N.; Chen, C.; Li, X.; Chu, X.; Li, J.; Deng, X. Anti-inflammatory effects of ivermectin in mouse model of allergic asthma. Agents Actions 2011, 60, 589–596. [Google Scholar] [CrossRef]
- Kircik, L.H.; Del Rosso, J.Q.; Layton, A.M.; Schauber, J. Over 25 Years of Clinical Experience with Ivermectin: An Overview of Safety for an Increasing Number of Indications. J. Drugs Dermatol. 2016, 15, 325–332. [Google Scholar]
- Taylor, D.; Lindsay, A.C.; Halcox, J.P. Correspondance Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2010, 382, 1564–1567. [Google Scholar]
- Montaño, L.M.; Sommer, B.; Gomez-Verjan, J.C.; Morales-Paoli, G.S.; Ramírez-Salinas, G.L.; Solís-Chagoyán, H.; Sanchez-Florentino, Z.A.; Calixto, E.; Pérez-Figueroa, G.E.; Carter, R.; et al. Theophylline: Old Drug in a New Light, Application in COVID-19 through Computational Studies. Int. J. Mol. Sci. 2022, 23, 4167. [Google Scholar] [CrossRef]
- Gorkhali, R.; Koirala, P.; Rijal, S.; Mainali, A.; Baral, A.; Bhattarai, H.K. Structure and Function of Major SARS-CoV-2 and SARS-CoV Proteins. Bioinform. Biol. Insights 2021, 15, 11779322211025876. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Hikmet, F.; Méar, L.; Edvinsson, Å.; Micke, P.; Uhlén, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Xu, H.; Zhong, L.; Deng, J.; Peng, J.; Dan, H.; Zeng, X.; Li, T.; Chen, Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int. J. Oral Sci. 2020, 12, 8. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Gonzalez-Ramirez, J.A.; Ramírez-Nava, J.C.; Gonzalez-Lopez, S.; Sommer, B.; Solís-Chagoyán, H.; Montaño, L.M.; Romero-Martínez, B.S.; Flores-Soto, E. Hyperbaric oxygen therapy in overweight and obese patients with COVID-19. World Acad. Sci. J. 2021, 3, 1–7. [Google Scholar] [CrossRef]
- Jin, J.M.; Bai, P.; He, W.; Wu, F.; Liu, X.F.; Han, D.M.; Liu, S.; Yang, J.K. Gender Differences in Patients with COVID-19: Focus on Severity and Mortality. Front. Public Health 2020, 8, 152. [Google Scholar] [CrossRef]
- Brodin, P. Why is COVID-19 so mild in children? Acta Paediatr. Int. J. Paediatr. 2020, 109, 1082–1083. [Google Scholar] [CrossRef]
- Montaño, L.M.; Sommer, B.; Solís-Chagoyán, H.; Romero-Martínez, B.S.; Aquino-Gálvez, A.; Gomez-Verjan, J.C.; Calixto, E.; González-Avila, G.; Flores-Soto, E. Could Lower Testosterone in Older Men Explain Higher COVID-19 Morbidity and Mortalities? Int. J. Mol. Sci. 2022, 23, 935. [Google Scholar] [CrossRef]
- Klein, S.L.; Morgan, R. The impact of sex and gender on immunotherapy outcomes. Biol. Sex Differ. 2020, 11, 24. [Google Scholar] [CrossRef]
- Chen, T.; Wu, D.; Chen, H.; Yan, W.; Yang, D.; Chen, G.; Ma, K.; Xu, D.; Yu, H.; Wang, H.; et al. Clinical characteristics of 113 deceased patients with coronavirus disease 2019: Retrospective study. BMJ 2020, 368, m1091. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.P. Risk factors for COVID-19 in India. Monaldi Arch. Chest Dis. 2022, 92. [Google Scholar] [CrossRef] [PubMed]
- Santorelli, G.; McCooe, M.; Sheldon, T.A.; Wright, J.; Lawton, T. Ethnicity, pre-existing comorbidities, and outcomes of hospital-ised patients with COVID-19. Wellcome Open Res. 2021, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Mathur, R.; Rentsch, C.T.; Morton, C.E.; Hulme, W.J.; Schultze, A.; MacKenna, B.; Eggo, R.M.; Bhaskaran, K.; Wong, A.Y.; Williamson, E.J.; et al. Ethnic differences in SARS-CoV-2 infection and COVID-19-related hospitalisation, intensive care unit admission, and death in 17 million adults in England: An observational cohort study using the OpenSAFELY platform. Lancet 2021, 397, 1711–1724. [Google Scholar] [CrossRef]
- Wang, L.; Lv, Q.; Zhang, X.; Jiang, B.; Liu, E.; Xiao, C.; Yu, X.; Yang, C.; Chen, L. The utility of MEWS for predicting the mortality in the elderly adults with COVID-19: A retrospective cohort study with comparison to other predictive clinical scores. PeerJ 2020, 8, e10018. [Google Scholar] [CrossRef]
- Jin, M.; Lu, Z.; Zhang, X.; Wang, Y.; Wang, J.; Cai, Y.; Tian, K.; Xiong, Z.; Zhong, Q.; Ran, X.; et al. Clinical characteristics and risk factors of fatal patients with COVID-19: A retrospective cohort study in Wuhan, China. BMC Infect. Dis. 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Schwab, K.; Schwitzer, E.; Qadir, N. Postacute Sequelae of COVID-19 Critical Illness. Crit. Care Clin. 2022, 38, 455–472. [Google Scholar] [CrossRef]
- Jin, Y.; Yang, H.; Ji, W.; Wu, W.; Chen, S.; Zhang, W.; Duan, G. Virology, epidemiology, pathogenesis, and control of COVID-19. Viruses 2020, 12, 372. [Google Scholar] [CrossRef]
- Parasher, A. COVID-19: Current understanding of its Pathophysiology, Clinical presentation and Treatment. Postgrad. Med, J. 2020, 97, 312–320. [Google Scholar] [CrossRef]
- Rochwerg, B.; Siemieniuk, R.A.; Agoritsas, T.; Lamontagne, F.; Askie, L.; Lytvyn, L.; Agarwal, A.; Leo, Y.-S.; Macdonald, H.; Zeng, L.; et al. A living WHO guideline on drugs for COVID-19. BMJ 2020, 370, m3379. [Google Scholar] [CrossRef]
- Romero-Martínez, B.S.; Montaño, L.M.; Solís-Chagoyán, H.; Sommer, B.; Ramírez-Salinas, G.L.; Pérez-Figueroa, G.E.; Flores-Soto1, E. Possible beneficial actions of caffeine in SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 5460. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Das, N.C.; Patra, R.; Mukherjee, S. In silico analyses on the comparative sensing of SARS-CoV-2 mRNA by the intracellular TLRs of humans. J. Med. Virol. 2021, 93, 2476–2486. [Google Scholar] [CrossRef] [PubMed]
- Patra, R.; Das, N.C.; Mukherjee, S. Targeting human TLRs to combat COVID-19: A solution? J. Med Virol. 2020, 93, 615–617. [Google Scholar] [CrossRef]
- Choudhury, A.; Mukherjee, S. In silico studies on the comparative characterization of the interactions of SARS-CoV-2 spike glycoprotein with ACE-2 receptor homologs and human TLRs. J. Med. Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S. Toll-like receptor 4 in COVID-19: Friend or foe? Future Virol. 2022, 17, 415–417. [Google Scholar] [CrossRef]
- Low, Z.Y.; Yip, A.J.W.; Lal, S.K. Repositioning Ivermectin for COVID-19 treatment: Molecular mechanisms of action against SARS-CoV-2 replication. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166294. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Orozco, S.; Roldan-Gomez, M.C.; Ramirez-Nava, J.C.; Rodriguez-Ayala, V.; Sommer, B.; Solís-Chagoyán, H.; Calixto, E.; Montaño, L.M.; Romero-Martínez, B.S.; Flores-Soto, E. Hyperbaric Oxygen Therapy Efficacy as an Adjuvant for the Systemic Inflammation Reduction in Patients with SARS-CoV-2 Infection. J. Clin. Respir. 2022, 4, 1. [Google Scholar] [CrossRef]
- Machado-Curbelo, C.; González-Quevedo, A. Hypoxemia and cytokine storm in COVID-19: Clinical implications. Med. Rev. 2021, 23, 54–59. [Google Scholar]
- Ginestra, J.C.; Mitchell, O.J.; Anesi, G.L.; Christie, J.D. COVID-19 Critical Illness: A Data-Driven Review. Annu. Rev. Med. 2022, 73, 95–111. [Google Scholar] [CrossRef]
- Pfortmueller, C.A.; Spinetti, T.; Urman, R.D.; Luedi, M.M.; Schefold, J.C. COVID-19-associated acute respiratory distress syndrome (CARDS): Current knowledge on pathophysiology and ICU treatment—A narrative review. Best Pract. Res. Clin. Anaesthesiol. 2020, 35, 351–368. [Google Scholar] [CrossRef]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Qu, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. China medical treatment expert group for COVID-19 2020. Clinical Characteristics of coronavirus disease in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- IVERMECTIN. Micromedex® (Electronic Version); IBM Watson Health: Greenwood Village, CO, USA; Available online: https://www.micromedexsolutions.com/ (accessed on 14 August 2022).
- González, P.; González, F.A.; Ueno, K. Ivermectin in human medicine, an overview of the current status of its clinical applications. Curr. Pharm. Biotechnol. 2012, 13, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Ashour, D.S. Ivermectin: From theory to clinical application. Int. J. Antimicrob. Agents 2019, 54, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antivir. Res. 2020, 178, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Heidary, F.; Gharebaghi, R. Ivermectin: A systematic review from antiviral effects to COVID-19 complementary regimen. J. Antibiot. 2020, 73, 593–602. [Google Scholar] [CrossRef]
- Li, N.; Zhao, L.; Zhan, X. Quantitative proteomics reveals a broad-spectrum antiviral property of ivermectin, benefiting for COVID-19 treatment. J. Cell Physiol. 2021, 236, 2959–2975. [Google Scholar] [CrossRef]
- Wagstaff, K.; Sivakumaran, H.; Heaton, S.; Harrich, D.; Jans, D. Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Rawlinson, S.M.; Hearps, A.C.; Jans, D.A. An AlphaScreen®-based assay for high-throughput screening for spe-cific inhibitors of nuclear import. J. Biomol. Screen. 2011, 16, 192–200. [Google Scholar] [CrossRef]
- Yang, S.N.Y.; Atkinson, S.C.; Wang, C.; Lee, A.; Bogoyevitch, M.A.; Borg, N.A.; Jans, D.A. The broad spectrum antiviral ivermectin targets the host nuclear transport importin α/β1 heterodimer. Antivir. Res. 2020, 177, 104760. [Google Scholar] [CrossRef]
- Lv, C.; Liu, W.; Wang, B.; Dang, R.; Qiu, L.; Ren, J.; Yan, C.; Yang, Z.; Wang, X. Ivermectin inhibits DNA polymerase UL42 of pseudorabies virus entrance into the nucleus and proliferation of the virus in vitro and vivo. Antivir. Res. 2018, 159, 55–62. [Google Scholar] [CrossRef]
- Wolff, G.; Limpens, R.W.; Zevenhoven-Dobbe, J.C.; Laugks, U.; Zheng, S.; de Jong, A.W.; Koning, R.I.; Agard, D.A.; Grünewald, K.; Koster, A.J.; et al. A molecular pore spans the double membrane of the coronavirus replication organelle. Science 2020, 369, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Limpens, R.W.A.L.; de Wilde, A.H.; de Jong, A.W.M.; Zevenhoven-Dobbe, J.C.; Maier, H.J.; Faas, F.F.G.A.; Koster, A.J.; Bárcena, M. A unifying structural and functional model of the coronavirus replication organelle: Tracking down RNA synthesis. PLoS Biol. 2020, 18, e3000715. [Google Scholar] [CrossRef] [PubMed]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar]
- Schmith, V.D.; Zhou, J.; Lohmer, L.R. The Approved Dose of Ivermectin Alone is not the Ideal Dose for the Treatment of COVID-19. Clin. Pharmacol. Ther. 2020, 108, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, S.; Rheinstein, P.H. Ivermectin docks to the SARS-CoV-2 spike receptor-binding domain attached to ACE2. In Vivo 2020, 34, 3023–3026. [Google Scholar] [CrossRef] [PubMed]
- Elalfy, H.; Besheer, T.; El-Mesery, A.; El-Gilany, A.H.; Soliman, M.A.A.; Alhawarey, A.; Alegezy, M.; Elhadidy, T.; Hewidy, A.A.; Zaghloul, H.; et al. Effect of a combination of nitazoxanide, ribavirin, and ivermectin plus zinc supplement (MANS.NRIZ study) on the clearance of mild COVID-19. J. Med. Virol. 2021, 93, 3176–3183. [Google Scholar] [CrossRef]
- Choudhury, A.; Das, N.C.; Patra, R.; Bhattacharya, M.; Ghosh, P.; Patra, B.C.; Mukherjee, S. Exploring the bind-ing efficacy of ivermectin against the key proteins of SARS-CoV-2 pathogenesis: An in silico approach. Future Virol. 2021, 16, 277–291. [Google Scholar] [CrossRef]
- Yuce, M.; Cicek, E.; Inan, T.; Dag, A.B.; Kurkcuoglu, O.; Sungur, F.A. Repurposing of FDA-approved drugs against active site and potential allosteric drug-binding sites of COVID-19 main protease. Proteins Struct. Funct. Bioinform. 2021, 89, 1425–1441. [Google Scholar] [CrossRef]
- Ahmed, S.; Karim, M.M.; Ross, A.G.; Hossain, M.S.; Clemens, J.D.; Sumiya, M.K.; Phru, C.S.; Rahman, M.; Zaman, K.; Somani, J.; et al. A five-day course of ivermectin for the treatment of COVID-19 may reduce the duration of illness. Int. J. Infect. Dis. 2021, 103, 214–216. [Google Scholar] [CrossRef]
- de Melo, G.D.; Lazarini, F.; Larrous, F.; Feige, L.; Kornobis, E.; Levallois, S.; Marchio, A.; Kergoat, L.; Hardy, D.; Cokelaer, T.; et al. Attenuation of clinical and immunological outcomes during SARS-CoV-2 infection by ivermectin. EMBO Mol. Med. 2021, 13, e14122. [Google Scholar] [CrossRef]
- Mahmud, R.; Rahman, M.M.; Alam, I.; Ahmed, K.G.U.; Kabir, A.H.; Sayeed, S.J.B.; Rassel, M.A.; Monayem, F.B.; Islam, M.S.; Islam, M.M.; et al. Ivermectin in combination with doxycycline for treating COVID-19 symptoms: A randomized trial. J. Int. Med. Res. 2021, 49, 03000605211013550. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.L.; Tan, K.S.W.; Chu, J.J.H.; Chow, V.T. Combination Treatment with Remdesivir and Ivermectin Exerts Highly Synergistic and Potent Antiviral Activity Against Murine Coronavirus Infection. Front. Cell. Infect. Microbiol. 2021, 11, 700502. [Google Scholar] [CrossRef]
- Cadegiani, F.A.; Goren, A.; Wambier, C.G.; McCoy, J. Early COVID-19 therapy with azithromycin plus nitazoxanide, ivermectin or hydroxychloroquine in outpatient settings significantly improved COVID-19 outcomes compared to known outcomes in untreated patients. New Microbes New Infect. 2021, 43, 100915. [Google Scholar] [CrossRef]
- Lima-Morales, R.; Méndez-Hernández, P.; Flores, Y.N.; Osorno-Romero, P.; Sancho-Hernández, C.R.; Cuecuecha-Rugerio, E.; Nava-Zamora, A.; Hernández-Galdamez, D.R.; Romo-Dueñas, D.K.; Salmeron, J. Effectiveness of a multidrug therapy consisting of Ivermectin, Azithromycin, Montelukast, and Acetylsalicylic acid to prevent hospitalization and death among ambulatory COVID-19 cases in Tlaxcala, Mexico. Int. J. Infect. Dis. 2021, 105, 598–605. [Google Scholar] [CrossRef]
- Procter, B.C.; Ross, C.; Pickard, V.; Smith, E.; Hanson, C.; McCullough, P.A. Clinical outcomes after early ambulatory multidrug therapy for high-risk SARS-CoV-2 (COVID-19) infection. Rev. Cardiovasc. Med. 2021, 21, 611–614. [Google Scholar]
- Okumuş, N.; Demirtürk, N.; Çetinkaya, R.A.; Güner, R.; Avcı, İ.Y.; Orhan, S.; Konya, P.; Şaylan, B.; Karalezli, A.; Yamanel, L.; et al. Evaluation of the effectiveness and safety of adding ivermectin to treatment in severe COVID-19 patients. BMC Infect. Dis. 2021, 21, 411. [Google Scholar] [CrossRef] [PubMed]
- Laura CMiller, Y.S. Macrophage Polarization in Virus-Host Interactions. J. Clin. Cell Immunol. 2015, 6, 311. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki-Krause, R.; Buisson, B.; Bertrand, S.; Corringer, P.-J.; Galzi, J.-L.; Changeux, J.-P.; Bertrand, D. Ivermectin: A Positive Allosteric Effector of the α7 Neuronal Nicotinic Acetylcholine Receptor. Mol. Pharmacol. 1998, 53, 283–294. [Google Scholar] [CrossRef]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic acetylcholine receptor α7 subunit is an essen-tial regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef]
- Galvis, G.; Lips, K.S.; Kummer, W. Expression of Nicotinic Acetylcholine Receptors on Murine Alveolar Macrophages. J. Mol. Neurosci. 2006, 30, 107–108. [Google Scholar] [CrossRef]
- Rahbar, A.; Shakyba, S.; Ghaderi, M.; Kazemi, K.; Fagheh, A.F.; Farsinejad, P.; Khosravi, A.; Louyeh, P.A.; Mirzaeyian, E.; Chamanara, M.; et al. Ivermectin-functionalized multiwall carbon nanotube enhanced the locomotor activity and neuropathic pain by modulating M1/M2 macrophage and decrease oxidative stress in rat model of spinal cord injury. Heliyon 2021, 7, e07311. [Google Scholar] [CrossRef] [PubMed]
- Ebbelaar, C.C.F.; Venema, A.W.; Van Dijk, M.R. Topical Ivermectin in the Treatment of Papulopustular Rosacea: A Systematic Review of Evidence and Clinical Guideline Recommendations. Dermatol. Ther. 2018, 8, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Li, H.; Yu, Q.; Zhang, X.; Yu, L.; Chen, N.; Song, Y.; Deng, X. Avermectin exerts anti-inflammatory effect by downregulating the nuclear transcription factor kappa-B and mitogen-activated protein kinase activation pathway. Fundam. Clin. Pharmacol. 2009, 23, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Di Nicolantonio, J.J.; Barroso, J.; Mccarty, M. Ivermectin may be a clinically useful anti-inflammatory agent for late-stage COVID-19. Open Heart 2020, 7, e001350. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Chen, C.; Ci, X.; Yu, Q.; Zhang, X.; Deng, X. Protective effect of abamectin on acute lung injury induced by lipopoly-saccharide in mice. Fundam. Clin. Pharmacol. 2011, 25, 700–707. [Google Scholar] [CrossRef]
- Zhang, X.; Song, Y.; Ci, X.; An, N.; Ju, Y.; Li, H.; Wang, X.; Han, C.; Cui, J.; Deng, X. Ivermectin inhibits LPS-induced production of inflammatory cytokines and improves LPS-induced survival in mice. Agents Actions 2008, 57, 524–529. [Google Scholar] [CrossRef]
- Rynkiewicz, E.C.; Clerc, M.; Babayan, S.; Pedersen, A.B. Variation in Local and Systemic Pro-Inflammatory Immune Markers of Wild Wood Mice after Anthelmintic Treatment. Integr. Comp. Biol. 2019, 59, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, T.; Kubli, S.P.; Yoshinaga, S.K.; Pfeffer, K.; Mak, T.W. An aberrant STAT pathway is central to COVID-19. Cell Death Differ. 2020, 27, 3209–3225. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.; Cai, B.; Bai, X.; Zhu, Y. Ivermectin accelerates autophagic death of glioma cells by inhibiting glycolysis through blocking GLUT4 mediated JAK/STAT signaling pathway activation. Environ. Toxicol. 2021, 37, 754–764. [Google Scholar] [CrossRef]
- Lu, H.; Zhou, L.; Zuo, H.; Le, W.; Hu, J.; Zhang, T.; Li, M.; Yuan, Y. Ivermectin synergizes sorafenib in hepatocellular carcinoma via targeting multiple oncogenic pathways. Pharmacol. Res. Perspect. 2022, 10, e00954. [Google Scholar] [CrossRef]
- Zhang, X.; Song, Y.; Xiong, H.; Ci, X.; Li, H.; Yu, L.; Zhang, L.; Deng, X. Inhibitory effects of ivermectin on nitric oxide and prostaglandin E2 pro-duction in LPS-stimulated RAW 264.7 macrophages. Int. Immunopharmacol. 2009, 9, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Siemieniuk, R.A.C.; Bartoszko, J.J.; Ge, L.; Zeraatkar, D.; Izcovich, A.; Kum, E.; Pardo-Hernandez, H.; Rochwerg, B.; Lamontagne, F.; Han, M.A.; et al. Drug treatments for COVID-19: Living systematic review and network meta-analysis. BMJ 2020, 370, m2980. [Google Scholar] [CrossRef] [PubMed]
- Juarez, M.; Schcolnik-Cabrera, A.; Dueñas-Gonzalez, A. The multitargeted drug ivermectin: From an antiparasitic agent to a repositioned cancer drug. Am. J. Cancer Res. 2018, 8, 317–331. [Google Scholar]
- Liu, Y.; Fang, S.; Sun, Q.; Liu, B. Anthelmintic drug ivermectin inhibits angiogenesis, growth and survival of glioblastoma through inducing mitochondrial dysfunction and oxidative stress. Biochem. Biophys. Res. Commun. 2016, 480, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Juarez, M.; Schcolnik-Cabrera, A.; Dominguez-Gomez, G.; Chavez-Blanco, A.; Diaz-Chavez, J.; Duenas-Gonzalez, A. Antitumor effects of ivermectin at clinically feasible concentrations support its clinical development as a repositioned cancer drug. Cancer Chemother. Pharmacol. 2020, 85, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Didier, A.; Loor, F. The abamectin derivative ivermectin is a potent P-glycoprotein inhibitor. Anti-Cancer Drugs 1996, 7, 745–751. [Google Scholar] [CrossRef]
- Yin, J.; Park, G.; Lee, J.E.; Choi, E.Y.; Park, J.Y.; Kim, T.-H.; Park, N.; Jin, X.; Jung, J.-E.; Shin, D.; et al. DEAD-box RNA helicase DDX23 modulates glioma malignancy via elevating miR-21 biogenesis. Brain 2015, 138, 2553–2570. [Google Scholar] [CrossRef]
- DraDraganov, D.; Gopalakrishna-Pillai, S.; Chen, Y.R.; Zuckerman, N.; Moeller, S.; Wang, C.; Ann, D.; Lee, P.P. Modulation of P2X4/P2X7/Pannexin-1 sensitivity to extracellular ATP via Ivermectin induces a non-apoptotic and inflammatory form of cancer cell death. Sci. Rep. 2015, 5, 16222. [Google Scholar] [CrossRef]
- Nörenberg, W.; Sobottka, H.; Hempel, C.; Plötz, T.; Fischer, W.; Schmalzing, G.; Schaefer, M. Positive allosteric modulation by ivermectin of human but not murine P2X7 receptors. J. Cereb. Blood Flow Metab. 2012, 167, 48–66. [Google Scholar] [CrossRef]
- Sharmeen, S.; Skrtic, M.; Sukhai, M.A.; Hurren, R.; Gronda, M.; Wang, X.; Fonseca, S.B.; Sun, H.; Wood, T.E.; Ward, R.; et al. The antiparasitic agent ivermectin induces chlo-ride-dependent membrane hyperpolarization and cell death in leukemia cells. Blood 2010, 116, 3593–3603. [Google Scholar] [CrossRef]
- Dominguez-Gomez, G.; Chavez-Blanco, A.; Medina-Franco, J.L.; Saldivar-Gonzalez, F.; Flores-Torrontegui, Y.; Juarez, M.; Díaz-Chávez, J.; Gonzalez-Fierro, A.; Dueñas-González, A. Ivermectin as an inhibitor of cancer stem-like cells. Mol. Med. Rep. 2017, 17, 3397–3403. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Prichard, R. Ivermectin exhibits potent anti-mitotic activity. Vet. Parasitol. 2016, 226, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Liang, H.; Qu, B.; Li, Y.; Liu, J.; Zhang, Y.; Li, L.; Hu, L.; Zhang, X.; Gao, A. Ivermectin inhibits the growth of glioma cells by inducing cell cycle arrest and apoptosis in vitro and in vivo. J. Cell. Biochem. 2018, 120, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Xu, Q.; Long, J.; Xie, H. Suppressing ROS-TFE3-dependent autophagy enhances ivermectin-induced apoptosis in hu-man melanoma cells. J. Cell Biochem. 2019, 120, 1702–1715. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Lim, W.; Ham, J.; Kim, J.; You, S.; Song, G. Ivermectin induces apoptosis of porcine trophectoderm and uterine luminal epithelial cells through loss of mitochondrial membrane potential, mitochondrial calcium ion overload, and reactive oxygen species generation. Pestic. Biochem. Physiol. 2019, 159, 144–153. [Google Scholar] [CrossRef]
- Kilic, M.; Kasperczyk, H.; Fulda, S.; Debatin, K.-M. Role of hypoxia inducible factor-1 alpha in modulation of apoptosis resistance. Oncogene 2006, 26, 2027–2038. [Google Scholar] [CrossRef]
- Rankin, E.B.; Giaccia, A.J. The role of hypoxia-inducible factors in tumorigenesis. Cell Death Differ. 2008, 15, 678–685. [Google Scholar] [CrossRef]
- Kosyna, F.K.; Nagel, M.; Kluxen, L.; Kraushaar, K.; Depping, R. The importin α/β-specific inhibitor Ivermectin affects HIF-dependent hypoxia response pathways. Biol. Chem. 2015, 396, 1357–1367. [Google Scholar] [CrossRef]
- Vishnoi, K.; Viswakarma, N.; Rana, A.; Rana, B. Transcription Factors in Cancer Development and Therapy. Cancers 2020, 12, 2296. [Google Scholar] [CrossRef]
- Tian, M.; Liu, W.; Li, X.; Zhao, P.; Shereen, M.A.; Zhu, C.; Huang, S.; Liu, S.; Yu, X.; Yue, M.; et al. HIF-1α promotes SARS-CoV-2 infection and aggravates inflammatory responses to COVID-19. Signal Transduct Target Ther. 2021, 6, 308. [Google Scholar] [CrossRef]
- Rakedzon, S.; Neuberger, A.; Domb, A.J.; Petersiel, N.; Schwartz, E. From hydroxychloroquine to ivermectin: What are the anti-viral properties of anti-parasitic drugs to combat SARS-CoV-2? J. Travel Med. 2021, 28, taab005. [Google Scholar] [CrossRef] [PubMed]
- Popp, M.; Stegemann, M.; Metzendorf, M.-I.; Kranke, P.; Meybohm, P.; Skoetz, N.; Weibel, S. Ivermectin for preventing and treating COVID-19. Cochrane Database Syst. Rev. 2021, 7, CD015017. [Google Scholar] [CrossRef] [PubMed]
- Cruciani, M.; Pati, I.; Masiello, F.; Malena, M.; Pupella, S.; De Angelis, V. Ivermectin for prophylaxis and treatment of COVID-19: A systematic review and meta-analysis. Diagnostics 2021, 11, 1645. [Google Scholar] [CrossRef] [PubMed]
- Murchu, E.; Spillane, S.; Byrne, P.; O’Neill, M.; Harrington, P.; Ryan, M. Interventions in an Ambulatory Setting to Prevent Pro-gression to Severe Disease in Patients with COVID-19: A Systematic Review. Ann. Pharmacother. 2022, 56, 309–318. [Google Scholar] [CrossRef]
- Roman, Y.M.; Burela, P.A.; Pasupuleti, V.; Piscoya, A.; E Vidal, J.; Hernandez, A.V. Ivermectin for the Treatment of Coronavirus Disease 2019: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Clin. Infect. Dis. 2021, 74, 1022–1029. [Google Scholar] [CrossRef]
- Deng, J.; Zhou, F.; Ali, S.; Heybati, K.; Hou, W.; Huang, E.; Wong, C.Y. Efficacy and safety of ivermectin for the treatment of COVID-19: A systematic review and meta-analysis. QJM Int. J. Med. 2021, 114, 721–732. [Google Scholar] [CrossRef]
- de Lima Machado, M.L.; Souza, A.T.B.; Linhares, P.V.A.; Ferreira, C.F.M.; Silva, D.F.O.; Martins, R.R.; Cobucci, R. N Effectiveness and safety of ivermectin in the treatment of C OVID-19: Protocol for a systematic review and meta-analysis. BMJ Open 2021, 11, e050532. [Google Scholar] [CrossRef]
- Padhy, B.M.; Mohanty, R.R.; Das, S.; Meher, B.R. Therapeutic potential of ivermectin as add-on treatment in COVID 19: A systematic review and meta-analysis. J. Pharm. Pharm. Sci. 2020, 23, 462–469. [Google Scholar] [CrossRef]
- Chaudhry, M.W.; Zubair, S.M.; Zubairi, A.B.S.; Irfan, M. Role of Ivermectin in Patients Hospitalized with COVID-19: A Systematic Review of Literature. Adv. Respir. Med. 2021, 89, 413–418. [Google Scholar] [CrossRef]
- Bhowmick, S.; Dang, A.; Vallish, B.N.; Dang, S. and Efficacy of Ivermectin and Doxycycline Monotherapy and in Combi-nation in the Treatment of COVID-19: A Scoping Review. Drug Saf. 2021, 44, 635–644. [Google Scholar] [CrossRef]
- Bryant, A.; Lawrie, T.A.; Fordham, E.J. Ivermectin for Prevention and Treatment of COVID-19 Infection: A Systematic Review, Meta-analysis, and Trial Sequential Analysis to Inform Clinical Guidelines. Am. J. Ther. 2021, 28, e434–e460. [Google Scholar] [CrossRef] [PubMed]
- Kudzi, W.; Dodoo, A.N.O.; Mills, J.J. Genetic polymorphisms in MDR1, CYP3A4 and CYP3A5 genes in a Ghanaian population: A plausible explanation for altered metabolism of ivermectin in humans? BMC Med. Genet. 2010, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- Zein, A.F.M.Z.; Sulistiyana, C.S.; Raffaelo, W.M.; Wibowo, A.; Pranata, R. Ivermectin and mortality in patients with COVID-19: A systematic review, meta-analysis, and meta-regression of randomized controlled trials. Diabetes Metab. Syndr. 2021, 15, 102186. [Google Scholar] [CrossRef] [PubMed]
- Elgazzar, A.; Eltaweel, A.; Youssef, S.A.; Hany, B.; Hafez, M.; Moussa, H. Efficacy and safety of ivermectin for treatment and prophylaxis of COVID-19 pandemic. 2020; preprint. [Google Scholar]
- Héctor, C.; Roberto, H.; Psaltis, A.; Veronica, C. Study of the Efficacy and Safety of Topical Ivermectin + Iota-Carrageenan in the Prophylaxis against COVID-19 in Health Personnel. J. Biomed. Res. Clin. Investig. 2020, 2, 1007. [Google Scholar] [CrossRef]
- Marcolino, M.S.; Meira, K.C.; Guimarães, N.S.; Motta, P.P.; Chagas, V.S.; Kelles, S.M.B.; de Sá, L.C.; Valacio, R.A.; Ziegelmann, P.K. Sys-tematic review and meta-analysis of ivermectin for treatment of COVID-19: Evidence beyond the hype. BMC Infect. Dis. 2022, 22, 639. [Google Scholar] [CrossRef]
- Camprubí, D.; Almuedo-Riera, A.; Martí-Soler, H.; Soriano, A.; Hurtado, J.C.; Subirà, C.; Grau-Pujol, B.; Krolewiecki, A.; Muñoz, J. Correction: Lack of efficacy of standard doses of ivermectin in severe COVID-19 patients. PLoS ONE 2022, 17, e0268667. [Google Scholar] [CrossRef]
- López-Medina, E.; López, P.; Hurtado, I.C.; Dávalos, D.M.; Ramirez, O.; Martínez, E.; Díazgranados, J.A.; Oñate, J.M.; Chavarriaga, H.; Herrera, S.; et al. Effect of Ivermectin on Time to Resolution of Symptoms Among Adults with Mild COVID-19: A Randomized Clinical Trial. JAMA 2021, 325, 1426–1435. [Google Scholar] [CrossRef]
- Krolewiecki, A.; Lifschitz, A.; Moragas, M.; Travacio, M.; Valentini, R.; Alonso, D.F.; Solari, R.; Tinelli, M.A.; Cimino, R.O.; Álvarez, L.; et al. Antiviral effect of high-dose ivermectin in adults with COVID-19: A proof-of-concept randomized trial. EClinicalMedicine 2021, 37, 100959. [Google Scholar] [CrossRef]
- Navarro, M.; Camprubí, D.; Requena-Méndez, A.; Buonfrate, D.; Giorli, G.; Kamgno, J.; Gardon, J.; Boussinesq, M.; Muñoz, J.; Krolewiecki, A. Safety of high-dose ivermectin: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2020, 75, 827–834. [Google Scholar] [CrossRef]
- Gil, E.; Martyn, E.; Rokadiya, S.; Jain, S.; Chin, T.L. Bacterial Coinfection in COVID-19. Clin. Infect. Dis. 2021, 73, e843–e845. [Google Scholar] [CrossRef]
- Mahmoudi, H. Bacterial co-infections and antibiotic resistance in patients with COVID-19. GMS Hyg. Infect. Control 2020, 15, 35. [Google Scholar] [CrossRef]
- Rawson, T.M.; Moore, L.S.; Zhu, N.; Ranganathan, N.; Skolimowska, K.; Gilchrist, M.; Satta, G.; Cooke, G.; Holmes, A. Bacterial and fungal co-infection in individuals with coronavirus: A rapid review to support COVID-19 antimicrobial prescribing Timothy. Clin Infect. Dis. 2020, 71, 2459–2468. [Google Scholar] [PubMed]
- Chaudhry, R.; Sreenath, K.; Batra, P.; Vinayaraj, E.V.; Rathor, N.; Saikiran, K.; Aravindan, A.; Singh, V.; Brijwal, M.; Soneja, M.; et al. Atypical bacterial co-infections among patients with COVID-19: A study from India. J. Med. Virol. 2022, 94, 303–309. [Google Scholar] [CrossRef]
- Yang, S.; Hua, M.; Liu, X.; Du, C.; Pu, L.; Xiang, P.; Wang, L.; Liu, J. Bacterial and fungal co-infections among COVID-19 patients in intensive care unit. Microbes Infect. 2021, 23, 104806. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, V.; Lawrence, H.; Lansbury, L.E.; Webb, K.; Safavi, S.; Zainuddin, N.I.; Huq, T.; Eggleston, C.; Ellis, J.; Thakker, C.; et al. Co-infection in critically ill patients with COVID-19: An observational cohort study from England. J. Med. Microbiol. 2021, 70, 001350. [Google Scholar] [CrossRef]
- Gambhir, R.S.; Aggarwal, A.; Bhardwaj, A.; Kaur, A.; Sohi, R.K.; Mehta, S. COVID-19 and mucormycosis (Black Fungus): An epidemic within the pandemic. Rocz. Państwowego Zakładu Hig. 2021, 72, 239–244. [Google Scholar]
- Elabbadi, A.; Turpin, M.; Gerotziafas, G.T.; Teulier, M.; Voiriot, G.; Fartoukh, M. Bacterial coinfection in critically ill COVID-19 pa-tients with severe pneumonia. Infection 2021, 49, 559–562. [Google Scholar] [CrossRef]
- Chong, W.H.; Saha, B.K.; Ramani, A.; Chopra, A. State-of-the-art review of secondary pulmonary infections in patients with COVID-19 pneumonia. Infection 2021, 49, 591–605. [Google Scholar] [CrossRef]
- Mirzaei, R.; Goodarzi, P.; Asadi, M.; Soltani, A.; Aljanabi, H.A.A.; Jeda, A.S.; Dashtbin, S.; Jalalifar, S.; Mohammadzadeh, R.; Teimoori, A.; et al. Bacterial co-infections with SARS-CoV-2. IUBMB Life 2020, 72, 2097–2111. [Google Scholar] [CrossRef] [PubMed]
- Ny, P.; Kelsom, C.; Chron, A.; Lou, M.; Nieberg, P.; Shriner, K.; Huse, H.; Wong-Beringer, A. Factors associated with prompt recovery among hospitalised patients with coronavirus disease 2019. Int. J. Clin. Pract. 2021, 75, e14818. [Google Scholar] [CrossRef]
- Lim, L.E.; Vilchèze, C.; Ng, C.; Jacobs, W.R.; Ramón-García, S.; Thompson, C.J. Anthelmintic avermectins kill mycobacterium tu-berculosis, including multidrug-resistant clinical strains. Antimicrob. Agents Chemother. 2013, 57, 1040–1046. [Google Scholar] [CrossRef]
- Ashraf, S.; Chaudhry, U.; Raza, A.; Ghosh, D.; Zhao, X. In vitro activity of ivermectin against Staphylococcus aureus clinical isolates. Antimicrob. Resist. Infect. Control 2018, 7, 7–12. [Google Scholar] [CrossRef]
- Čulić, O.; Eraković, V.; Parnham, M.J. Anti-inflammatory effects of macrolide antibiotics. Eur. J. Pharmacol. 2001, 429, 209–229. [Google Scholar] [CrossRef]
- Csóka, B.; Németh, Z.H.; Szabó, I.; Davies, D.L.; Varga, Z.V.; Pálóczi, J.; Falzoni, S.; Di Virgilio, F.; Muramatsu, R.; Yamashita, T.; et al. Macrophage P2X4 receptors augment bacterial killing and protect against sepsis. JCI Insight 2018, 3, e99431. [Google Scholar] [CrossRef]
- Yim, J.; Lim, H.H.; Kwon, Y. COVID-19 and pulmonary fibrosis: Therapeutics in clinical trials, repurposing, and potential development. Arch. Pharmacal Res. 2021, 44, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D.; Costabel, U.; Hansell, D.M.; King, T.E.; Lynch, D.A.; Nicholson, A.G.; Ryerson, C.J.; Ryu, J.H.; Selman, M.; Wells, A.U.; et al. An official American Thoracic Socie-ty/European Respiratory Society statement: Update of the international multidisciplinary classification of the idiopathic interstitial pneumonias. Am. J. Respir. Crit. Care Med. 2013, 188, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Sia, D.K.; Mensah, K.B.; Opoku-Agyemang, T.; Folitse, R.D.; Darko, D.O. Mechanisms of ivermectin-induced wound healing. BMC Vet.-Res. 2020, 16, 397. [Google Scholar] [CrossRef] [PubMed]
- Feaster, B.; Cline, A.; Feldman, S.R.; Taylor, S. Clinical effectiveness of novel rosacea therapies. Curr. Opin. Pharmacol. 2019, 46, 14–18. [Google Scholar] [CrossRef]
- Charnowski, S.; Wollenberg, A.; Reinholz, M. Perioral Dermatitis Successfully Treated with Topical Ivermectin. Ann. Dermatol. 2019, 31, S27–S28. [Google Scholar] [CrossRef]
- Ventre, E.; Rozières, A.; Lenief, V.; Albert, F.; Rossio, P.; Laoubi, L.; Dombrowicz, D.; Staels, B.; Ulmann, L.; Julia, V.; et al. Topical ivermectin improves allergic skin inflammation. Allergy 2017, 72, 1212–1221. [Google Scholar] [CrossRef]
- Barańska-Rybak, W.; Kowalska-Olędzka, E. New indications for topical ivermectin 1% cream: A case series study. Adv. Dermatol. Allergol. 2019, 36, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Reis, G.; Silva, E.A.; Silva, D.C.; Thabane, L.; Milagres, A.C.; Ferreira, T.S.; dos Santos, C.V.; Campos, V.H.; Nogueira, A.M.; de Almeida, A.P.; et al. Effect of Early Treatment with Ivermectin among Patients with COVID-19. N. Engl. J. Med. 2022, 386, 1721–1731. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study Population | Combination IVM with: | Results |
|---|---|---|
| Sixty-two patients on a triple combination therapy versus fifty-one patients on symptomatic supportive therapy matched for age and sex. | Nitazoxanide and Ribavirin compared to routine supportive treatment. | This study showed that the clearance rates were 58.1% and 0% on day 7 and 73.1% and 13.7% on day 15 in the combined antiviral group compared to the symptomatic support treatment group. Therefore, the combined use of nitazoxanide, ribavirin and ivermectin plus a zinc supplement effectively eliminated SARS-CoV2 from the nasopharynx in a shorter time than symptomatic therapy [57]. |
| Two hundred patients with mild to moderate symptoms of COVID-19 were randomly assigned to the treatment group and two hundred to the placebo group. | Doxycycline versus placebo. | The median time to recovery was seven days (4–10) in the treatment group and 9 (5–12) in the placebo group, while the percentage of patients with a recovery of ≤7 days was 61% and 44%, respectively [62]. |
| In vitro model of RAW264.7 macrophages infected with MHV. | Remdesivir. | The combination of remdesivir and ivermectin showed a highly potent synergism by significantly reducing the 7-log10 of live virus and 2.5-log10 of viral RNA in infected macrophages. This combination also resulted in the lowest IL-6, TNF-a and leukemia inhibitory factors [63]. |
| The intervention group of five hundred and eighty-five patients and control group of five hundred eighty-five patients were treated with a placebo, along with a second control group of one hundred and thirty-seven untreated patients. | Azithromycin plus nitazoxanide or hydroxychloroquine. | Compared with control group 1 and control group 2, the intervention group showed a 31.5 to 36.5% reduction in viral excretion (p < 0.0001), 70 to 85% in the duration of symptoms (p < 0.0001) and 100% in respiratory complications, hospitalization, mechanical ventilation, deaths and post-COVID manifestations (p < 0.0001). For every 1000 confirmed cases of COVID-19, at least 70 hospitalizations, 50 mechanical ventilation and 5 deaths were averted [64]. |
| Four hundred and eighty-one patients with combined therapy and two hundred and eighty-seven with standard treatment. | Azithromycin, montelukast, and acetylsalicylic acid vs. standard therapy. | A total of 85% of cases who received the combined therapy recovered within 14 days, and the total was 59% in the comparison group. The likelihood of recovery within 14 days was 3.4 times greater among the combined therapy group than in the comparison group. Patients treated with the combined therapy had a 75% and 81% lower risk of being hospitalized and death, respectively, than the comparison group [65]. |
| Nine hundred and twenty-two outpatients, of which three hundred and twenty were given a multidrug therapy with ivermectin. | At least two agents with antiviral activity against SARS-CoV-2 (zinc, hydroxychloroquine) and one antibiotic (azithromycin, doxycycline, ceftriaxone). | A total of 320/922 (34.7%) patients were treated, resulting in 6/320 (1.9%) and 1/320 (0.3%) patients hospitalized and who died, respectively. We concluded that early ambulatory (not hospitalized, treated at home) multidrug therapy is safe, feasible and associated with low rates of hospitalization and death [66]. |
| Sixty-six patients were included in the study, with thirty-six in the study group and thirty in the control group. | Reference treatment protocol: hydroxychloroquine + favipiravir + azithromycin. Patients in the control group received only standard treatment with three other drugs, without ivermectin. | At the end of the first 5-day follow-up period, the rate of clinical improvement was 73.3% (22/30) in the study group and 53.3% (16/30) in the control group (p = 0.10). At the end of the follow-up period, the mean peripheral capillary oxygen saturation (SpO2) values of the study and control groups were 93.5 and 93.0%, respectively. PaO2/FiO2 ratios were determined as 236.3 ± 85.7 and 220.8 ± 127.3 in the study and control groups, respectively. At the end of the follow-up period, mortality was recorded for 6 patients (20%) in the study group and 9 (30%) patients in the control group (p = 0.37) [67]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillejos-López, M.; Torres-Espíndola, L.M.; Huerta-Cruz, J.C.; Flores-Soto, E.; Romero-Martinez, B.S.; Velázquez-Cruz, R.; Higuera-Iglesias, A.; Camarena, Á.; Torres-Soria, A.K.; Salinas-Lara, C.; et al. Ivermectin: A Controversial Focal Point during the COVID-19 Pandemic. Life 2022, 12, 1384. https://doi.org/10.3390/life12091384
Castillejos-López M, Torres-Espíndola LM, Huerta-Cruz JC, Flores-Soto E, Romero-Martinez BS, Velázquez-Cruz R, Higuera-Iglesias A, Camarena Á, Torres-Soria AK, Salinas-Lara C, et al. Ivermectin: A Controversial Focal Point during the COVID-19 Pandemic. Life. 2022; 12(9):1384. https://doi.org/10.3390/life12091384
Chicago/Turabian StyleCastillejos-López, Manuel, Luz Maria Torres-Espíndola, Juan Carlos Huerta-Cruz, Edgar Flores-Soto, Bianca S. Romero-Martinez, Rafael Velázquez-Cruz, Anjarath Higuera-Iglesias, Ángel Camarena, Ana Karen Torres-Soria, Citlaltepetl Salinas-Lara, and et al. 2022. "Ivermectin: A Controversial Focal Point during the COVID-19 Pandemic" Life 12, no. 9: 1384. https://doi.org/10.3390/life12091384
APA StyleCastillejos-López, M., Torres-Espíndola, L. M., Huerta-Cruz, J. C., Flores-Soto, E., Romero-Martinez, B. S., Velázquez-Cruz, R., Higuera-Iglesias, A., Camarena, Á., Torres-Soria, A. K., Salinas-Lara, C., Fernández-Plata, R., Alvarado-Vásquez, N., Solís-Chagoyán, H., Ruiz, V., & Aquino-Gálvez, A. (2022). Ivermectin: A Controversial Focal Point during the COVID-19 Pandemic. Life, 12(9), 1384. https://doi.org/10.3390/life12091384