
Prospective Role of Bioactive Molecules and Exosomes in the Therapeutic Potential of Camel Milk against Human Diseases: An Updated Perspective


 ,
,  , , , , ,
, , , , ,  ,
,  , , ,
, , , 

Abstract
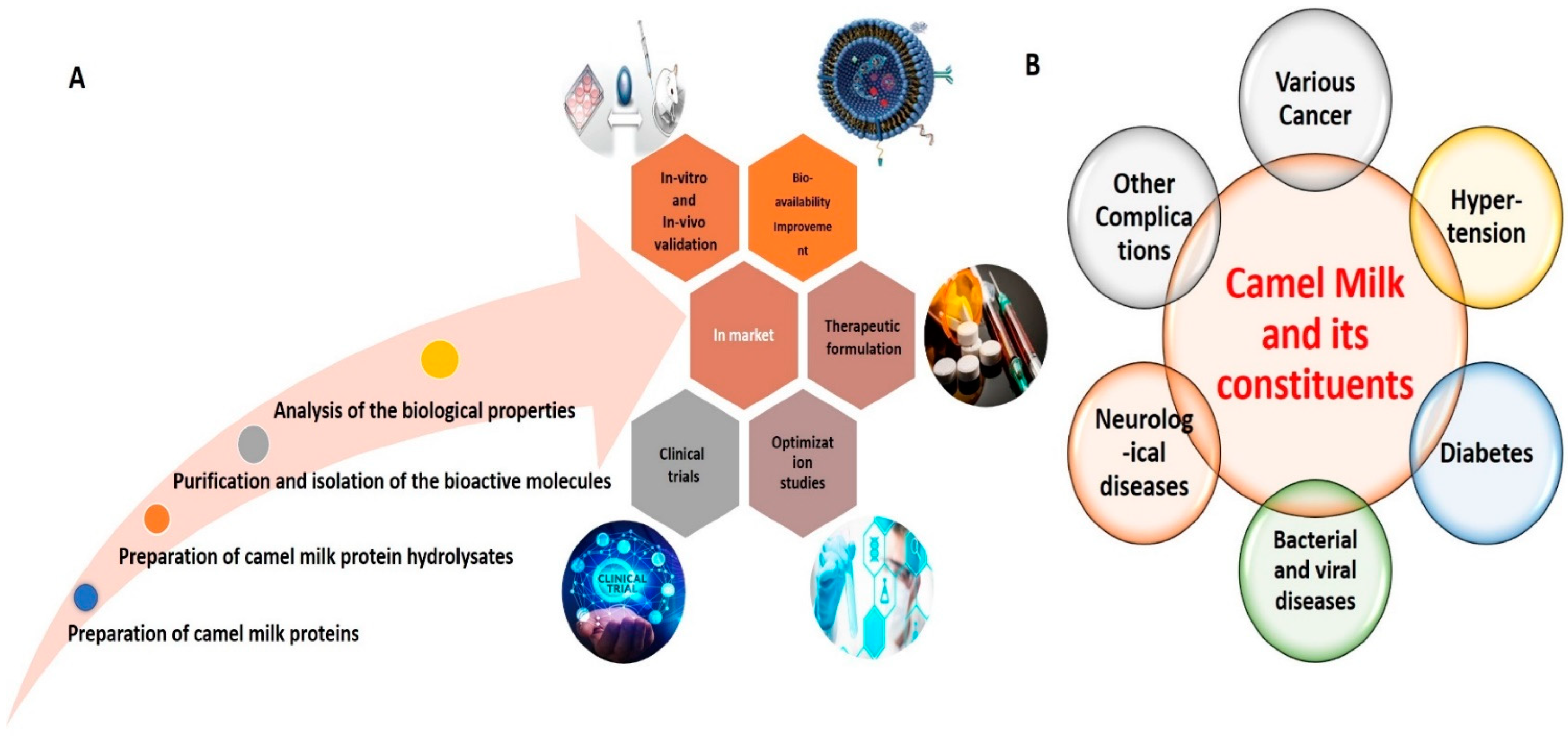
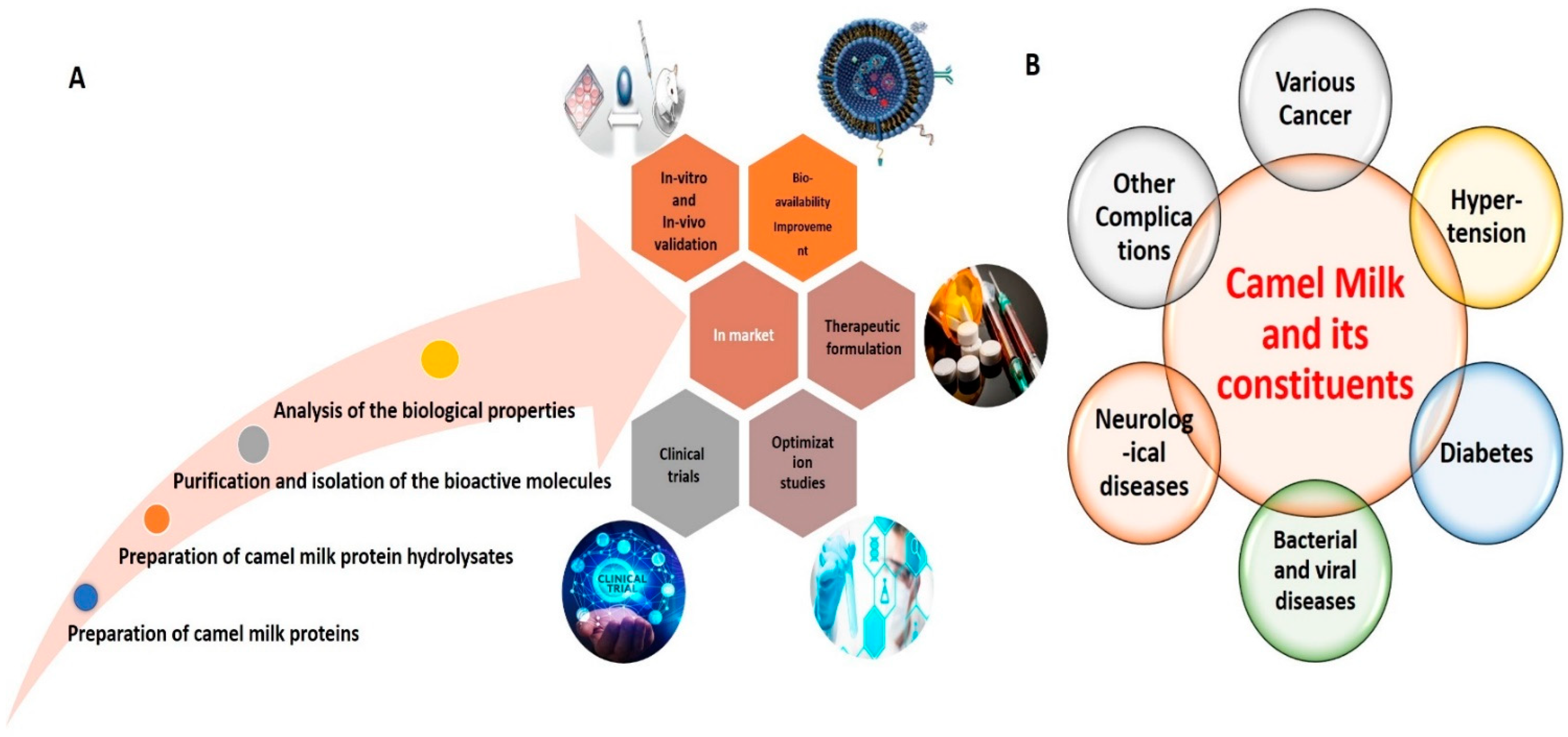
:1. Introduction
2. Brief Overview of Camel Milk (CM) Components
2.1. Nutritional Components (Macronutrients and Micronutrients)
2.2. Pharmacological Properties of CM Bioactive Molecules against Various Pathological Conditions
2.2.1. Molecular Intricacies of Anti-Cancer Effects
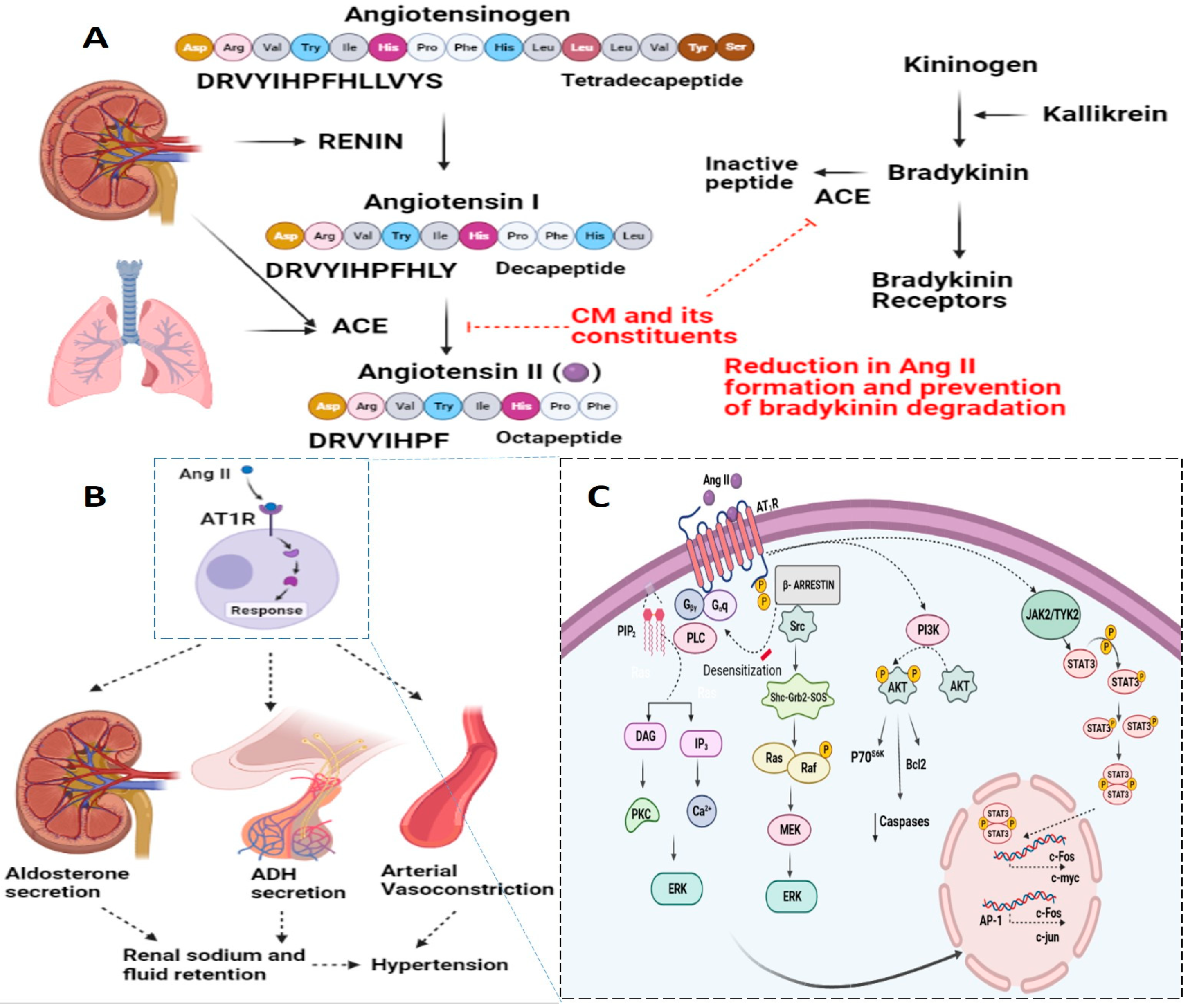
2.2.2. Molecular Intricacies of CM’s Anti-Hypertensive Potential
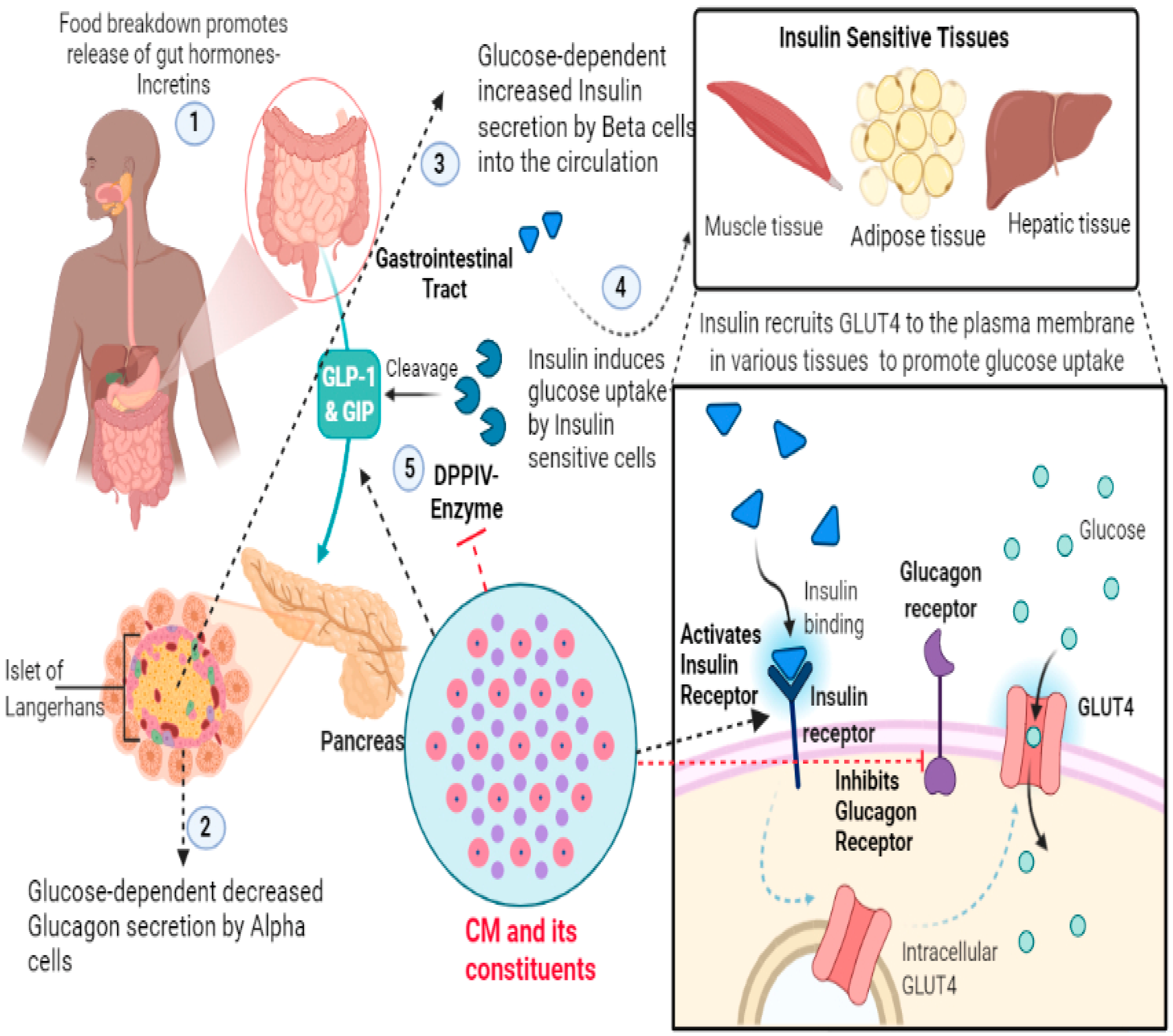
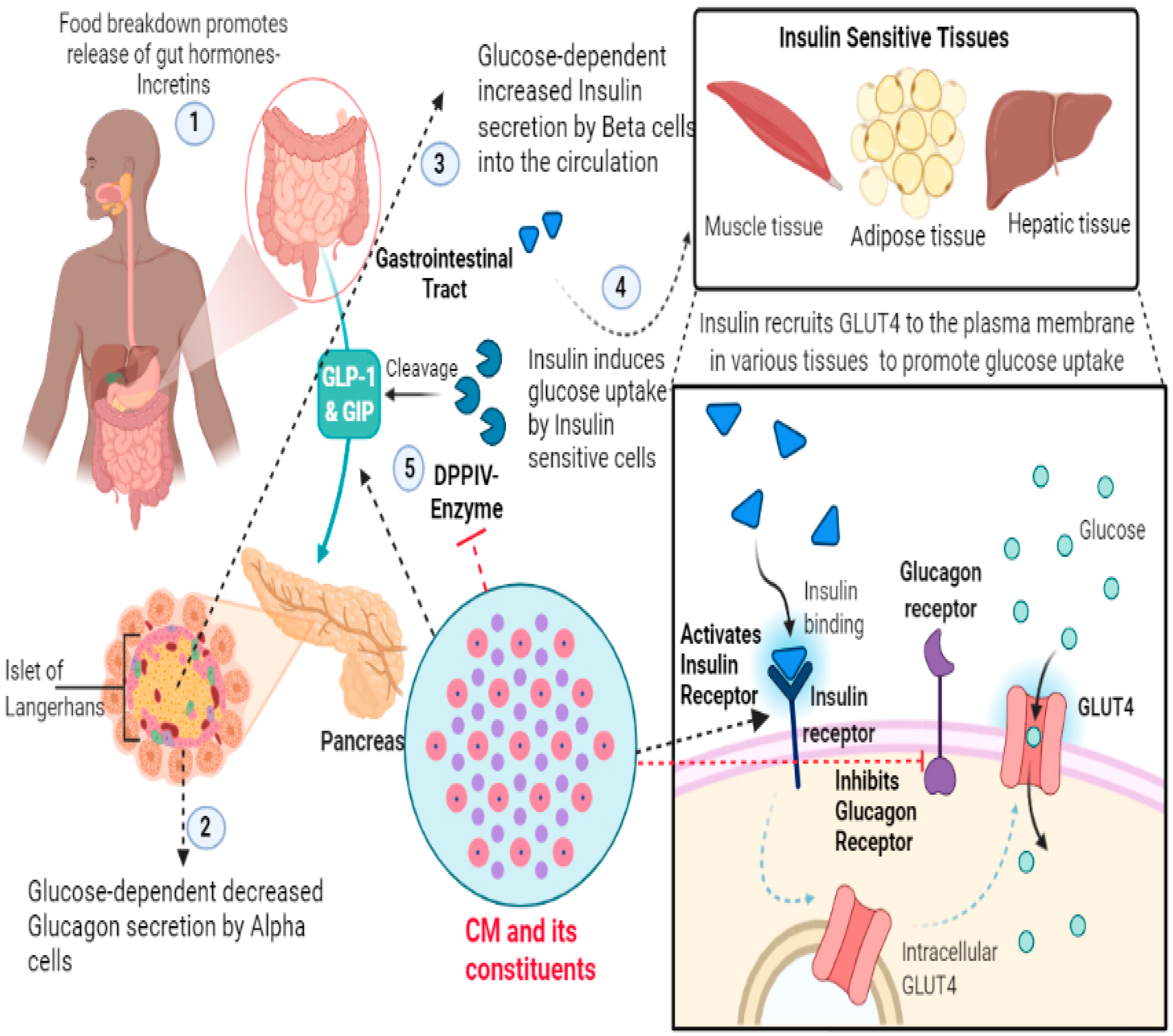
2.2.3. Molecular Intricacies of CM’s Anti-Diabetic Potential
2.2.4. Molecular Intricacies of CM’s Anti-Microbial Potential
2.3. Non-Nutritional Components (CM Exosomes)
3. Pharmacological Properties of CM Exosomes
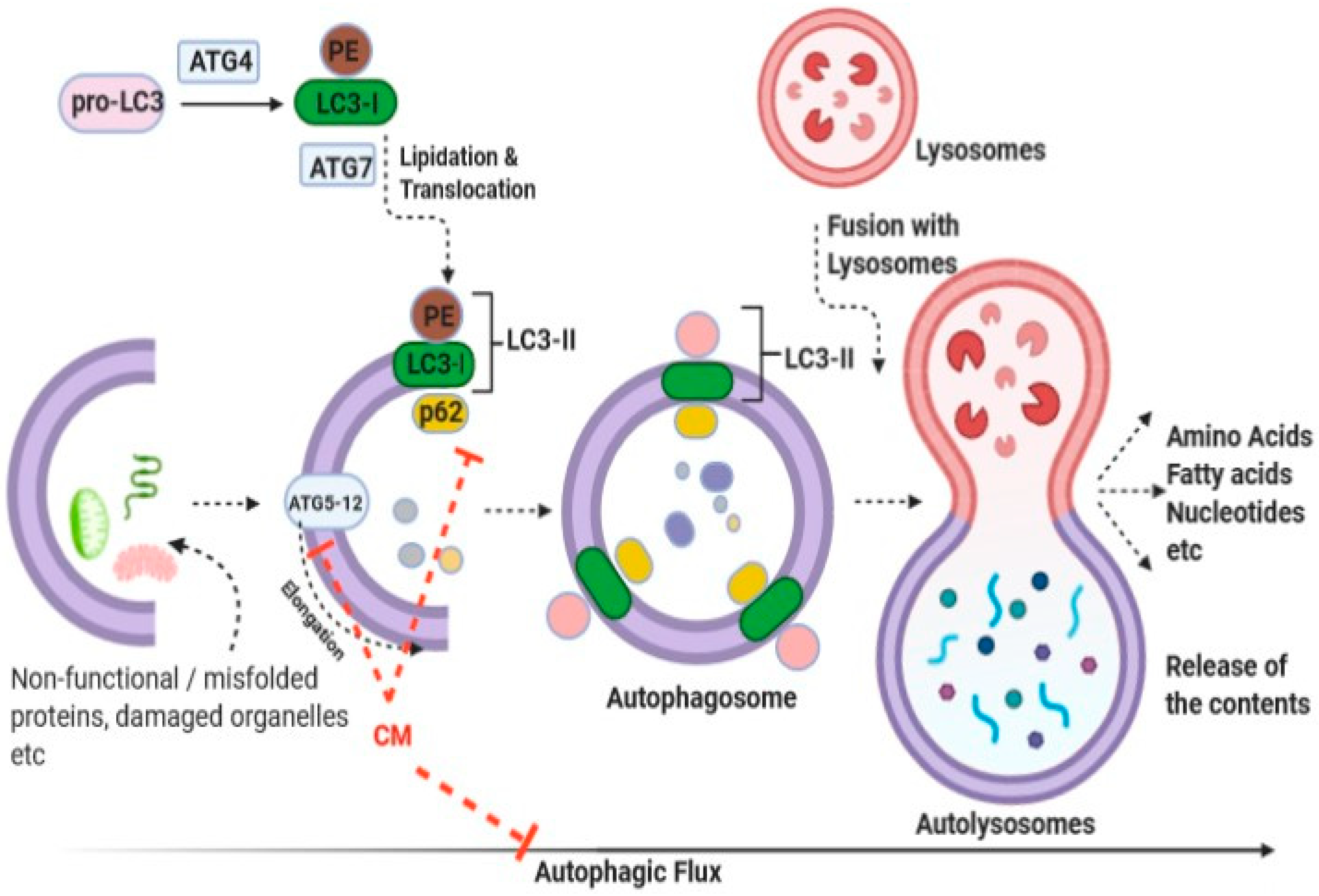
3.1. Molecular Intricacies of Their Anti-Cancer Potential
3.2. CM Exosomes as a Natural Biogenic Nano-Delivery Platform for Therapeutics
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| CM | Camel milk |
| CP | Casein protein |
| WP | whey protein |
| α-LA | α-Lactalbumin |
| CSA | camel serum al-bumin |
| LF | lactoferrin |
| ACE | Angiotensin converting Enzyme |
| Ang II | Angiotensin II |
| CMPH | CM protein hydrolysates |
| RAS | Renin Angiotensin system |
References
- Badawy, A.A.; El-Magd, M.A.; AlSadrah, S.A. Therapeutic Effect of Camel Milk and Its Exosomes on MCF7 Cells In Vitro and In Vivo. Integr. Cance Ther. 2018, 17, 1235–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, H.M.; Ibrahim, W.H.; Schneider-Stock, R.; Hassan, H.M. Camel Milk Lactoferrin Reduces the Proliferation of Colorectal Cancer Cells and Exerts Antioxidant and DNA Damage Inhibitory Activities. Food Chem. 2013, 141, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.M.; Mohammed-Geba, K.; Tawfic, A.A.; El-Magd, M.A. Camel Milk Exosomes Modulate Cyclophosphamide-Induced Oxidative Stress and Immuno-Toxicity in Rats. Food Funct. 2019, 10, 7523–7532. [Google Scholar] [CrossRef] [PubMed]
- Arain, M.A.; Khaskheli, G.B.; Shah, A.H.; Marghazani, I.B.; Barham, G.S.; Shah, Q.A.; Khand, F.M.; Buzdar, J.A.; Soomro, F.; Fazlani, S.A. Nutritional Significance and Promising Therapeutic/Medicinal Application of Camel Milk as a Functional Food in Human and Animals: A Comprehensive Review. Anim. Biotechnol. 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Anwar, I.; Khan, F.B.; Maqsood, S.; Ayoub, M.A. Camel Milk Targeting Insulin Receptor—Toward Understanding the Antidiabetic Effects of Camel Milk. Front. Nutr. 2022, 8, 819278. [Google Scholar] [CrossRef]
- Khan, F.B.; Anwar, I.; Redwan, E.M.; Palakkott, A.; Ashraf, A.; Kizhakkayil, J.; Iratni, R.; Maqsood, S.; Akli Ayoub, M. Camel and Bovine Milk Lactoferrins Activate Insulin Receptor and Its Related AKT and ERK1/2 Pathways. J. Dairy Sci. 2022, 105, 1848–1861. [Google Scholar] [CrossRef]
- Ayoub, M.A.; Palakkott, A.R.; Ashraf, A.; Iratni, R. The Molecular Basis of the Anti-Diabetic Properties of Camel Milk. Diabetes Res. Clin. Pract. 2018, 146, 305–312. [Google Scholar] [CrossRef]
- Muthukumaran, M.S.; Mudgil, P.; Baba, W.N.; Ayoub, M.A.; Maqsood, S. A Comprehensive Review on Health Benefits, Nutritional Composition and Processed Products of Camel Milk. Food Rev. Int. 2022, 1–37. [Google Scholar] [CrossRef]
- Elagamy, E.I. Effect of Heat Treatment on Camel Milk Proteins with Respect to Antimicrobial Factors: A Comparison with Cows’ and Buffalo Milk Proteins. Food Chem. 2000, 68, 227–232. [Google Scholar] [CrossRef]
- Mudgil, P.; Baby, B.; Ngoh, Y.-Y.; Kamal, H.; Vijayan, R.; Gan, C.-Y.; Maqsood, S. Molecular Binding Mechanism and Identification of Novel Anti-Hypertensive and Anti-Inflammatory Bioactive Peptides from Camel Milk Protein Hydrolysates. LWT 2019, 112, 108193. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Beniwal, R.; Kochar, D.K.; Tuteja, F.C.; Ghorui, S.K.; Sahani, M.S.; Sharma, S. Camel Milk as an Adjunct to Insulin Therapy Improves Long-Term Glycemic Control and Reduction in Doses of Insulin in Patients with Type-1 Diabetes A 1 Year Randomized Controlled Trial. Diabetes Res Clin Pract. 2005, 68, 176–177. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Al-Dhaheri, A.S.; Al Mahadin, S.; Kizhakkayil, J.; Abushelaibi, A. In Vitro Investigation of Anticancer, Antihypertensive, Antidiabetic, and Antioxidant Activities of Camel Milk Fermented with Camel Milk Probiotic: A Comparative Study with Fermented Bovine Milk. J. Dairy Sci. 2018, 101, 900–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Guo, K.; Chen, Q.; Wang, Y. Jirimutu Camel Milk Modulates the Gut Microbiota and Has Anti-Inflammatory Effects in a Mouse Model of Colitis. J. Dairy Sci. 2022, 105, 3782–3793. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.; Mudgil, P.; Palakkott, A.; Iratni, R.; Gan, C.-Y.; Maqsood, S.; Ayoub, M.A. Molecular Basis of the Anti-Diabetic Properties of Camel Milk through Profiling of Its Bioactive Peptides on Dipeptidyl Peptidase IV (DPP-IV) and Insulin Receptor Activity. J. Dairy Sci. 2021, 104, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Abdulrahman, A.O.; Ismael, M.A.; Al-Hosaini, K.; Rame, C.; Al-Senaidy, A.M.; Dupont, J.; Ayoub, M.A. Differential Effects of Camel Milk on Insulin Receptor Signaling–Toward Understanding the Insulin-Like Properties of Camel Milk. Front. Endocrinol. 2016, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, R.P.; Budania, S.; Sharma, P.; Gupta, R.; Kochar, D.K.; Panwar, R.B.; Sahani, M.S. Zero Prevalence of Diabetes in Camel Milk Consuming Raica Community of North-West Rajasthan, India. Diabetes Res. Clin. Pract. 2007, 76, 290–296. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Saran, S.; Sharma, P.; Gupta, R.P.; Kochar, D.K.; Sahani, M.S. Effect of Camel Milk on Residual Beta-Cell Function in Recent Onset Type 1 Diabetes. Diabetes Res. Clin. Pract. 2007, 77, 494–495. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Sharma, P.; Gafoorunissa, S.J.; Ibrahim, S.A.; Shah, B.; Shukla, D.K.; Kaur, T. Effect of Camel Milk on Glucose Metabolism in Adults with Normal Glucose Tolerance and Type 2 Diabetes in Raica Community: A Crossover Study. Acta Biomed. 2011, 82, 181–186. [Google Scholar]
- Abou-Soliman, N.H.I.; Awad, S.; Desouky, M.M. Effect of Digestive Enzymes on the Activity of Camel-Milk Insulin. Int. J. Dairy Technol. 2020, 73, 341–344. [Google Scholar] [CrossRef]
- Vaisman, N.; Reuven, Y.; Uzi, M.; Georgi, G.; Boehm, G. Camel’s Milk and Gastric Emptying. Clin. Nutr. 2006, 25, 622–625. [Google Scholar] [CrossRef]
- Malik, A.; Al-Senaidy, A.; Skrzypczak-Jankun, E.; Jankun, J. A Study of the Anti-Diabetic Agents of Camel Milk. Int. J. Mol. Med. 2012, 30, 585–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mati, A.; Senoussi-Ghezali, C.; Zennia, S.S.A.; Almi-Sebbane, D.; El-Hatmi, H.; Girardet, J.-M. Dromedary Camel Milk Proteins, a Source of Peptides Having Biological Activities—A Review. Int. Dairy J. 2017, 73, 25. [Google Scholar] [CrossRef]
- Ali Redha, A.; Valizadenia, H.; Siddiqui, S.A.; Maqsood, S. A State-of-Art Review on Camel Milk Proteins as an Emerging Source of Bioactive Peptides with Diverse Nutraceutical Properties. Food Chem. 2022, 373, 131444. [Google Scholar] [CrossRef] [PubMed]
- Izadi, A.; Khedmat, L.; Mojtahedi, S.Y. Nutritional and Therapeutic Perspectives of Camel Milk and Its Protein Hydrolysates: A Review on Versatile Biofunctional Properties. J. Funct. Foods 2019, 60, 103441. [Google Scholar] [CrossRef]
- El-Agamy, E.I. Bioactive Components in Camel Milk. Bioact. Compon. Milk Dairy Prod. 2009, 107, 159–192. [Google Scholar]
- Swelum, A.A.; El-Saadony, M.T.; Abdo, M.; Ombarak, R.A.; Hussein, E.O.S.; Suliman, G.; Alhimaidi, A.R.; Ammari, A.A.; Ba-Awadh, H.; Taha, A.E.; et al. Nutritional, Antimicrobial and Medicinal Properties of Camel’s Milk: A Review. Saudi J. Biol. Sci. 2021, 28, 3126–3136. [Google Scholar] [CrossRef]
- Kumar, D.; Chatli, M.K.; Singh, R.; Mehta, N.; Kumar, P. Antioxidant and Antimicrobial Activity of Camel Milk Casein Hydrolysates and Its Fractions. Small Rumin. Res. 2016, 139, 20–25. [Google Scholar] [CrossRef]
- Salami, M.; Moosavi-Movahedi, A.A.; Ehsani, M.R.; Yousefi, R.; Haertlé, T.; Chobert, J.-M.; Razavi, S.H.; Henrich, R.; Balalaie, S.; Ebadi, S.A.; et al. Improvement of the Antimicrobial and Antioxidant Activities of Camel and Bovine Whey Proteins by Limited Proteolysis. J. Agric. Food Chem. 2010, 58, 3297–3302. [Google Scholar] [CrossRef]
- Agamy, E.S.I.E.; Ruppanner, R.; Ismail, A.; Champagne, C.P.; Assaf, R. Antibacterial and Antiviral Activity of Camel Milk Protective Proteins. J. Dairy Res. 1992, 59, 169–175. [Google Scholar] [CrossRef]
- Kaskous, S. Importance of Camel Milk for Human Health. Emir. J. Food Agric. 2016, 28, 158–163. [Google Scholar] [CrossRef]
- AL-Ayadhi, L.Y.; Elamin, N.E. Camel Milk as a Potential Therapy as an Antioxidant in Autism Spectrum Disorder (ASD). Evid. Based Complement. Altern. Med. 2013, 2013, 602834. [Google Scholar] [CrossRef] [PubMed]
- Alavi, F.; Salami, M.; Emam-Djomeh, Z.; Mohammadian, M. Nutraceutical Properties of Camel Milk. In Nutrients in Dairy and Their Implications for Health and Disease; Elsevier: Amsterdam, The Netherlands, 2017; pp. 451–468. ISBN 978-0-12-809762-5. [Google Scholar]
- Maqsood, S.; Al-Dowaila, A.; Mudgil, P.; Kamal, H.; Jobe, B.; Hassan, H.M. Comparative Characterization of Protein and Lipid Fractions from Camel and Cow Milk, Their Functionality, Antioxidant and Antihypertensive Properties upon Simulated Gastro-Intestinal Digestion. Food Chem. 2019, 279, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Mal, G.; Kumar, D.; Patil, N.V.; Pathak, K.M.L. Camel Milk: An Important Natural Adjuvant. Agric. Res. 2017, 6, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-U.; Kim, W.-H.; Jeong, C.H.; Yi, D.Y.; Min, H. More than Nutrition: Therapeutic Potential of Breast Milk-Derived Exosomes in Cancer. Int. J. Mol. Sci. 2020, 21, 7327. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xiao, Y.; Orgoldol, K.; Ming, L.; Yi, L.; Ji, R. Effects of Geographic Region on the Composition of Bactrian Camel Milk in Mongolia. Animals 2019, 9, 890. [Google Scholar] [CrossRef] [Green Version]
- Konuspayeva, G.; Faye, B.; Loiseau, G. The Composition of Camel Milk: A Meta-Analysis of the Literature Data. J. Food Compos. Anal. 2009, 22, 95–101. [Google Scholar] [CrossRef]
- Al haj, O.A.; Al Kanhal, H.A. Compositional, Technological and Nutritional Aspects of Dromedary Camel Milk. Int. Dairy J. 2010, 20, 811–821. [Google Scholar] [CrossRef]
- Farah, Z.; Rettenmaier, R.; Atkins, D. Vitamin Content of Camel Milk. Int. J. Vitam. Nutr. Res. 1992, 62, 30–33. [Google Scholar]
- Khalesi, M.; Salami, M.; Moslehishad, M.; Winterburn, J.; Moosavi-Movahedi, A.A. Biomolecular Content of Camel Milk: A Traditional Superfood towards Future Healthcare Industry. Trends Food Sci. Technol. 2017, 62, 49–58. [Google Scholar] [CrossRef]
- Meena, S.; Rajput, Y.S.; Sharma, R. Comparative Fat Digestibility of Goat, Camel, Cow and Buffalo Milk. Int. Dairy J. 2014, 35, 153–156. [Google Scholar] [CrossRef]
- Shuiep, E.T.S.; Giambra, I.J.; El-Zubeir, I.E.Y.M.; Erhardt, G. Biochemical and Molecular Characterization of Polymorphisms of As1-Casein in Sudanese Camel (Camelus Dromedarius) Milk. Int. Dairy J. 2013, 28, 88–93. [Google Scholar] [CrossRef]
- Hailu, Y.; Hansen, E.B.; Seifu, E.; Eshetu, M.; Ipsen, R.; Kappeler, S. Functional and Technological Properties of Camel Milk Proteins: A Review. J. Dairy Res. 2016, 83, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Khaskheli, M.; Arain, M.A.; Chaudhry, S.; Soomro, A.H.; Qureshi, T.A. Physico-Chemical Quality of Camel Milk. J. Agric. Soc. Sci. 2005, 2, 164–166. [Google Scholar]
- Mehaia, M.A.; Hablas, M.A.; Abdel-Rahman, K.M.; El-Mougy, S.A. Milk Composition of Majaheim, Wadah and Hamra Camels in Saudi Arabia. Food Chem. 1995, 52, 115–122. [Google Scholar] [CrossRef]
- Omar, A.; Harbourne, N.; Oruna-Concha, M.J. Quantification of Major Camel Milk Proteins by Capillary Electrophoresis. Int. Dairy J. 2016, 58, 31–35. [Google Scholar] [CrossRef]
- Felfoul, I.; Jardin, J.; Gaucheron, F.; Attia, H.; Ayadi, M.A. Proteomic Profiling of Camel and Cow Milk Proteins under Heat Treatment. Food Chem. 2017, 216, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Kappeler, S.R.; Heuberger, C.; Farah, Z.; Puhan, Z. Expression of the Peptidoglycan Recognition Protein, PGRP, in the Lactating Mammary Gland. J. Dairy Sci. 2004, 87, 2660–2668. [Google Scholar] [CrossRef] [Green Version]
- Stahl, T.; Sallmann, H.-P.; Duehlmeier, R.; Wernery, U. Selected Vitamins and Fatty Acid Patterns in Dromedary Milk and Colostrum. J. Camel Pract. Res. 2006, 13, 53–57. [Google Scholar]
- Alhaj, O.A.; Altooq, N.J.; Alenezi, A.F.; Janahi, A.I.; Janahi, M.I.; Humood, A.M.; AlRasheed, M.M.; Bragazzi, N.L.; Jahrami, H.A.; Faye, B. Camel Milk Composition by Breed, Season, Publication Year, and Country: A Global Systematic Review, Meta-Analysis, and Meta-Regression. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2520–2559. [Google Scholar] [CrossRef]
- Yaqoob, M.; Nawaz, H. Potential of Pakistani Camel for Dairy and Other Uses. Anim. Sci. J. 2007, 78, 467–475. [Google Scholar] [CrossRef]
- Abrhaley, A.; Leta, S. Medicinal Value of Camel Milk and Meat. J. Appl. Anim. Res. 2018, 46, 552–558. [Google Scholar] [CrossRef] [Green Version]
- Kilari, B.P.; Mudgil, P.; Azimullah, S.; Bansal, N.; Ojha, S.; Maqsood, S. Effect of Camel Milk Protein Hydrolysates against Hyperglycemia, Hyperlipidemia, and Associated Oxidative Stress in Streptozotocin (STZ)-Induced Diabetic Rats. J. Dairy Sci. 2021, 104, 1304–1317. [Google Scholar] [CrossRef] [PubMed]
- Baba, W.N.; Mudgil, P.; Kamal, H.; Kilari, B.P.; Gan, C.-Y.; Maqsood, S. Identification and Characterization of Novel α-Amylase and α-Glucosidase Inhibitory Peptides from Camel Whey Proteins. J. Dairy Sci. 2021, 104, 1364–1377. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Sharma, H.; Deshwal, G.K.; Rao, P.S. A Comprehensive Review on Bioactive Peptides Derived from Milk and Milk Products of Minor Dairy Species. Food Prod. Process. Nutr. 2021, 3, 2. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Isono, H.; Miyata, T. Potential Antioxidant Bioactive Peptides from Camel Milk Proteins. Anim. Nutr. 2018, 4, 273–280. [Google Scholar] [CrossRef]
- Chiang, J.-T.; Badrealam, K.F.; Shibu, M.A.; Cheng, S.-F.; Shen, C.-Y.; Chang, C.-F.; Lin, Y.-M.; Viswanadha, V.P.; Liao, S.-C.; Huang, C.-Y. Anti-Apoptosis and Anti-Fibrosis Effects of Eriobotrya Japonica in Spontaneously Hypertensive Rat Hearts. Int. J. Mol. Sci. 2018, 19, 1638. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.-T.; Badrealam, K.F.; Shibu, M.A.; Kuo, C.-H.; Huang, C.-Y.; Chen, B.-C.; Lin, Y.-M.; Viswanadha, V.P.; Kuo, W.-W.; Huang, C.-Y. Eriobotrya Japonica Ameliorates Cardiac Hypertrophy in H9c2 Cardiomyoblast and in Spontaneously Hypertensive Rats. Environ. Toxicol. 2018, 33, 1113–1122. [Google Scholar] [CrossRef]
- Xiong, X. Integrating Traditional Chinese Medicine into Western Cardiovascular Medicine: An Evidence-Based Approach. Nat. Rev. Cardiol. 2015, 12, 374. [Google Scholar] [CrossRef] [Green Version]
- Zibaee, S.; Yousefi, M.; Taghipour, A.; Kiani, M.A.; Noras, M.R. Nutritional and Therapeutic Characteristics of Camel Milk in Children: A Systematic Review. Electron. Physician 2015, 7, 1523–1528. [Google Scholar] [CrossRef] [Green Version]
- Al-Shamsi, K.A.; Mudgil, P.; Hassan, H.M.; Maqsood, S. Camel Milk Protein Hydrolysates with Improved Technofunctional Properties and Enhanced Antioxidant Potential in in Vitro and in Food Model Systems. J. Dairy Sci. 2018, 101, 47–60. [Google Scholar] [CrossRef]
- Kamal, H.; Jafar, S.; Mudgil, P.; Murali, C.; Amin, A.; Maqsood, S. Inhibitory Properties of Camel Whey Protein Hydrolysates toward Liver Cancer Cells, Dipeptidyl Peptidase-IV, and Inflammation. J. Dairy Sci. 2018, 101, 8711–8720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudgil, P.; Kamal, H.; Yuen, G.C.; Maqsood, S. Characterization and Identification of Novel Antidiabetic and Anti-Obesity Peptides from Camel Milk Protein Hydrolysates. Food Chem. 2018, 259, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Cadamuro, C.; Le Gouic, A.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Properties of a Camel Whey Protein Enriched Hydrolysate Preparation. Food Chem. 2019, 279, 70–79. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Paolella, S.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Properties of Camel Milk Protein Hydrolysates Generated with Trypsin. J. Funct. Foods 2017, 34, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Yassin, A.M.; Hamid, M.I.A.; Farid, O.A.; Amer, H.; Warda, M. Dromedary Milk Exosomes as Mammary Transcriptome Nano-Vehicle: Their Isolation, Vesicular and Phospholipidomic Characterizations. J. Adv. Res. 2016, 7, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Mudgil, P.; Maqsood, S. Camel Whey Protein Microparticles for Safe and Efficient Delivery of Novel Camel Milk Derived Probiotics. LWT 2019, 108, 81–88. [Google Scholar] [CrossRef]
- Yu, M.; Wu, J.; Shi, J.; Farokhzad, O.C. Nanotechnology for Protein Delivery: Overview and Perspectives. J. Control. Release 2016, 240, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Korashy, H.M.; Maayah, Z.H.; Abd-Allah, A.R.; El-Kadi, A.O.S.; Alhaider, A.A. Camel Milk Triggers Apoptotic Signaling Pathways in Human Hepatoma HepG2 and Breast Cancer MCF7 Cell Lines through Transcriptional Mechanism. J. Biomed. Biotechnol. 2012, 2012, 593195. [Google Scholar] [CrossRef] [PubMed]
- Korashy, H.M.; El Gendy, M.A.M.; Alhaider, A.A.; El-Kadi, A.O. Camel Milk Modulates the Expression of Aryl Hydrocarbon Receptor-Regulated Genes, Cyp1a1, Nqo1, and Gsta1, in Murine Hepatoma Hepa 1c1c7 Cells. J. Biomed. Biotechnol. 2012, 2012, e782642. [Google Scholar] [CrossRef] [Green Version]
- Krishnankutty, R.; Iskandarani, A.; Therachiyil, L.; Uddin, S.; Azizi, F.; Kulinski, M.; Bhat, A.A.; Mohammad, R.M. Anticancer Activity of Camel Milk via Induction of Autophagic Death in Human Colorectal and Breast Cancer Cells. Asian Pac. J. Cancer Prev. 2018, 19, 3501–3509. [Google Scholar] [CrossRef] [Green Version]
- Hasson, S.S.A.A.; Al-Busaidi, J.Z.; Al-Qarni, Z.A.M.; Rajapakse, S.; Al-Bahlani, S.; Idris, M.A.; Sallam, T.A. In Vitro Apoptosis Triggering in the BT-474 Human Breast Cancer Cell Line by Lyophilised Camel’s Milk. Asian Pac. J. Cancer Prev. 2015, 16, 6651–6661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayyash, M.; Al-Nuaimi, A.K.; Al-Mahadin, S.; Liu, S.-Q. In Vitro Investigation of Anticancer and ACE-Inhibiting Activity, α-Amylase and α-Glucosidase Inhibition, and Antioxidant Activity of Camel Milk Fermented with Camel Milk Probiotic: A Comparative Study with Fermented Bovine Milk. Food Chem. 2018, 239, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Shariatikia, M.; Behbahani, M.; Mohabatkar, H. Anticancer Activity of Cow, Sheep, Goat, Mare, Donkey and Camel Milks and Their Caseins and Whey Proteins and in Silico Comparison of the Caseins. Mol. Biol. Res. Commun. 2017, 6, 57–64. [Google Scholar] [PubMed]
- Abdallah, L.A.; Sawafta, A.M.; Ali, S.A.B.; Baradia, H.A. Cytotoxic Potential of Camel Whey and Milk on Cervix Cancer (HeLa) Cell Line. Asian J. Med. Biol. Res. 2019, 5, 231–236. [Google Scholar] [CrossRef]
- Yang, J.; Dou, Z.; Peng, X.; Wang, H.; Shen, T.; Liu, J.; Li, G.; Gao, Y. Transcriptomics and Proteomics Analyses of Anti-Cancer Mechanisms of TR35-An Active Fraction from Xinjiang Bactrian Camel Milk in Esophageal Carcinoma Cell. Clin. Nutr. 2019, 38, 2349–2359. [Google Scholar] [CrossRef]
- El Miniawy, H.M.F.; Ahmed, K.A.; Mansour, S.A.; Khattab, M.M.S. In Vivo Antitumour Potential of Camel’s Milk against Hepatocellular Carcinoma in Rats and Its Improvement of Cisplatin Renal Side Effects. Pharm. Biol. 2017, 55, 1513–1520. [Google Scholar] [CrossRef] [Green Version]
- Al-Omari, M.M.; Razan, B.; Al-Ghariebeh, A.A.A.A.; Zoubi, H.A.-; Al-Qaoud, K.M. Camel Milk Whey Inhibits Inflammatory Colorectal Cancer Development Via Down Regulation of Pro-Inflammatory Cytokines in Induced AOM/DSS Mouse Model. Emir. J. Food Agric. 2019, 31, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Alhaider, A.A.; Abdel Gader, A.G.M.; Almeshaal, N.; Saraswati, S. Camel Milk Inhibits Inflammatory Angiogenesis via Downregulation of Proangiogenic and Proinflammatory Cytokines in Mice. APMIS 2014, 122, 599–607. [Google Scholar] [CrossRef]
- El-kattawy, A.M.; Algezawy, O.; Alfaifi, M.Y.; Noseer, E.A.; Hawsawi, Y.M.; Alzahrani, O.R.; Algarni, A.; Kahilo, K.A.; El-Magd, M.A. Therapeutic Potential of Camel Milk Exosomes against HepaRG Cells with Potent Apoptotic, Anti-Inflammatory, and Anti-Angiogenesis Effects for Colostrum Exosomes. Biomed. Pharmacother. 2021, 143, 112220. [Google Scholar] [CrossRef]
- Rahimi, M.; Ghaffari, S.M.; Salami, M.; Mousavy, S.J.; Niasari-Naslaji, A.; Jahanbani, R.; Yousefinejad, S.; Khalesi, M.; Moosavi-Movahedi, A.A. ACE-Inhibitory and Radical Scavenging Activities of Bioactive Peptides Obtained from Camel Milk Casein Hydrolysis with Proteinase K. Dairy Sci. Technol. 2016, 96, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Yahya, M.A.; Alhaj, O.A.; Al-Khalifah, A.S. Antihypertensive Effect of Fermented Skim Camel (Camelus Dromedarius) Milk on Spontaneously Hypertensive Rats. Nutr. Hosp. 2017, 34, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Jrad, Z.; Hatmi, H.E.; Adt, I.; Khorchani, T.; Degraeve, P.; Oulahal, N. Antimicrobial Activity of Camel Milk Casein and Its Hydrolysates. Acta Aliment. 2015, 44, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Mudgil, P.; AlMazroui, M.; Redha, A.A.; Kilari, B.P.; Srikumar, S.; Maqsood, S. Cow and Camel Milk-Derived Whey and Casein Protein Hydrolysates Demonstrated Effective Antifungal Properties against Selected Candida Species. J. Dairy Sci. 2022, 105, 1878–1888. [Google Scholar] [CrossRef]
- Quan, S.; Tsuda, H.; Miyamoto, T. Angiotensin I-Converting Enzyme Inhibitory Peptides in Skim Milk Fermented with Lactobacillus Helveticus 130B4 from Camel Milk in Inner Mongolia, China. J. Sci. Food Agric. 2008, 88, 2688–2692. [Google Scholar] [CrossRef]
- Jrad, Z.; El Hatmi, H.; Adt, I.; Girardet, J.-M.; Cakir-Kiefer, C.; Jardin, J.; Degraeve, P.; Khorchani, T.; Oulahal, N. Effect of Digestive Enzymes on Antimicrobial, Radical Scavenging and Angiotensin I-Converting Enzyme Inhibitory Activities of Camel Colostrum and Milk Proteins. Dairy Sci. Technol. 2014, 94, 205–224. [Google Scholar] [CrossRef]
- Alhaj, O.A. Identification of Potential ACE-Inhibitory Peptides from Dromedary Fermented Camel Milk. CyTA-J. Food 2017, 15, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Solanki, D.; Hati, S.; Sakure, A. In Silico and In Vitro Analysis of Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Bioactive Peptides Derived from Fermented Camel Milk (Camelus Dromedarius). Int. J. Pept. Res. Ther. 2017, 23, 441–459. [Google Scholar] [CrossRef]
- Soleymanzadeh, N.; Mirdamadi, S.; Kianirad, M. Antioxidant Activity of Camel and Bovine Milk Fermented by Lactic Acid Bacteria Isolated from Traditional Fermented Camel Milk (Chal). Dairy Sci. Technol. 2016, 96, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Paolella, S.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Identification of Novel Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides in Camel Milk Protein Hydrolysates. Food Chem. 2018, 244, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I.; et al. Molecular Definitions of Autophagy and Related Processes. EMBO J. 2017, 36, 1811–1836. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-M.; Badrealam, K.F.; Kuo, W.-W.; Lai, P.F.; Shao-Tsu Chen, W.; Hsuan Day, C.; Ho, T.-J.; Viswanadha, V.P.; Shibu, M.A.; Huang, C.-Y. Nerolidol Improves Cardiac Function in Spontaneously Hypertensive Rats by Inhibiting Cardiac Inflammation and Remodelling Associated TLR4/ NF-ΚB Signalling Cascade. Food Chem. Toxicol. 2021, 147, 111837. [Google Scholar] [CrossRef]
- Garbers, D.L.; Dubois, S.K. The Molecular Basis of Hypertension. Annu. Rev. Biochem. 1999, 68, 127–155. [Google Scholar] [CrossRef]
- Te Riet, L.; van Esch, J.H.M.; Roks, A.J.M.; van den Meiracker, A.H.; Danser, A.H.J. Hypertension: Renin-Angiotensin-Aldosterone System Alterations. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef]
- Mehta, P.K.; Griendling, K.K. Angiotensin II Cell Signaling: Physiological and Pathological Effects in the Cardiovascular System. Am. J. Physiol.-Cell Physiol. 2007, 292, C82–C97. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Shamsia, S.; Conte, A. Release of Angiotensin Converting Enzyme-Inhibitory Peptides during in Vitro Gastro-Intestinal Digestion of Camel Milk. Int. Dairy J. 2016, 56, 119–128. [Google Scholar] [CrossRef]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef]
- Ogihara, T.; Higashimori, K.; Masuo, K.; Mikami, H. Pilot Study of a New Angiotensin II Receptor Antagonist, TCV-116: Effects of a Single Oral Dose on Blood Pressure in Patients with Essential Hypertension. Clin. Ther. 1993, 15, 684–691. [Google Scholar] [PubMed]
- Ji, Y.; Liu, J.; Wang, Z.; Liu, N. Angiotensin II Induces Inflammatory Response Partly via Toll-like Receptor 4-Dependent Signaling Pathway in Vascular Smooth Muscle Cells. Cell Physiol. Biochem. 2009, 23, 265–276. [Google Scholar] [CrossRef]
- Kang, P.M.; Landau, A.J.; Eberhardt, R.T.; Frishman, W.H. Angiotensin II Receptor Antagonists: A New Approach to Blockade of the Renin-Angiotensin System. Am. Heart J. 1994, 127, 1388–1401. [Google Scholar] [CrossRef]
- Timmermans, P.B.; Wong, P.C.; Chiu, A.T.; Herblin, W.F.; Benfield, P.; Carini, D.J.; Lee, R.J.; Wexler, R.R.; Saye, J.A.; Smith, R.D. Angiotensin II Receptors and Angiotensin II Receptor Antagonists. Pharm. Rev. 1993, 45, 205–251. [Google Scholar] [PubMed]
- Matsubara, H. Pathophysiological Role of Angiotensin II Type 2 Receptor in Cardiovascular and Renal Diseases. Circ. Res. 1998, 83, 1182–1191. [Google Scholar] [CrossRef]
- Carey, R.M.; Wang, Z.Q.; Siragy, H.M. Role of the Angiotensin Type 2 Receptor in the Regulation of Blood Pressure and Renal Function. Hypertension 2000, 35, 155–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, Y.; Ozono, R.; Yano, Y.; Teranishi, Y.; Akishita, M.; Horiuchi, M.; Oshima, T.; Kambe, M. Cardioprotective Role of AT2 Receptor in Postinfarction Left Ventricular Remodeling. Hypertension 2003, 41, 814–818. [Google Scholar] [CrossRef] [Green Version]
- Tsutsumi, Y.; Matsubara, H.; Ohkubo, N.; Mori, Y.; Nozawa, Y.; Murasawa, S.; Kijima, K.; Maruyama, K.; Masaki, H.; Moriguchi, Y.; et al. Angiotensin II Type 2 Receptor Is Upregulated in Human Heart With Interstitial Fibrosis, and Cardiac Fibroblasts Are the Major Cell Type for Its Expression. Circ. Res. 1998, 83, 1035–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallianou, N.G.; Geladari, E.V.; Kounatidis, D.; Geladari, C.V.; Stratigou, T.; Dourakis, S.P.; Andreadis, E.A.; Dalamaga, M. Diabetes Mellitus in the Era of Climate Change. Diabetes Metab. 2021, 47, 101205. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and Societal Implications of the Diabetes Epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- AlKurd, R.; Hanash, N.; Khalid, N.; Abdelrahim, D.N.; Khan, M.A.B.; Mahrous, L.; Radwan, H.; Naja, F.; Madkour, M.; Obaideen, K.; et al. Effect of Camel Milk on Glucose Homeostasis in Patients with Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2022, 14, 1245. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.P.; Dogra, R.; Mohta, N.; Tiwari, R.; Singhal, S.; Sultania, S. Beneficial Effect of Camel Milk in Diabetic Nephropathy. Acta Biomed. 2009, 80, 131–134. [Google Scholar]
- Contreras-Ferrat, A.; Lavandero, S.; Jaimovich, E.; Klip, A. Calcium Signaling in Insulin Action on Striated Muscle. Cell Calcium 2014, 56, 390–396. [Google Scholar] [CrossRef]
- Muoio, D.M.; Newgard, C.B. Mechanisms of Disease:Molecular and Metabolic Mechanisms of Insulin Resistance and Beta-Cell Failure in Type 2 Diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Galali, Y.; Galali, Y. Miraculous Properties of Camel Milk and Perspective of Modern Science. J. Fam. Med. Dis. Prev. 2019, 5, 1–7. [Google Scholar] [CrossRef]
- Mudgil, P.; Baba, W.N.; Kamal, H.; FitzGerald, R.J.; Hassan, H.M.; Ayoub, M.A.; Gan, C.-Y.; Maqsood, S. A Comparative Investigation into Novel Cholesterol Esterase and Pancreatic Lipase Inhibitory Peptides from Cow and Camel Casein Hydrolysates Generated upon Enzymatic Hydrolysis and In-Vitro Digestion. Food Chem. 2022, 367, 130661. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.P.; Jain, S.; Shah, S.; Chopra, A.; Agarwal, V. Effect of Camel Milk on Glycemic Control and Insulin Requirement in Patients with Type 1 Diabetes: 2-Years Randomized Controlled Trial. Eur. J. Clin. Nutr. 2011, 65, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Ejtahed, H.S.; Niasari Naslaji, A.; Mirmiran, P.; Zraif Yeganeh, M.; Hedayati, M.; Azizi, F.; Moosavi Movahedi, A. Effect of Camel Milk on Blood Sugar and Lipid Profile of Patients with Type 2 Diabetes: A Pilot Clinical Trial. Int. J. Endocrinol. Metab. 2015, 13, e21160. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.; Wattoo, F.H.; Wattoo, M.H.S.; Gulfraz, M.; Masud, T.; Shah, I.; Ali, S.; Alavi, S.E. Camel Milk as an Alternative Treatment Regimen for Diabetes Therapy. Food Sci. Nutr. 2021, 9, 1347–1356. [Google Scholar] [CrossRef]
- Mirmiran, P.; Ejtahed, H.-S.; Angoorani, P.; Eslami, F.; Azizi, F. Camel Milk Has Beneficial Effects on Diabetes Mellitus: A Systematic Review. Int. J. Endocrinol. Metab. 2017, 15, e42150. [Google Scholar] [CrossRef] [Green Version]
- Shori, A.B. Camel Milk as a Potential Therapy for Controlling Diabetes and Its Complications: A Review of In Vivo Studies. J. Food Drug Anal. 2015, 23, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Quijia, C.R.; Alves, R.C.; Hanck-Silva, G.; Galvão Frem, R.C.; Arroyos, G.; Chorilli, M. Metal-Organic Frameworks for Diagnosis and Therapy of Infectious Diseases. Crit. Rev. Microbiol. 2022, 48, 161–196. [Google Scholar] [CrossRef]
- El-Fakharany, E.M.; El-Baky, N.A.; Linjawi, M.H.; Aljaddawi, A.A.; Saleem, T.H.; Nassar, A.Y.; Osman, A.; Redwan, E.M. Influence of Camel Milk on the Hepatitis C Virus Burden of Infected Patients. Exp. Ther. Med. 2017, 13, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Behrouz, S.; Saadat, S.; Memarzia, A.; Sarir, H.; Folkerts, G.; Boskabady, M.H. The Antioxidant, Anti-Inflammatory and Immunomodulatory Effects of Camel Milk. Front. Immunol. 2022, 13, 855342. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xi, Q.-Y.; Ye, R.-S.; Cheng, X.; Qi, Q.-E.; Wang, S.-B.; Shu, G.; Wang, L.-N.; Zhu, X.-T.; Jiang, Q.-Y.; et al. Exploration of MicroRNAs in Porcine Milk Exosomes. BMC Genom. 2014, 15, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yanoshita, R. Exosome-like Vesicles with Dipeptidyl Peptidase IV in Human Saliva. Biol. Pharm. Bull. 2008, 31, 1059–1062. [Google Scholar] [CrossRef] [Green Version]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filén, J.-J.; Lahesmaa, R.; Norman, M.; Neve, E.P.A.; Scheynius, A.; Gabrielsson, S. Exosomes with Immune Modulatory Features Are Present in Human Breast Milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef]
- Hata, T.; Murakami, K.; Nakatani, H.; Yamamoto, Y.; Matsuda, T.; Aoki, N. Isolation of Bovine Milk-Derived Microvesicles Carrying MRNAs and MicroRNAs. Biochem. Biophys. Res. Commun. 2010, 396, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Huang-Doran, I.; Zhang, C.-Y.; Vidal-Puig, A. Extracellular Vesicles: Novel Mediators of Cell Communication In Metabolic Disease. Trends Endocrinol. Metab. 2017, 28, 3–18. [Google Scholar] [CrossRef]
- Zempleni, J.; Aguilar-Lozano, A.; Sadri, M.; Sukreet, S.; Manca, S.; Wu, D.; Zhou, F.; Mutai, E. Biological Activities of Extracellular Vesicles and Their Cargos from Bovine and Human Milk in Humans and Implications for Infants. J. Nutr. 2017, 147, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Reif, S.; Elbaum Shiff, Y.; Golan-Gerstl, R. Milk-Derived Exosomes (MDEs) Have a Different Biological Effect on Normal Fetal Colon Epithelial Cells Compared to Colon Tumor Cells in a MiRNA-Dependent Manner. J. Transl. Med. 2019, 17, 325. [Google Scholar] [CrossRef]
- Agrawal, A.K.; Aqil, F.; Jeyabalan, J.; Spencer, W.A.; Beck, J.; Gachuki, B.W.; Alhakeem, S.S.; Oben, K.; Munagala, R.; Bondada, S.; et al. Milk-Derived Exosomes for Oral Delivery of Paclitaxel. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.K.; Gupta, R. Exosomes for the Enhanced Tissue Bioavailability and Efficacy of Curcumin. AAPS J. 2017, 19, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.K.; Kyakulaga, A.-H.; Wilcher, S.A.; Gupta, R.C. Milk Exosomes-Natural Nanoparticles for SiRNA Delivery. Cancer Lett. 2019, 449, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; Aqil, F.; Tyagi, N.; Gupta, R. Milk Exosomes: A Biogenic Nanocarrier for Small Molecules and Macromolecules to Combat Cancer. Am. J. Reprod. Immunol. 2021, 85, e13349. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine Milk-Derived Exosomes for Drug Delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Camel Milk Constituents | Cell Line/Animal Model/Assay Used | Mechanism | Reference |
|---|---|---|---|---|
| Cancer | ||||
| Nutritional Components | ||||
| 1 | Camel Milk | Human Hepatoma Cell Line (Hep-G2) and Human Breast Cancer Cell Line (MCF-7) | Induction of Apoptosis | [69] |
| 2 | Camel Milk | Murine Hepatoma Hepa 1c1c7 Cell Line | Modulation of the expression of cancer-related genes (Cyp1a1), (Nqo1), and (Gsta1) at the transcriptional and post-transcriptional levels | [70] |
| 3 | Camel Milk | Human Colorectal Cancer Cell line (HCT-116) and Breast Cancer Cell Line (MCF-7) | Induction of Autophagic Cell Death | [71] |
| 4 | Lyophilised Camel Milk | Human Breast Cancer Cell Line (BT-474) | Induction of Apoptosis | [72] |
| 5 | Camel Milk Lactoferrin | Human Colon Cancer Cell Line (HCT-116) | Inhibition of Human colorectal cancer cell line (HCT-116) proliferation and DNA damage inhibitory activities | [2] |
| 6 | Camel Milk fermented with Camel Milk probiotic strain Lactococcus lactis KX881782 (Lc.K782) and control Lactobacillus acidophilus DSM9126 (La.DSM) | Human Colorectal Adenocarcinoma Cell Line (Caco-2), Human Breast Cancer Cell Line (MCF-7), and and Human Adenocarcinoma Cell Line (HELA) | Inhibition of proliferation | [12] |
| 7 | Camel Milk fermented with Camel Milk probiotic strains Lb. reuteri-KX881777, Lb. plantarum-KX881772, Lb. plantarum-KX881779 and a control strain Lb. plantarum DSM2468 | Human Colorectal Adenocarcinoma Cell Line (Caco-2), Human Breast Cancer Cell Line (MCF-7) and Human Adenocarcinoma Cell Line (HELA) | Inhibition of proliferation | [73] |
| 8 | Camel Milk, Casein and Whey Proteins | Human Breast Cancer Cell Line (MCF-7) | Inhibition of proliferation as evident through MTT assay | [74] |
| 9 | Camel Milk and Whey proteins | Human Adenocarcinoma Cell Line (HELA) | Inhibition of proliferation as evident through MTT assay | [75] |
| 10 | TR35-An active fraction from Xinjiang Bactrian Camel Whey | In vitro Human Esophageal Carcinoma Cell Line (Eca-109) In vivo BALB/c nude mice subcutaneously injected with 2 × 106 Eca-109 cells | Inhibition of Eca-109 cell proliferation and induction of apoptosis | [76] |
| 11 | Camel Milk | Induced diethylnitrosamine and phenobarbitone Hepatic Cancer Wistar Rat Model | Potent Inhibitory effect on hepatocarcinogenesis in Wistar Rats was observed | [77] |
| 12 | Camel Milk Whey Protein | Induced Azoxymethane (AOM)/Dextran sodium sulfate (DSS) Mouse Model | Inhibition of inflammatory colorectal cancer development via down-regulation of pro-inflammatory cytokines | [78] |
| 13 | Camel Milk | Sponge implant angiogenesis Male Swiss Albino Mice Model | Inhibition of inflammatory angiogenesis via down-regulation of pro-angiogenic and pro-inflammatory cytokines | [79] |
| Non-Nutritional Components | ||||
| 14 | Camel Milk Exosomes | Human Colorectal Cancer Cell Line(HCT-116) and Human Breast Cancer Cell Line (MCF-7) | Induction of Autophagy | [1] |
| 15 | Camel Milk Exosomes | Albino Rat Model | Mitigation of oxidative stress and immune-toxic responses induced by the chemotherapeutic drug viz. cyclophosphamide (CTX) | [3] |
| 16 | Camel Milk Exosomes | HepaRG cells | Potential apoptotic, anti-inflammatory, and anti-angiogenesis effects against HepaRG cells | [80] |
| Hypertension | ||||
| 1 | Camel Milk Protein and Lipid fractions | Colorimetry based analytical technique | Inhibition of Angiotensin-1 converting enzyme (ACE) | [33] |
| 2 | Bioactive Peptides from Camel Milk Protein Hydrolysates | Colorimetry based analytical technique | Inhibition of Angiotensin-1 converting enzyme (ACE) and anti-inflammatory responses | [10] |
| 3 | Bioactive Peptides from Camel Milk Casein Hydrolysates | Colorimetry based analytical technique | Inhibition of Angiotensin-1 converting enzyme (ACE) and radical scavenging activities | [81] |
| 4 | Camel Milk fermented with Camel Milk probiotic strain Lactococcus lactis KX881782 (Lc.K782) and control Lactobacillus acidophilus DSM9126 (La.DSM) | Colorimetry based analytical technique | Inhibition of Angiotensin-1 converting enzyme (ACE) | [12] |
| 5 | Camel Milk fermented with Camel Milk probiotic strains Lb. reuteri-KX881777, Lb. plantarum-KX881772, Lb. plantarum-KX881779 and a control strain Lb. plantarum DSM2468 | Colorimetry based analytical technique | Inhibition of Angiotensin-1 converting enzyme (ACE) | [73] |
| 6 | Fermented Skim Camel Milk | Spontaneously Hypertensive Rats | Attenuation of systolic and diastolic blood pressure, Inhibition of Angiotensin-1 converting enzyme (ACE) | [82] |
| Diabetes | ||||
| 1 | Camel Milk and Protein Fractions | Human Embryonic Kidney Cell Line (HEK-293) | Allosteric effect on insulin receptor conformation and activation; and modulation of downstream signalling | [15] |
| 2 | Camel Milk Whey Protein and Camel Milk Whey Protein Hydrolysates | Human Liver Cancer Cell Line (Hep-G2) and Human Embryonic Kidney Cell Line (HEK-293) | Inhibition of Dipeptidyl peptidase-IV (DPP-IV), Activation of insulin receptor and Positive Regulation on Glucose Uptake | [14] |
| 3 | Camel Milk Protein Hydrolysates | Colorimetry based analytical technique | Inhibition of α-amylase | [63] |
| 4 | Camel Milk Whey Protein Hydrolysates | Colorimetry based analytical technique | Inhibition of α-amylase and α-glucosidase | [54] |
| 5 | Camel Whey Protein Hydrolysates | Colorimetry based analytical technique | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) and inflammation | [62] |
| 6 | Camel Milk Protein Hydrolysates | Colorimetry based analytical technique | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [65] |
| 7 | Camel Milk Protein Hydrolysates | Colorimetry based analytical technique | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [64] |
| 8 | Camel Whey Protein Enriched Hydrolysates | Colorimetry based analytical technique | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [64] |
| 9 | Camel Milk Protein Hydrolysates | Streptozotocin (STZ)-induced Diabetic Rats | Potent Hypoglycemic activity, as evident by reduction in fasting Blood Glucose and Oral glucose tolerance test (OGTT) levels; Preservation of β-cells was also observed | [53] |
| 10 | Camel Milk Protein Lactoferrin | HEK-293 and Hep-G2 cells | Modulation of Insulin Receptor and downstream signalling | [6] |
| Anti-microbial | ||||
| 1 | Camel Milk Casein Protein hydrolysates and its fraction | Gram positive bacteria: Staphylococcus aureus, Bacillus cereus and Listeria monocytogenes Gram negative bacteria: Escherichia coli | Significant anti-microbial activity was observed against all the microbial strain tested for all the fractions | [27] |
| 2 | Camel Whey Proteins and hydrolysates | Gram negative bacteria: Escherichia coli Dh1α | Improved anti-microbial activities of Camel Whey Proteins were observed, particularly for limited Proteolysed fractions | [28] |
| 3 | Lysozyme(LZ), lactoferrin(LF), lactoperoxidase(LP), immunoglobulin G (IgG) and secretory immunoglobulin A(Ig A) extracted from camel milk | Gram positive bacteria: Lactococcus lactis subsp. cremoris Gram negative bacteria: Escherichia coli, Staphylococcus aureus, Salmonella typhimurium Virus: Rotavirus | Camel milk LF showed intriguing antibacterial activity. The camel milk LP was bacteriostatic against the Gram-positive strains and was bactericidal against Gram-negative cultures. The immunoglobulins had little effect against the bacteria but high titres of antibodies against rotavirus were found in camel milk. The LP system was ineffective against rotavirus | [29] |
| 4 | Camel Casein Proteins and hydrolysates | Gram positive bacteria: Listeria innocua, Bacillus cereus, and Staphylococcus aureus Gram-negative bacteria: Escherichia coli XL1 bleu and Pseudomonas aeruginosa | Camel milk casein hydrolysates exhibited anti-bacterial activity; Gram-positive strain growth was not affected by intact camel casein fraction, whereas the respective hydrolysates slightly inhibited the growth of the bacteria | [83] |
| 5 | Camel and Cow Casein Proteins and hydrolysates | Candida krusei, Candida parapsilosis | Camel milk protein hydrolysates were more potent in inhibiting pathogenic Candida species compared with cow milk protein hydrolysates | [84] |
| S.No | Bioactive Peptide | Mechanism | Reference |
|---|---|---|---|
| Hypertension | |||
| 1 | AIPPKKNQD | Inhibition of Angiotensin-1 converting enzyme (ACE) | [85] |
| 2 | DLENLHLPLPL; LTDLENLHLPLPL;TDLENLHLPLP; TDLENLHLPLPL; TLTDLENLHLPLPL | Inhibition of Angiotensin-1 converting enzyme (ACE) | [86] |
| 3 | LSLSQFKVLPVPQ; KVLPVPQQMVPYPQ;TDLENLHLPLPL | Inhibition of Angiotensin-1 converting enzyme (ACE) | [87] |
| 4 | AEWLHDWKL; SHSPLAGFR; LTMPQWW; CLSPLQMR and CLSPLQFR | Inhibition of Angiotensin-1 converting enzyme (ACE) | [10] |
| 5 | QSAPGNEAIPP | Inhibition of Angiotensin-1 converting enzyme (ACE) | [88] |
| 6 | MVPYPQR | Inhibition of Angiotensin-1 converting enzyme (ACE) | [89] |
| Diabetes | |||
| 1 | FLQY; FQLGASPY; ILDKEGIDY; ILELA; LLQLEAIR; LPVP; LQALHQGQIV; MPVQA; and SPVVPF | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [90] |
| 2 | VPV, YPI and VPF | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [64] |
| 3 | DNLMPQFM and WNWGWLLWQL | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [63] |
| 4 | INNQFLPYPYWL and IPAVF | Inhibition of Dipeptidyl peptidase-IV (DPP-IV) | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, F.B.; Ansari, M.A.; Uddin, S.; Palakott, A.R.; Anwar, I.; Almatroudi, A.; Alomary, M.N.; Alrumaihi, F.; Aba Alkhayl, F.F.; Alghamdi, S.; et al. Prospective Role of Bioactive Molecules and Exosomes in the Therapeutic Potential of Camel Milk against Human Diseases: An Updated Perspective. Life 2022, 12, 990. https://doi.org/10.3390/life12070990
Khan FB, Ansari MA, Uddin S, Palakott AR, Anwar I, Almatroudi A, Alomary MN, Alrumaihi F, Aba Alkhayl FF, Alghamdi S, et al. Prospective Role of Bioactive Molecules and Exosomes in the Therapeutic Potential of Camel Milk against Human Diseases: An Updated Perspective. Life. 2022; 12(7):990. https://doi.org/10.3390/life12070990
Chicago/Turabian StyleKhan, Farheen Badrealam, Mohammad Azam Ansari, Shahab Uddin, Abdul Rasheed Palakott, Irfa Anwar, Ahmad Almatroudi, Mohammad N. Alomary, Faris Alrumaihi, Faris F. Aba Alkhayl, Saad Alghamdi, and et al. 2022. "Prospective Role of Bioactive Molecules and Exosomes in the Therapeutic Potential of Camel Milk against Human Diseases: An Updated Perspective" Life 12, no. 7: 990. https://doi.org/10.3390/life12070990
APA StyleKhan, F. B., Ansari, M. A., Uddin, S., Palakott, A. R., Anwar, I., Almatroudi, A., Alomary, M. N., Alrumaihi, F., Aba Alkhayl, F. F., Alghamdi, S., Muhammad, K., Huang, C.-Y., Daddam, J. R., Khan, H., Maqsood, S., & Ayoub, M. A. (2022). Prospective Role of Bioactive Molecules and Exosomes in the Therapeutic Potential of Camel Milk against Human Diseases: An Updated Perspective. Life, 12(7), 990. https://doi.org/10.3390/life12070990