
Dihydroartemisinin Reduces Irradiation-Induced Mitophagy and Radioresistance in Lung Cancer A549 Cells via CIRBP Inhibition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Reagents
2.2. Radiation-Induced Radioresistance of Lung Cancer A549 Cells
2.3. Clonogenic Assay
2.4. Real-Time Polymerase Chain Reaction (RT-PCR)
| Gene Name | Sequence (5′ → 3′) | |
| CIRBP | Forward | AGGGCTGAGTTTTGACACCAA |
| Reverse | ACAAACCCAAATCCCCGAGAT | |
| PINK1 | Forward | GCCTCATCGAGGAAAAACAGG |
| Reverse | GTCTCGTGTCCAACGGGTC | |
| Actin | Forward | GCACCACACCTTCTACAATGA |
| Reverse | GTCATCTTCTCGCGGTTGGC |
2.5. Western Blot
2.6. Protein Chip
2.7. Gene Knockdown
2.8. Immunofluorescence and Confocal Imaging
2.9. Flow Cytometry
2.10. Statistics
3. Results
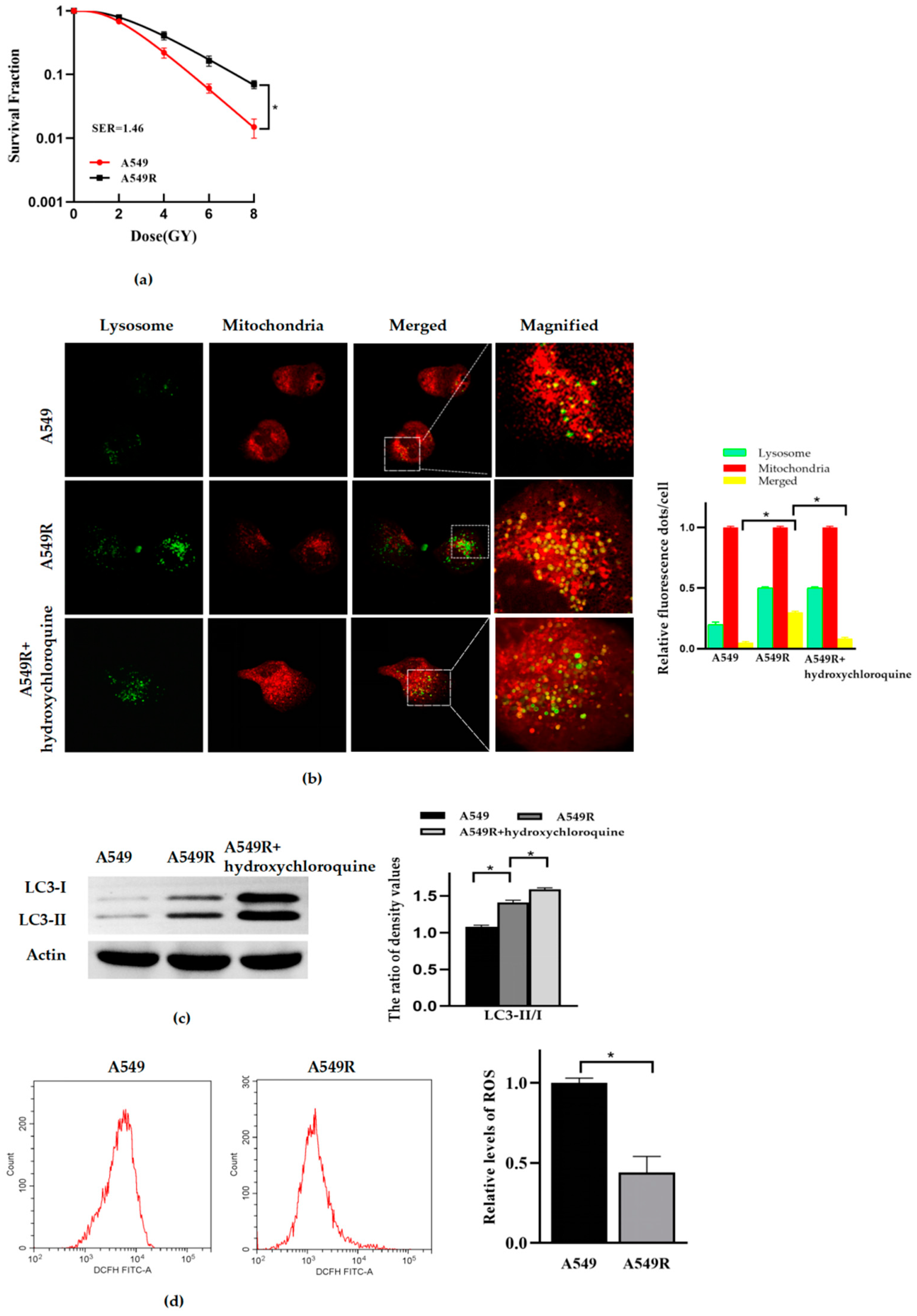
3.1. Mitophagy and Radioresistance Were Induced by Fraction Radiation in Lung Cancer A549 Cells
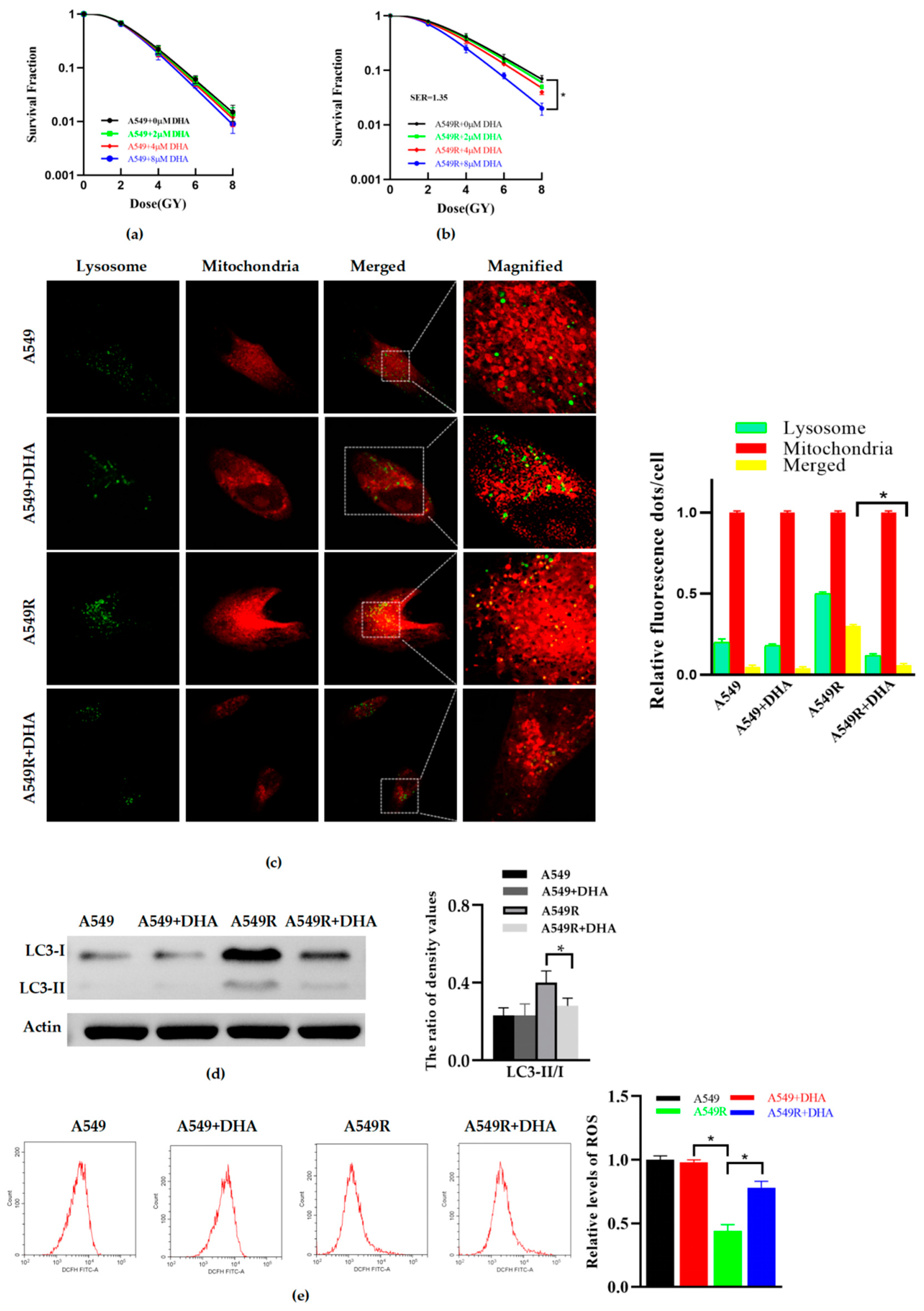
3.2. DHA Reduces Radioresistance and Mitophagy in A549R Cells
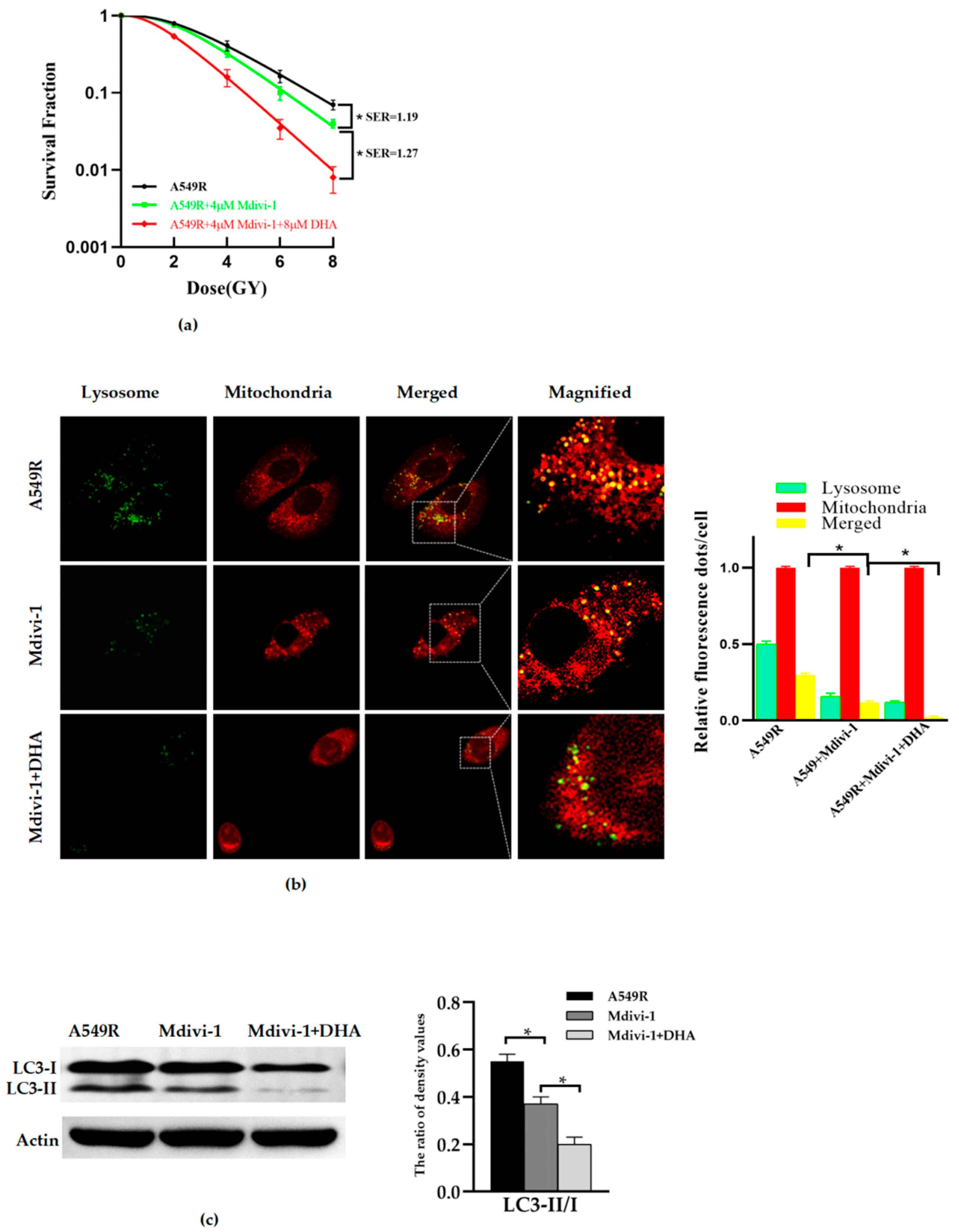
3.3. Inhibition of Mitophagy Reduces the Radioresistance of A549R Cells
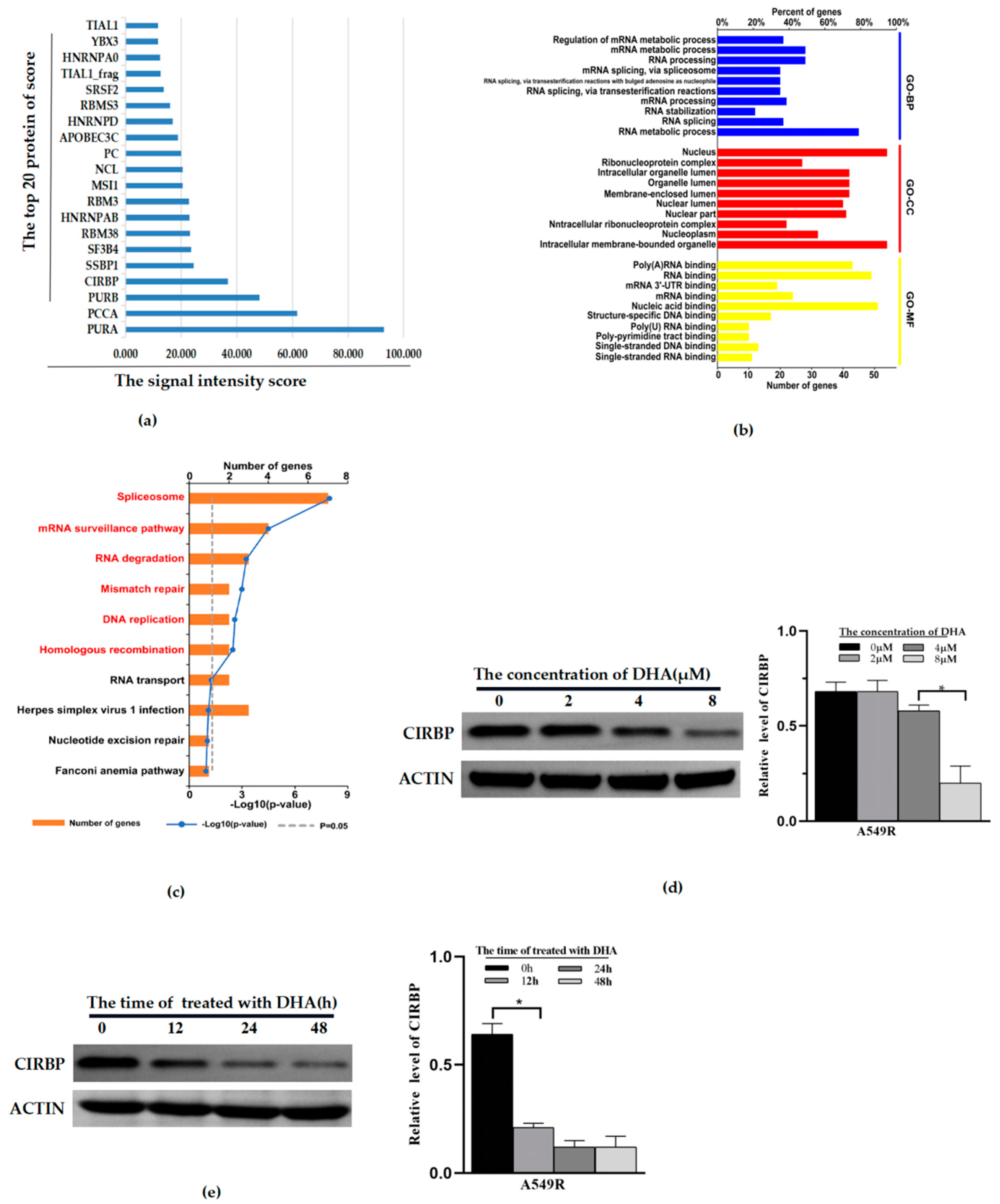
3.4. CIRBP Identified as a Potential Target of DHA-Reducing Mitophagy and Radioresistance
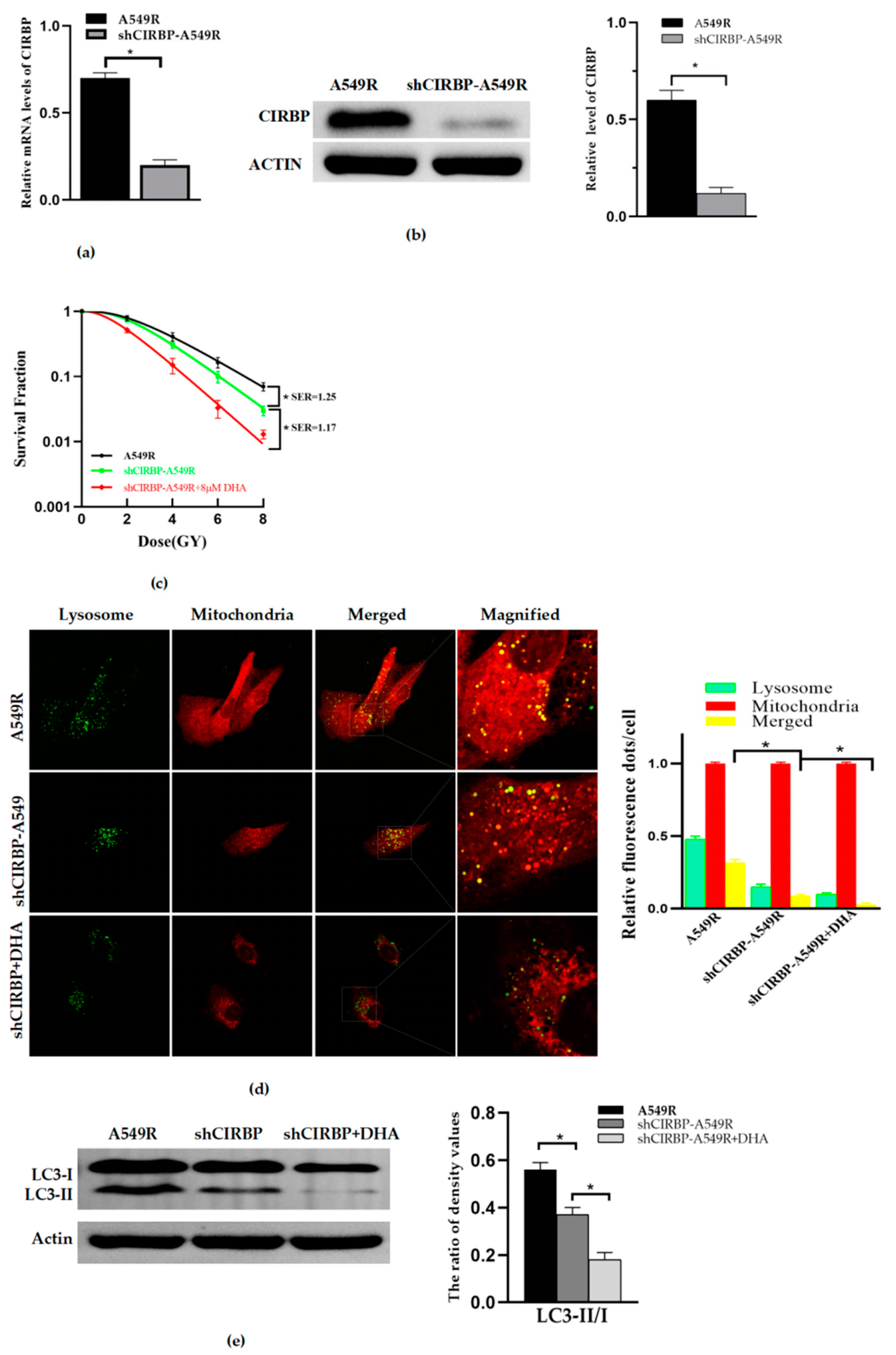
3.5. Knockdown of CIRBP Inhibited the Mitophagy and Radioresistance of A549R
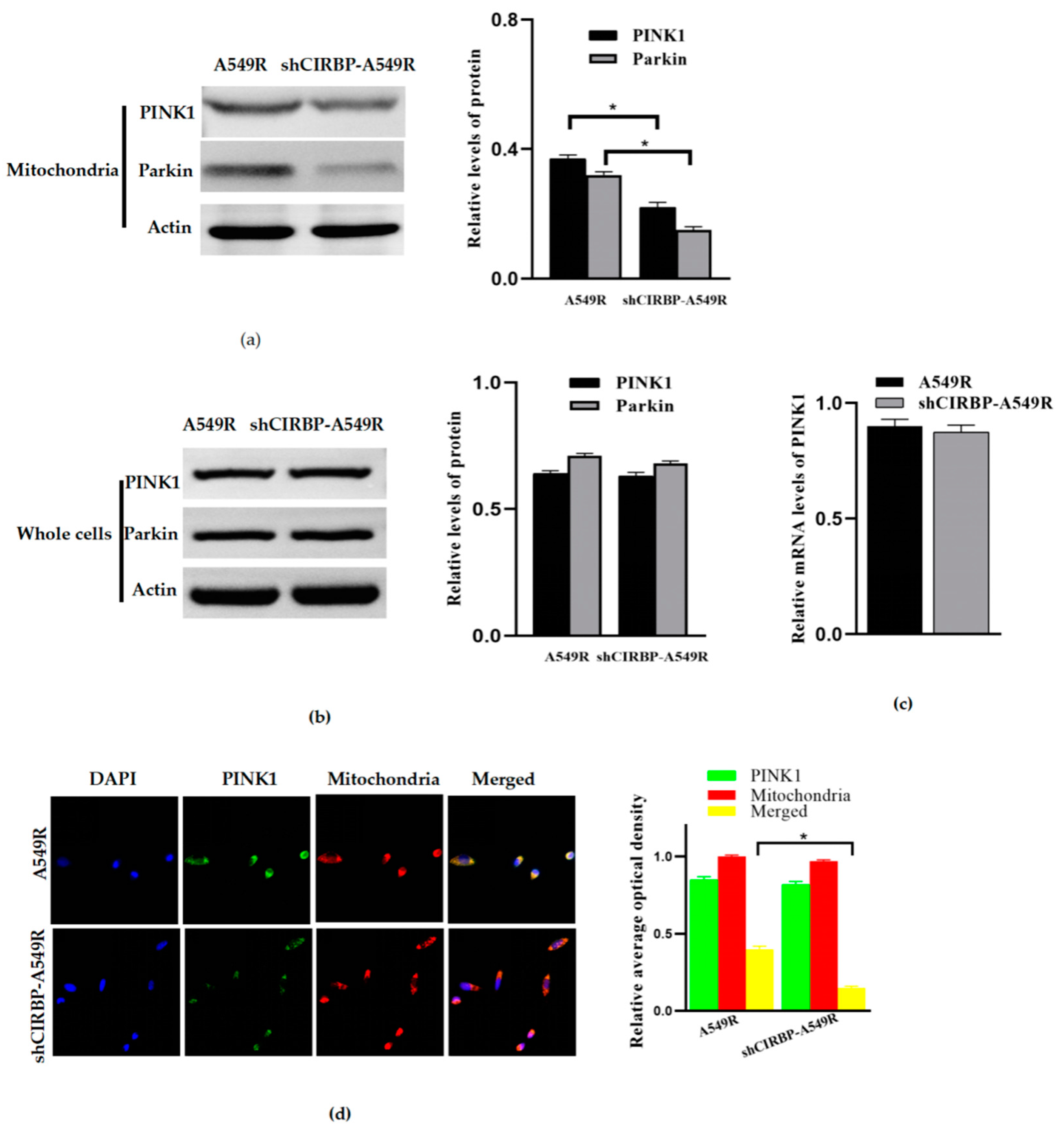
3.6. Knockdown of CIRBP Inhibited the PINK1/Parkin Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, S.; Banfill, K.; Aznar, M.C.; Whitehurst, P.; Finn, C.F. The evolving role of radiotherapy in non-small cell lung cancer. Br. J. Radiol. 2019, 92, 20190524. [Google Scholar] [CrossRef]
- Vinod, S.K.; Hau, E. Radiotherapy treatment for lung cancer: Current status and future directions. Respirology 2020, 25 (Suppl. 2), 61–71. [Google Scholar] [CrossRef] [PubMed]
- Shafifiq, J.; Hanna, T.P.; Vinod, S.K.; Delaney, G.P.; Barton, M.B. A population-based model of local control and survival benefifit of radiotherapy for lung cancer. Clin. Oncol. 2016, 28, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, D.S.; Wood, D.E.; Aisner, D.L.; Akerley, W.; Bauman, J.R.; Bharat, A.; Bruno, D.S.; Chang, J.Y.; Chirieac, L.R.; D’Amico, T.A.; et al. NCCN Guidelines Insights: Non-Small Cell Lung Cancer, Version 2.2021. J. Natl. Compr. Cancer Netw. JNCCN 2021, 19, 254–266. [Google Scholar] [CrossRef]
- Kim, B.M.; Hong, Y.; Lee, S.; Liu, P.; Lim, J.H.; Lee, Y.H.; Lee, T.H.; Chang, K.T.; Hong, Y. Therapeutic Implications for Overcoming Radiation Resistance in Cancer Therapy. Int. J. Mol. Sci. 2015, 16, 26880–26913. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ma, Q.; Cheng, J.; Zhou, X.; Pu, W.; Zhong, X.; Guo, X. Dihydroartemisinin as a Sensitizing Agent in Cancer Therapies. OncoTargets Ther. 2021, 14, 2563–2573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, F.; Wang, Y.; Xie, H.; Luo, S.; Meng, L.; Su, B.; Ye, Y.; Wu, K.; Xu, Y.; et al. Eliminating Radiation Resistance of Non-Small Cell Lung Cancer by Dihydroartemisinin Through Abrogating Immunity Escaping and Promoting Radiation Sensitivity by Inhibiting PD-L1 Expression. Front. Oncol. 2020, 10, 595466. [Google Scholar] [CrossRef]
- Liao, K.; Li, J.; Wang, Z. Dihydroartemisinin inhibits cell proliferation via AKT/GSK3β/cyclinD1 pathway and induces apoptosis in A549 lung cancer cells. Int. J. Clin. Exp. Pathol. 2014, 7, 8684–8691. [Google Scholar]
- Yu, L.; Yang, X.; Li, X.; Qin, L.; Xu, W.; Cui, H.; Jia, Z.; He, Q.; Wang, Z. Pink1/PARK2/mROS-Dependent Mitophagy Initiates the Sensitization of Cancer Cells to Radiation. Oxidative Med. Cell. Longev. 2021, 2021, 5595652. [Google Scholar] [CrossRef]
- Chen, Q.; Zheng, W.; Zhu, L.; Liu, H.; Song, Y.; Hu, S.; Bai, Y.; Pan, Y.; Zhang, J.; Guan, J.; et al. LACTB2 renders radioresistance by activating PINK1/Parkin-dependent mitophagy in nasopharyngeal carcinoma. Cancer Lett. 2021, 518, 127–139. [Google Scholar] [CrossRef]
- Li, S.; Lin, Q.; Shao, X.; Zhu, X.; Wu, J.; Wu, B.; Zhang, M.; Zhou, W.; Zhou, Y.; Jin, H.; et al. Drp1-regulated PARK2-dependent mitophagy protects against renal fibrosis in unilateral ureteral obstruction. Free Radic. Biol. Med. 2019, 152, 632–649. [Google Scholar] [CrossRef]
- Li, Y.J.; Wu, S.L.; Lu, S.M.; Chen, F.; Guo, Y.; Gan, S.M.; Shi, Y.L.; Liu, S.; Li, S.L. (-)-Epigallocatechin-3-gallate inhibits nasopharyngeal cancer stem cell self-renewal and migration and reverses the epithelial–mesenchymal transition via NF-κB p65 inactivation. Tumor Biol. 2015, 36, 2747–2761. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, Y.-D.; Ma, R.-Y.; Chen, Y.-J.; Xiang, X.-M.; Hou, D.-Y.; Li, X.-H.; Huang, H.; Li, T.; Duan, C.-Y. Activated Drp1 regulates p62-mediated autophagic flux and aggravates inflammation in cerebral ischemia-reperfusion via the ROS-RIP1/RIP3-exosome axis. Mil. Med Res. 2022, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Li, N.; Li, K.; Tan, Y.; Han, J.; Lin, B.; Lai, W.; Liu, H.; Shi, Y.; Xi, Z.; et al. Ambient ozone exposure induces ROS related-mitophagy and pyroptosis via NLRP3 inflammasome activation in rat lung cells. Ecotoxicol. Environ. Saf. 2022, 240, 113663. [Google Scholar] [CrossRef]
- Li, J.-Y.; Zhang, K.; Xu, D.; Zhou, W.-C.; Fang, W.-Q.; Wan, Y.-Y.; Yan, D.-D.; Guo, M.-Y.; Tao, J.-X.; Yang, F.; et al. Mitochondrial Fission Is Required for Blue Light-Induced Apoptosis and Mitophagy in Retinal Neuronal R28 Cells. Front. Mol. Neurosci. 2018, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi-Birgani, M.; Teimoori, A.; Ghadiri, A.; Mansoury-Asl, H.; Danyaei, A.; Khanbabaei, H. Fractionated radiotherapy might induce epithelial-mesenchymal transition and radioresistance in a cellular context manner. J. Cell. Biochem. 2018, 120, 8601–8610. [Google Scholar] [CrossRef]
- Liu, L.; Shang, M.; Song, X.; Zhang, C.; Guo, W. Fractionated Irradiation Enhances Invasion and Migration by Inducing Epithelial-Mesenchymal Transition and Stemness-Like Properties in A549 Cells. Ann. Clin. Lab Sci. 2021, 51, 521–528. [Google Scholar]
- Antunes, F.; Erustes, A.G.; Costa, A.J.; Nascimento, A.C.; Bincoletto, C.; Ureshino, R.P.; Pereira, G.J.S.; Smaili, S.S. Autophagy and intermittent fasting: The connection for cancer therapy? Clinics 2018, 73, e814s. [Google Scholar] [CrossRef]
- Onishi, M.; Yamano, K.; Sato, M.; Matsuda, N.; Okamoto, K. Molecular mechanisms and physiological functions of mitophagy. EMBO J. 2021, 40, e104705. [Google Scholar] [CrossRef]
- Chen, G.-Q.; Benthani, F.A.; Wu, J.; Liang, D.; Bian, Z.-X.; Jiang, X. Artemisinin compounds sensitize cancer cells to ferroptosis by regulating iron homeostasis. Cell Death Differ. 2019, 27, 242–254. [Google Scholar] [CrossRef]
- Cui, Z.; Wang, H.; Li, S.; Qin, T.; Shi, H.; Ma, J.; Li, L.; Yu, G.; Jiang, T.; Li, C. Dihydroartemisinin enhances the inhibitory effect of sorafenib on HepG2 cells by inducing ferroptosis and inhibiting energy metabolism. J. Pharmacol. Sci. 2021, 148, 73–85. [Google Scholar] [CrossRef]
- Jin, H.; Jiang, A.-Y.; Wang, H.; Cao, Y.; Wu, Y.; Jiang, X.-F. Dihydroartemisinin and gefitinib synergistically inhibit NSCLC cell growth and promote apoptosis via the Akt/mTOR/STAT3 pathway. Mol. Med. Rep. 2017, 16, 3475–3481. [Google Scholar] [CrossRef] [Green Version]
- Bader, S.; Wilmers, J.; Pelzer, M.; Jendrossek, V.; Rudner, J. Activation of anti-oxidant Keap1/Nrf2 pathway modulates efficacy of dihydroartemisinin-based monotherapy and combinatory therapy with ionizing radiation. Free Radic. Biol. Med. 2021, 168, 44–54. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, X.; Li, C.; Zhang, X.; Li, C.L. Rapamycin promotes the anticancer action of dihydroartemisinin in breast cancer MDA-MB-231 cells by regulating expression of Atg7 and DAPK. Oncol. Lett. 2018, 15, 5781–5786. [Google Scholar] [CrossRef]
- Tai, X.; Cai, X.-B.; Zhang, Z.; Wei, R. In vitro and in vivo inhibition of tumor cell viability by combined dihydroartemisinin and doxorubicin treatment, and the underlying mechanism. Oncol. Lett. 2016, 12, 3701–3706. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-S.; Wang, J.; Shen, Y.-B.; Guo, C.-C.; Sai, K.; Chen, F.-R.; Mei, X.; Han, F.; Chen, Z.-P. Dihydroartemisinin increases temozolomide efficacy in glioma cells by inducing autophagy. Oncol. Lett. 2015, 10, 379–383. [Google Scholar] [CrossRef]
- Lujan, D.A.; Ochoa, J.L.; Hartley, R.S. Cold-inducible RNA binding protein in cancer and inflammation. Wiley Interdiscip. Rev. RNA 2018, 9. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Hong, S. Controversial roles of cold-inducible RNA-binding protein in human cancer (Review). Int. J. Oncol. 2021, 59, 91. [Google Scholar] [CrossRef]
- Indacochea, A.; Guerrero, S.; Ureña, M.; Araujo, F.; Coll, O.; Lleonart, M.E.; Gebauer, F. Cold-inducible RNA binding protein promotes breast cancer cell malignancy by regulating Cystatin C levels. RNA 2020, 27, 190–201. [Google Scholar] [CrossRef]
- Zeng, Y.; Kulkarni, P.; Inoue, T.; Getzenberg, R.H. Down-regulating cold shock protein genes impairs cancer cell survival and enhances chemosensitivity. J. Cell. Biochem. 2009, 107, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Feng, J.; Sun, W.; Wu, C.; Li, J.; Jing, T.; Liang, Y.; Qian, Y.; Liu, W.; Wang, H. CIRP promotes the progression of non-small cell lung cancer through activation of Wnt/β-catenin signaling via CTNNB1. J. Exp. Clin. Cancer Res. 2021, 40, 275. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Fan, E.K.; Liu, J.; Scott, M.J.; Li, Y.; Li, S.; Xie, W.; Billiar, T.R.; Wilson, M.A.; Jiang, Y.; et al. Cold-inducible RNA-binding protein through TLR4 signaling induces mitochondrial DNA fragmentation and regulates macrophage cell death after trauma. Cell Death Dis. 2017, 8, e2775. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, Y.; Cheng, X.; Yuan, H.; Zhu, S.; Liu, J.; Wen, Q.; Xie, Y.; Liu, J.; Kroemer, G.; et al. PINK1 and PARK2 Suppress Pancreatic Tumorigenesis through Control of Mitochondrial Iron-Mediated Immunometabolism. Dev. Cell 2018, 46, 441–455.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Li, Z.; Li, H.; Liao, K. Dihydroartemisinin Reduces Irradiation-Induced Mitophagy and Radioresistance in Lung Cancer A549 Cells via CIRBP Inhibition. Life 2022, 12, 1129. https://doi.org/10.3390/life12081129
Wu S, Li Z, Li H, Liao K. Dihydroartemisinin Reduces Irradiation-Induced Mitophagy and Radioresistance in Lung Cancer A549 Cells via CIRBP Inhibition. Life. 2022; 12(8):1129. https://doi.org/10.3390/life12081129
Chicago/Turabian StyleWu, Shunlong, Zhaodong Li, Haiyu Li, and Kui Liao. 2022. "Dihydroartemisinin Reduces Irradiation-Induced Mitophagy and Radioresistance in Lung Cancer A549 Cells via CIRBP Inhibition" Life 12, no. 8: 1129. https://doi.org/10.3390/life12081129
APA StyleWu, S., Li, Z., Li, H., & Liao, K. (2022). Dihydroartemisinin Reduces Irradiation-Induced Mitophagy and Radioresistance in Lung Cancer A549 Cells via CIRBP Inhibition. Life, 12(8), 1129. https://doi.org/10.3390/life12081129