


Upregulation of Spinal miR-155-5p Contributes to Mechanical Hyperalgesia by Promoting Inflammatory Activation of Microglia in Bone Cancer Pain Rats

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bone Cancer Pain Model
2.2. Behavioral Assessment
2.3. Tissue Preparation
2.4. Sequencing and Analysis of miRNAs
2.5. N9 Cell Culture and Treatment
2.6. Real-Time Quantitative PCR (RT-qPCR)
2.7. Western Blot Analysis
2.8. Fluorescence In Situ Hybridization (FISH) and Immunofluorescence Assay
2.9. Dual-Luciferase Reporter Assay
2.10. Lentivirus Infection In Vitro and In Vivo
2.11. Statistical Analysis
3. Results
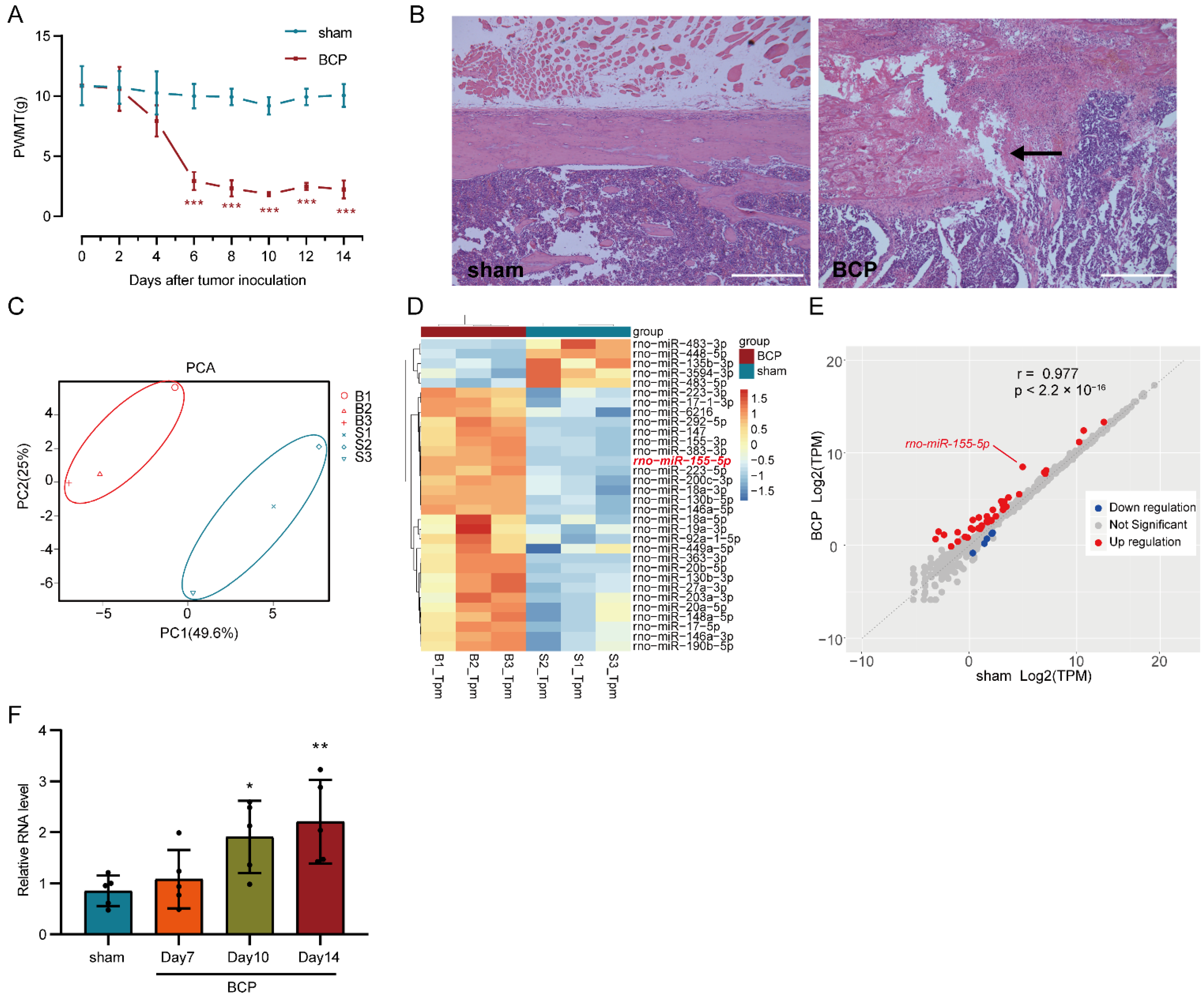
3.1. Mechanical Hyperalgesia Developed in BCP Rats and Differentially Expressed miRNAs Were Filtered Out Using RNA-seq
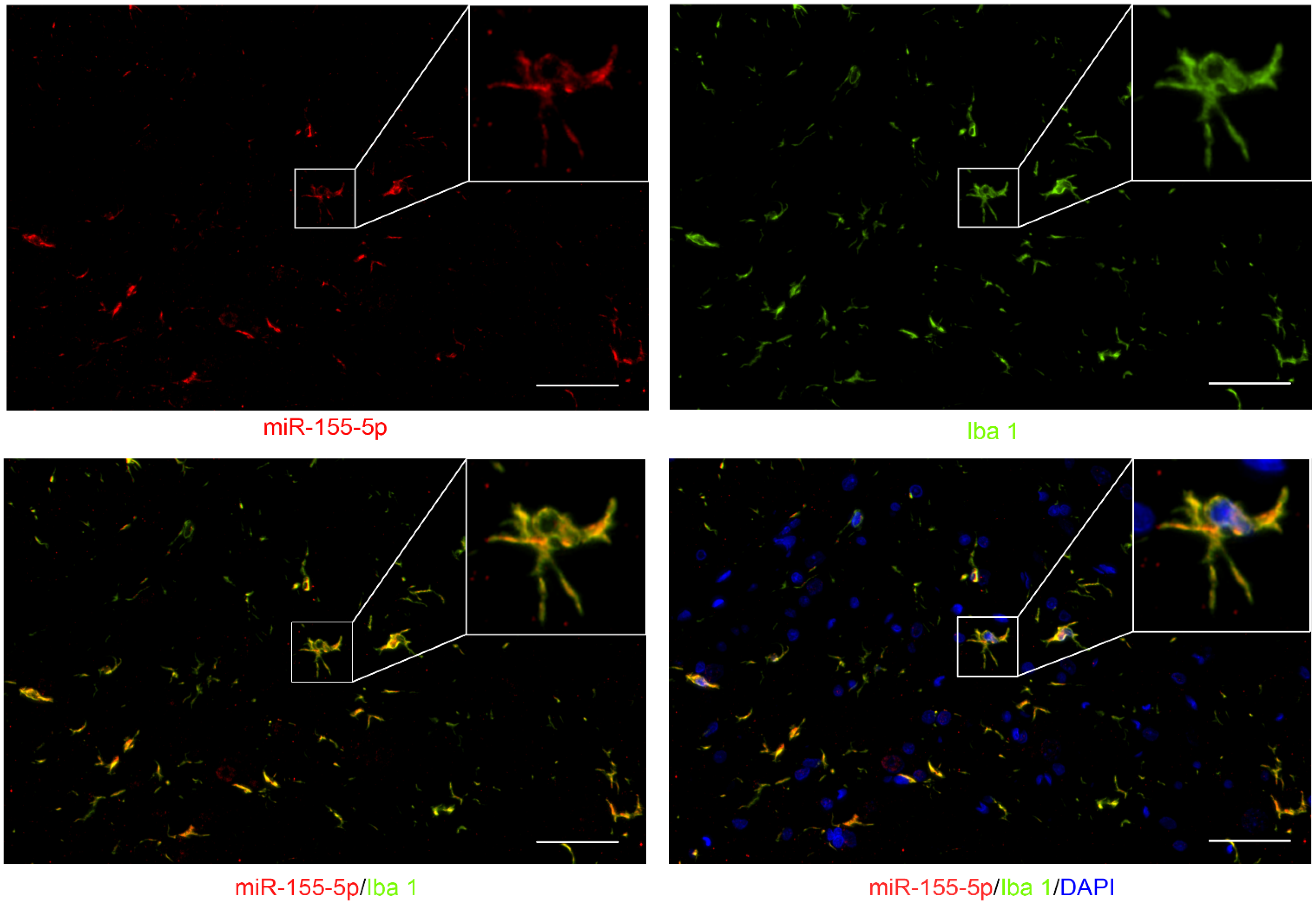
3.2. miR-155-5p Could Be Expressed in the Cytoplasm of Microglia in the Lumbar Spinal Cord of BCP Rats
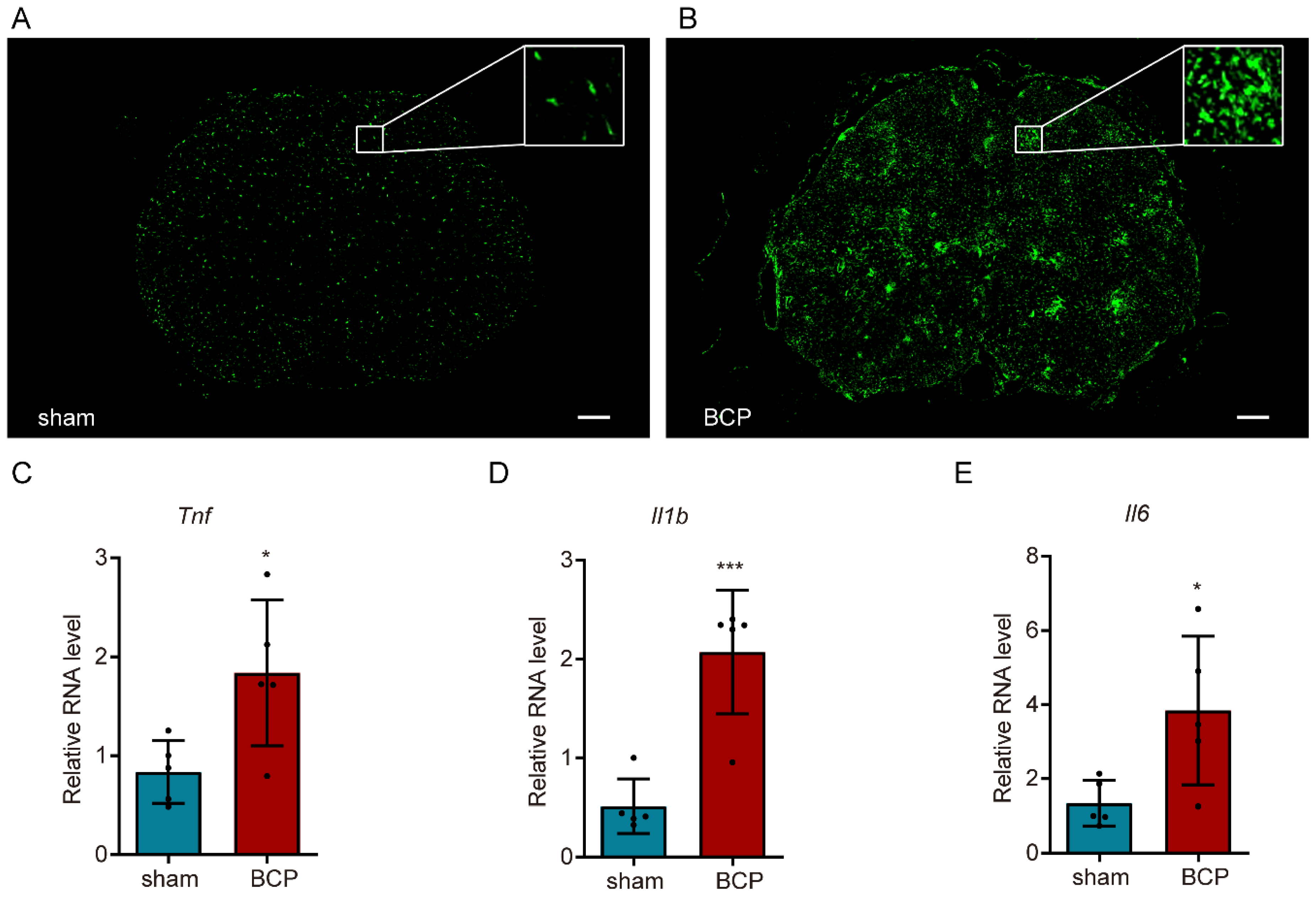
3.3. Microglia Were Dramatically Activated and Accompanied by the Increase in Inflammatory Cytokines in BCP Rats
3.4. The Expressions of miR-155-5p, Tnf, Il1b, and Il6 Were Increased in N9 Cells Treated with LPS
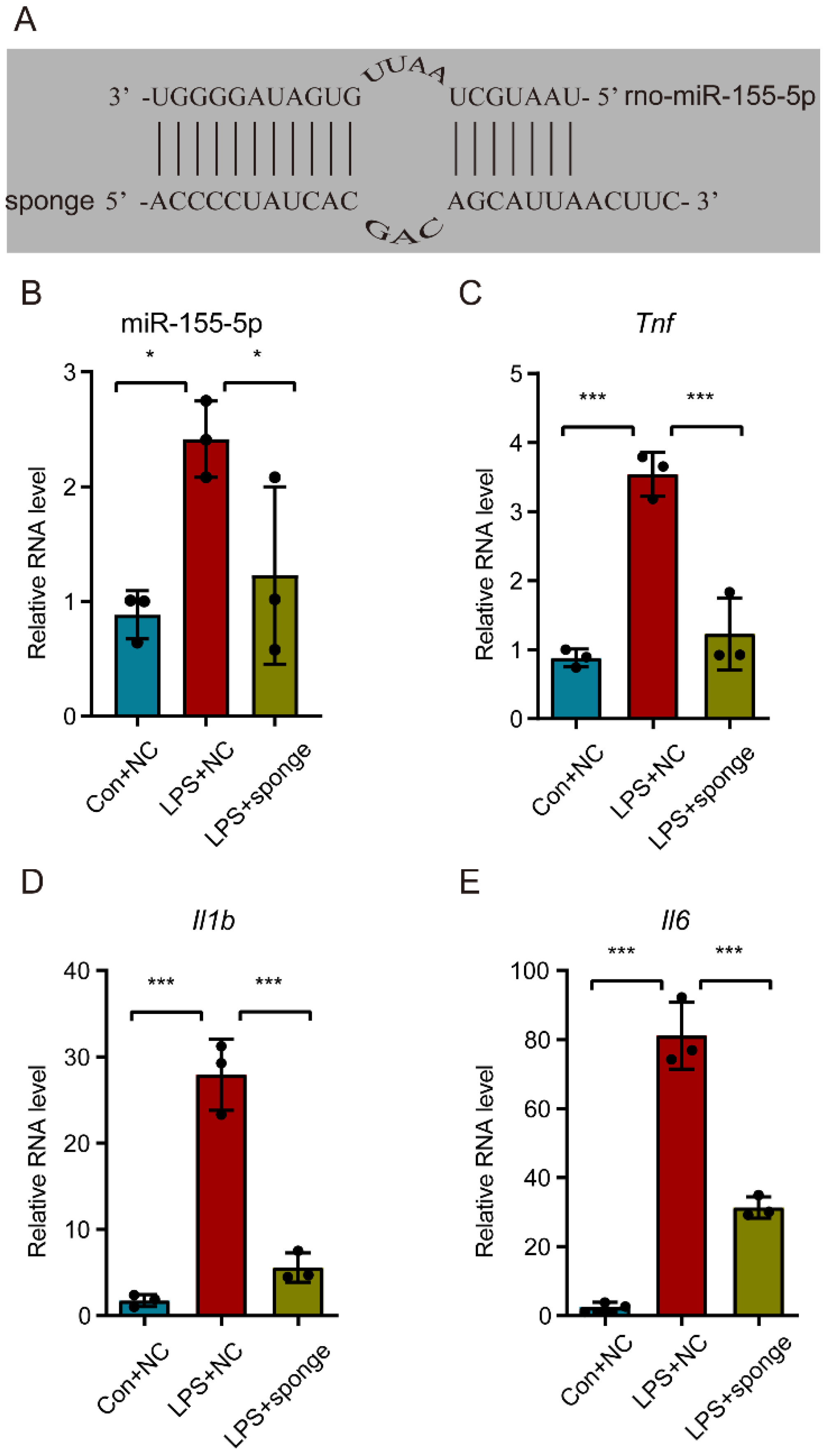
3.5. Knockdown of miR-155-5p Alleviated the Upregulation of Tnf, Il1b, and Il6 in LPS-Treated N9 Cells In Vitro
3.6. Sgk3 May Have Been a Direct Target of miR-155-5p In Vitro
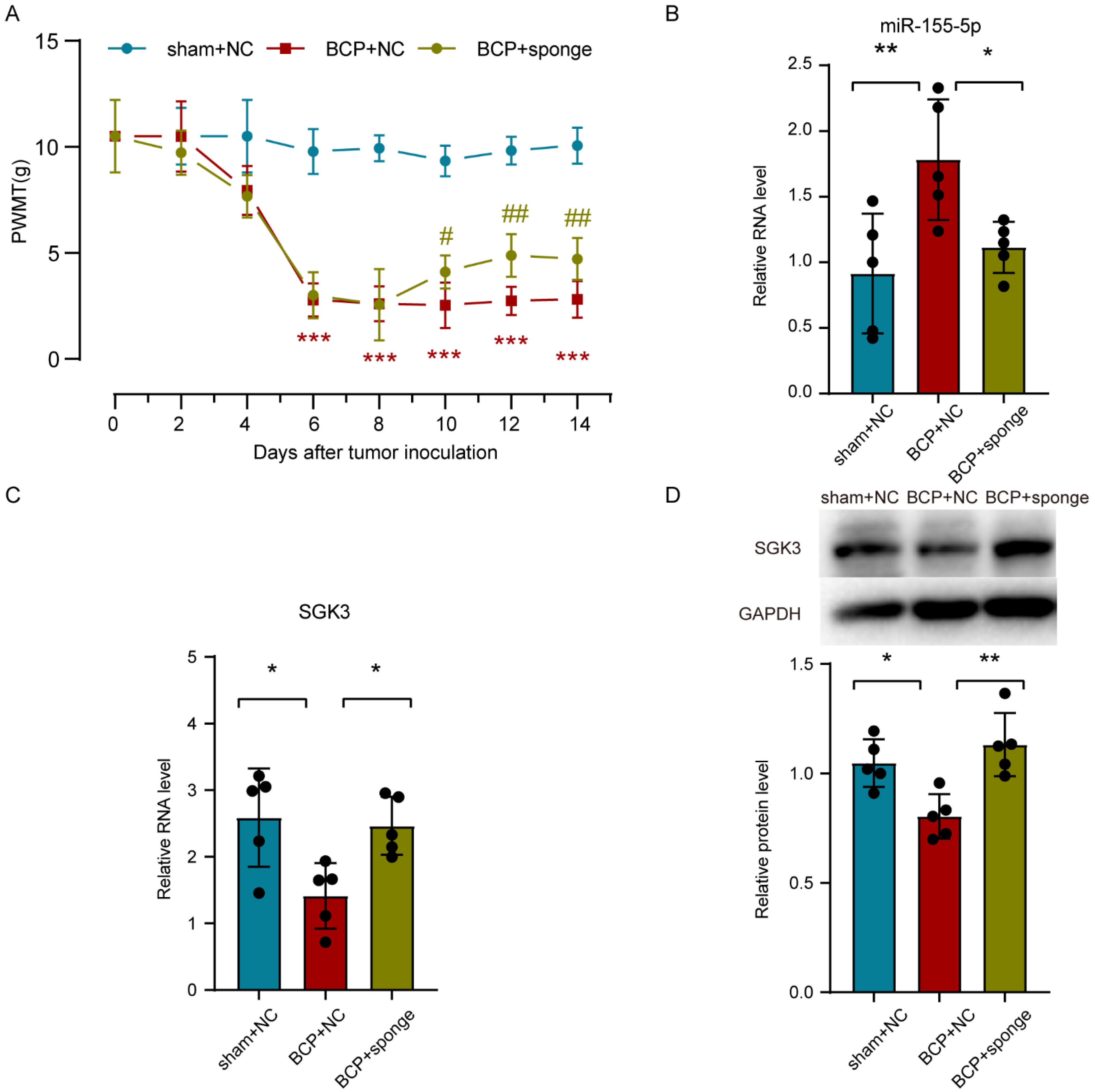
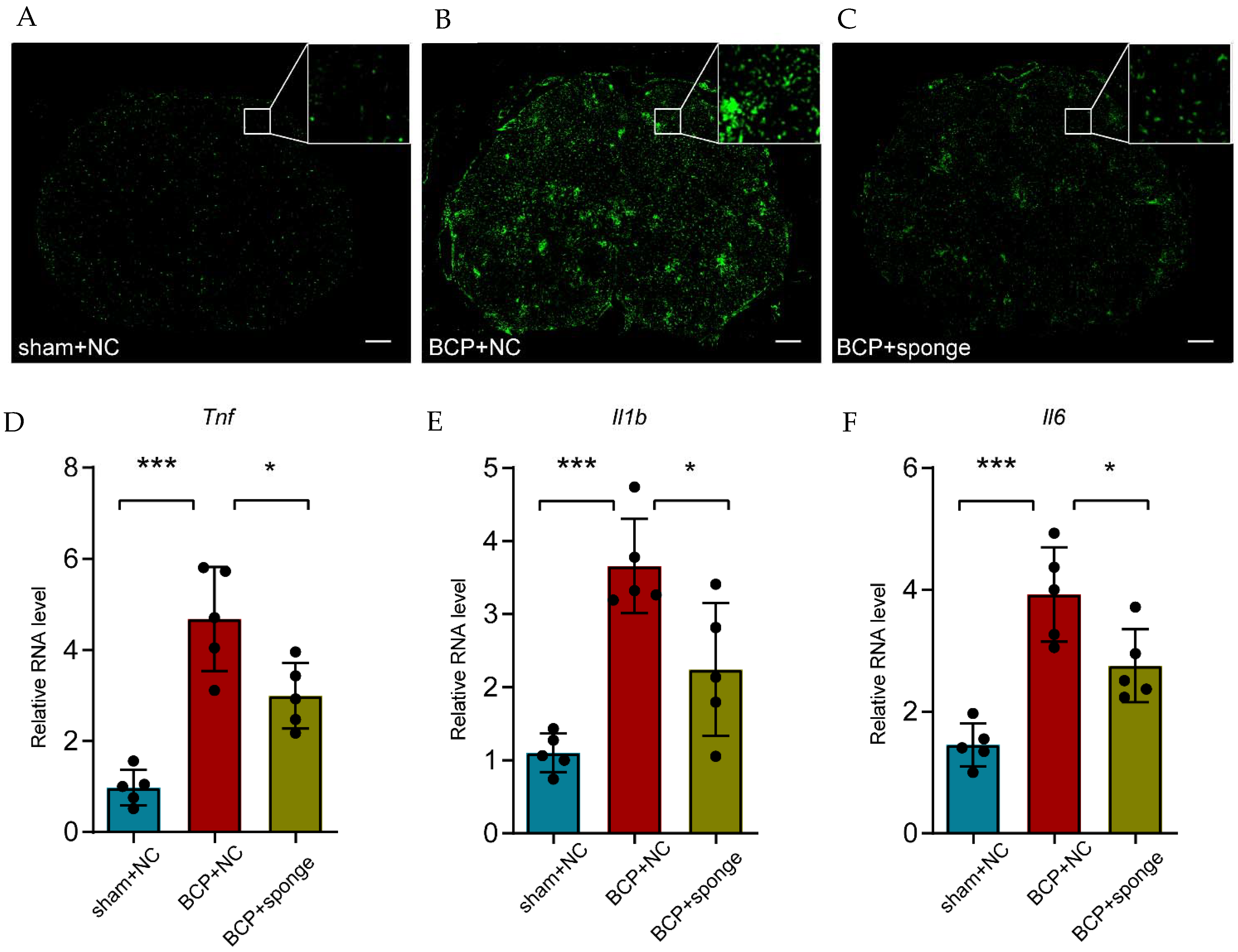
3.7. Inhibiting miR-155-5p Altered the Expression of Sgk3, Alleviated Spinal Cord Inflammation, and Attenuated Mechanical Hyperalgesia in BCP Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, M.; Kaasa, S.; Barke, A.; Korwisi, B.; Rief, W.; Treede, R. The IASP Classification of Chronic Pain for ICD-11: Chronic Cancer-Related Pain. Pain 2019, 160, 38–44. [Google Scholar] [CrossRef]
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wordliczek, J. Bone Pain in Cancer Patients: Mechanisms and Current Treatment. Int. J. Mol. Sci. 2019, 20, 6047. [Google Scholar] [CrossRef]
- Hernandez, R.K.; Wade, S.W.; Reich, A.; Pirolli, M.; Liede, A.; Lyman, G.H. Incidence of Bone Metastases in Patients with Solid Tumors: Analysis of Oncology Electronic Medical Records in the United States. BMC Cancer 2018, 18, 44. [Google Scholar] [CrossRef]
- Othman, A.; Winogradzki, M.; Lee, L.; Tandon, M.; Blank, A.; Pratap, J. Bone Metastatic Breast Cancer: Advances in Cell Signaling and Autophagy Related Mechanisms. Cancers 2021, 13, 4310. [Google Scholar] [CrossRef]
- Kane, C.M.; Hoskin, P.; Bennett, M.I. Cancer Induced Bone Pain. BMJ 2015, 350, h315. [Google Scholar] [CrossRef]
- Aielli, F.; Ponzetti, M.; Rucci, N. Bone Metastasis Pain, From the Bench to the Bedside. Int. J. Mol. Sci. 2019, 20, 280. [Google Scholar] [CrossRef]
- López-González, M.; Landry, M.; Favereaux, A. MicroRNA and Chronic Pain: From Mechanisms to Therapeutic Potential. Pharmacol. Ther. 2017, 180, 1–15. [Google Scholar] [CrossRef]
- Elramah, S.; López-González, M.J.; Bastide, M.; Dixmérias, F.; Roca-Lapirot, O.; Wielanek-Bachelet, A.C.; Vital, A.; Leste-Lasserre, T.; Brochard, A.; Landry, M.; et al. Spinal Mirna-124 Regulates Synaptopodin and Nociception in an Animal Model of Bone Cancer Pain. Sci. Rep. 2017, 7, 10949. [Google Scholar] [CrossRef]
- Liu, M.; Cheng, X.; Yan, H.; Chen, J.; Liu, C.; Chen, Z. MiR-135-5p Alleviates Bone Cancer Pain by Regulating Astrocyte-Mediated Neuroinflammation in Spinal Cord through JAK2/STAT3 Signaling Pathway. Mol. Neurobiol. 2021, 58, 4802–4815. [Google Scholar] [CrossRef]
- Tang, S.; Jing, H.; Song, F.; Huang, H.; Li, W.; Xie, G.; Zhou, J. MicroRNAs in the Spinal Microglia Serve Critical Roles in Neuropathic Pain. Mol. Neurobiol. 2021, 58, 132–142. [Google Scholar] [CrossRef]
- Wen, Q.; Wang, Y.; Pan, Q.; Tian, R.; Zhang, D.; Qin, G.; Zhou, J.; Chen, L. MicroRNA-155-5p Promotes Neuroinflammation and Central Sensitization Via Inhibiting SIRT1 in a Nitroglycerin-Induced Chronic Migraine Mouse Model. J. Neuroinflamm. 2021, 18, 287. [Google Scholar] [CrossRef]
- Duan, Z.; Zhang, J.; Li, J.; Pang, X.; Wang, H. Inhibition of microRNA-155 Reduces Neuropathic Pain during Chemotherapeutic Bortezomib via Engagement of Neuroinflammation. Front. Oncol. 2020, 10, 416. [Google Scholar] [CrossRef]
- Tan, Y.; Yang, J.; Xiang, K.; Tan, Q.; Guo, Q. Suppression of Microrna-155 Attenuates Neuropathic Pain by Regulating SOCS1 Signalling Pathway. Neurochem. Res. 2015, 40, 550–560. [Google Scholar] [CrossRef]
- Chen, J.; Li, C.; Liu, W.; Yan, B.; Hu, X.; Yang, F. Mirna-155 Silencing Reduces Sciatic Nerve Injury in Diabetic Peripheral Neuropathy. J. Mol. Endocrinol. 2019, 63, 227–238. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Fonken, L.K.; Ayala, M.T.; Maier, S.F.; Watkins, L.R. Aging and Mir-155 in Mice Influence Survival and Neuropathic Pain after Spinal Cord Injury. Brain Behav. Immun. 2021, 97, 365–370. [Google Scholar] [CrossRef]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the Pathophysiology of Neuropathic Pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef]
- Mao, Y.; Wang, C.; Tian, X.; Huang, Y.; Zhang, Y.; Wu, H.; Yang, S.; Xu, K.; Liu, Y.; Zhang, W.; et al. Endoplasmic Reticulum Stress Contributes to Nociception via Neuroinflammation in a Murine Bone Cancer Pain Model. Anesthesiology 2020, 132, 357–372. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.; Qadri, Y.; Serhan, C.; Ji, R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef]
- Inoue, K.; Tsuda, M. Microglia in Neuropathic Pain: Cellular and Molecular Mechanisms and Therapeutic Potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef]
- Tansley, S.; Gu, N.; Guzmán, A.U.; Cai, W.; Wong, C.; Lister, K.C.; Muñoz-Pino, E.; Yousefpour, N.; Roome, R.B.; Heal, J.; et al. Microglia-Mediated Degradation of Perineuronal Nets Promotes Pain. Science 2022, 377, 80–86. [Google Scholar] [CrossRef]
- Yang, Y.; Li, H.; Li, T.T.; Luo, H.; Gu, X.Y.; Lu, N.; Ji, R.R.; Zhang, Y.Q. Delayed Activation of Spinal Microglia Contributes to the Maintenance of Bone Cancer Pain in Female Wistar Rats Via P2X7 Receptor And IL-18. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 7950–7963. [Google Scholar] [CrossRef]
- Huo, W.; Liu, Y.; Lei, Y.; Zhang, Y.; Huang, Y.; Mao, Y.; Wang, C.; Sun, Y.; Zhang, W.; Ma, Z.; et al. Imbalanced Spinal Infiltration of Th17/Treg Cells Contributes to Bone Cancer Pain via Promoting Microglial Activation. Brain Behav. Immun. 2019, 79, 139–151. [Google Scholar] [CrossRef]
- Apryani, E.; Ali, U.; Wang, Z.Y.; Wu, H.Y.; Mao, X.F.; Ahmad, K.A.; Li, X.Y.; Wang, Y.X. The Spinal Microglial IL-10/Β-Endorphin Pathway Accounts for Cinobufagin-Induced Mechanical Antiallodynia in Bone Cancer Pain Following Activation of A7-Nicotinic Acetylcholine Receptors. J. Neuroinflamm. 2020, 17, 75. [Google Scholar] [CrossRef]
- Diaz-delCastillo, M.; Hansen, R.B.; Appel, C.K.; Nielsen, L.; Nielsen, S.N.; Karyniotakis, K.; Dahl, L.M.; Andreasen, R.B.; Heegaard, A.M. Modulation of Rat Cancer-Induced Bone Pain Is Independent of Spinal Microglia Activity. Cancers 2020, 12, 2740. [Google Scholar] [CrossRef]
- Ducourneau, V.R.R.; Dolique, T.; Hachem-Delaunay, S.; Miraucourt, L.S.; Amadio, A.; Blaszczyk, L.; Jacquot, F.; Ly, J.; Devoize, L.; Oliet, S.H.R.; et al. Cancer Pain Is not Necessarily Correlated with Spinal Overexpression of Reactive Glia Markers. Pain 2014, 155, 275–291. [Google Scholar] [CrossRef]
- Hou, X.; Weng, Y.; Guo, Q.; Ding, Z.; Wang, J.; Dai, J.; Wei, A.; Song, Z. Transcriptomic Analysis of Long Noncoding Rnas and Mrnas Expression Profiles in the Spinal Cord of Bone Cancer Pain Rats. Mol. Brain 2020, 13, 47. [Google Scholar] [CrossRef]
- Hou, X.; Weng, Y.; Ouyang, B.; Ding, Z.; Song, Z.; Zou, W.; Huang, C.; Guo, Q. HDAC Inhibitor TSA Ameliorates Mechanical Hypersensitivity and Potentiates Analgesic Effect of Morphine in a Rat Model of Bone Cancer Pain by Restoring Μ-Opioid Receptor in Spinal Cord. Brain Res. 2017, 1669, 97–105. [Google Scholar] [CrossRef]
- Hou, X.; Weng, Y.; Wang, T.; Ouyang, B.; Li, Y.; Song, Z.; Pan, Y.; Zhang, Z.; Zou, W.; Huang, C.; et al. Suppression of HDAC2 in Spinal Cord Alleviates Mechanical Hyperalgesia and Restores KCC2 Expression in a Rat Model of Bone Cancer Pain. Neuroscience 2018, 377, 138–149. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting Effective Microrna Target Sites in Mammalian Mrnas. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, B.; Sun, Y.; Xie, X. Mir-155 Modulates the Progression of Neuropathic Pain through Targeting SGK3. Int. J. Clin. Exp. Pathol. 2015, 8, 14374–14382. [Google Scholar]
- Zheng, X.Q.; Wu, Y.H.; Huang, J.F.; Wu, A.M. Neurophysiological Mechanisms of Cancer-Induced Bone Pain. J. Adv. Res. 2022, 35, 117–127. [Google Scholar] [CrossRef]
- Bali, K.K.; Selvaraj, D.; Satagopam, V.P.; Lu, J.; Schneider, R.; Kuner, R. Genome-Wide Identification and Functional Analyses of Microrna Signatures Associated with Cancer Pain. EMBO Mol. Med. 2013, 5, 1740–1758. [Google Scholar] [CrossRef]
- Gandla, J.; Lomada, S.K.; Lu, J.; Kuner, R.; Bali, K.K. Mir-34c-5p Functions as Pronociceptive Microrna in Cancer Pain by Targeting Cav2.3 Containing Calcium Channels. Pain 2017, 158, 1765–1779. [Google Scholar] [CrossRef]
- Wu, X.P.; Yang, Y.P.; She, R.X.; Xing, Z.M.; Chen, H.W.; Zhang, Y.W. Microrna-329 Reduces Bone Cancer Pain through the LPAR1-Dependent LPAR1/ERK Signal Transduction Pathway in Mice. Ther. Adv. Med. Oncol. 2019, 11, 1758835919875319. [Google Scholar] [CrossRef]
- Zingale, V.D.; Gugliandolo, A.; Mazzon, E. MiR-155: An Important Regulator of Neuroinflammation. Int. J. Mol. Sci. 2021, 23, 90. [Google Scholar] [CrossRef]
- Sierksma, A.; Lu, A.; Salta, E.; Vanden Eynden, E.; Callaerts-Vegh, Z.; D’Hooge, R.; Blum, D.; Buée, L.; Fiers, M.; De Strooper, B. Deregulation of Neuronal Mirnas Induced by Amyloid-Β or TAU Pathology. Mol. Neurodegener. 2018, 13, 54. [Google Scholar] [CrossRef]
- Lu, R.; Du, L.; Zhou, F.; Yuan, X.; Liu, X.; Zhang, L. Rosmarinic Acid Alleviates Inflammation, Apoptosis, and Oxidative Stress through Regulating miR-155-5p in a Mice Model of Parkinson’s Disease. ACS Chem. Neurosci. 2020, 11, 3259–3266. [Google Scholar] [CrossRef]
- Mycko, M.P.; Cichalewska, M.; Cwiklinska, H.; Selmaj, K.W. MiR-155-3p Drives the Development of Autoimmune Demyelination by Regulation of Heat Shock Protein 40. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 16504–16515. [Google Scholar] [CrossRef]
- Wu, X.; Wang, Y.; Yu, T.; Nie, E.; Hu, Q.; Wu, W.; Zhi, T.; Jiang, K.; Wang, X.; Lu, X.; et al. Blocking MIR155HG/Mir-155 Axis Inhibits Mesenchymal Transition in Glioma. Neuro-oncology 2017, 19, 1195–1205. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Pang, X.; Zhang, J.; Guan, Y. Role of Microrna-155 In Modifying Neuroinflammation and Γ-Aminobutyric Acid Transporters in Specific Central Regions after Post-Ischaemic Seizures. J. Cell. Mol. Med. 2019, 23, 5017–5024. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Q.; Nai, Y.; Cao, C. Suppression of Mir-155 Attenuates Neuropathic Pain by Inducing an M1 To M2 Switch in Microglia. Folia Neuropathol. 2020, 58, 70–82. [Google Scholar] [CrossRef]
- Heyn, J.; Luchting, B.; Hinske, L.C.; Hübner, M.; Azad, S.C.; Kreth, S. Mir-124a And Mir-155 Enhance Differentiation of Regulatory T Cells in Patients with Neuropathic Pain. J. Neuroinflamm. 2016, 13, 248. [Google Scholar] [CrossRef]
- Lu, C.; Liu, Y.; Sun, B.; Sun, Y.; Hou, B.; Zhang, Y.; Ma, Z.; Gu, X. Intrathecal Injection Of JWH-015 Attenuates Bone Cancer Pain via Time-Dependent Modification of Pro-Inflammatory Cytokines Expression and Astrocytes Activity in Spinal Cord. Inflammation 2015, 38, 1880–1890. [Google Scholar] [CrossRef]
- Wang, C.; Xu, K.; Wang, Y.; Mao, Y.; Huang, Y.; Liang, Y.; Liu, Y.; Hao, J.; Gu, X.; Ma, Z.; et al. Spinal Cannabinoid Receptor 2 Activation Reduces Hypersensitivity Associated with Bone Cancer Pain and Improves the Integrity of the Blood-Spinal Cord Barrier. Reg. Anesth. Pain Med. 2020, 45, 783–791. [Google Scholar] [CrossRef]
- Hu, X.F.; He, X.T.; Zhou, K.X.; Zhang, C.; Zhao, W.J.; Zhang, T.; Li, J.L.; Deng, J.P.; Dong, Y.L. The Analgesic Effects of Triptolide in the Bone Cancer Pain Rats via Inhibiting the Upregulation of Hdacs in Spinal Glial Cells. J. Neuroinflamm. 2017, 14, 213. [Google Scholar] [CrossRef]
- Butovsky, O.; Jedrychowski, M.P.; Cialic, R.; Krasemann, S.; Murugaiyan, G.; Fanek, Z.; Greco, D.J.; Wu, P.M.; Doykan, C.E.; Kiner, O.; et al. Targeting Mir-155 Restores Abnormal Microglia and Attenuates Disease in SOD1 Mice. Ann. Neurol. 2015, 77, 75–99. [Google Scholar] [CrossRef]
- Moore, C.S.; Rao, V.T.; Durafourt, B.A.; Bedell, B.J.; Ludwin, S.K.; Bar-Or, A.; Antel, J.P. Mir-155 As A Multiple Sclerosis-Relevant Regulator Of Myeloid Cell Polarization. Ann. Neurol. 2013, 74, 709–720. [Google Scholar] [CrossRef]
- Thome, A.D.; Harms, A.S.; Volpicelli-Daley, L.A.; Standaert, D.G. MicroRNA-155 Regulates Alpha-Synuclein-Induced Inflammatory Responses in Models of Parkinson Disease. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 2383–2390. [Google Scholar] [CrossRef]
- Henry, R.J.; Doran, S.J.; Barrett, J.P.; Meadows, V.E.; Sabirzhanov, B.; Stoica, B.A.; Loane, D.J.; Faden, A.I. Inhibition of MiR-155 Limits Neuroinflammation and Improves Functional Recovery after Experimental Traumatic Brain Injury in Mice. Neurother. J. Am. Soc. Exp. NeuroTherapeutics 2019, 16, 216–230. [Google Scholar] [CrossRef]
- Woodbury, M.E.; Freilich, R.W.; Cheng, C.J.; Asai, H.; Ikezu, S.; Boucher, J.D.; Slack, F.; Ikezu, T. MiR-155 Is Essential for Inflammation-Induced Hippocampal Neurogenic Dysfunction. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 9764–9781. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ma, J.; Ding, G.; Gong, Q.; Wang, Y.; Yu, H.; Cheng, X. Microglia Polarization from M1 toward M2 Phenotype Is Promoted by Astragalus Polysaccharides Mediated through Inhibition of miR-155 in Experimental Autoimmune Encephalomyelitis. Oxidative Med. Cell. Longev. 2021, 2021, 5753452. [Google Scholar] [CrossRef]
- Dou, L.; Lin, H.; Wang, K.; Zhu, G.; Zou, X.; Chang, E.; Zhu, Y. Long Non-Coding RNA CCAT1 Modulates Neuropathic Pain Progression through Sponging Mir-155. Oncotarget 2017, 8, 89949–89957. [Google Scholar] [CrossRef]
- Lang, F.; Bohmer, C.; Palmada, M.; Seebohm, G.; Strutz-Seebohm, N.; Vallon, V. (Patho)Physiological Significance of the Serum- and Glucocorticoid-Inducible Kinase Isoforms. Physiol. Rev. 2006, 86, 1151–1178. [Google Scholar] [CrossRef]
- Inoue, K.; Sakuma, E.; Morimoto, H.; Asai, H.; Koide, Y.; Leng, T.; Wada, I.; Xiong, Z.G.; Ueki, T. Serum- and Glucocorticoid-Inducible Kinases in Microglia. Biochem. Biophys. Res. Commun. 2016, 478, 53–59. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jian, Y.; Song, Z.; Ding, Z.; Wang, J.; Wang, R.; Hou, X. Upregulation of Spinal miR-155-5p Contributes to Mechanical Hyperalgesia by Promoting Inflammatory Activation of Microglia in Bone Cancer Pain Rats. Life 2022, 12, 1349. https://doi.org/10.3390/life12091349
Jian Y, Song Z, Ding Z, Wang J, Wang R, Hou X. Upregulation of Spinal miR-155-5p Contributes to Mechanical Hyperalgesia by Promoting Inflammatory Activation of Microglia in Bone Cancer Pain Rats. Life. 2022; 12(9):1349. https://doi.org/10.3390/life12091349
Chicago/Turabian StyleJian, Yanping, Zongbin Song, Zhuofeng Ding, Jian Wang, Ruike Wang, and Xinran Hou. 2022. "Upregulation of Spinal miR-155-5p Contributes to Mechanical Hyperalgesia by Promoting Inflammatory Activation of Microglia in Bone Cancer Pain Rats" Life 12, no. 9: 1349. https://doi.org/10.3390/life12091349
APA StyleJian, Y., Song, Z., Ding, Z., Wang, J., Wang, R., & Hou, X. (2022). Upregulation of Spinal miR-155-5p Contributes to Mechanical Hyperalgesia by Promoting Inflammatory Activation of Microglia in Bone Cancer Pain Rats. Life, 12(9), 1349. https://doi.org/10.3390/life12091349