
Engineering of Synthetic Transcriptional Switches in Yeast



Abstract
1. Introduction
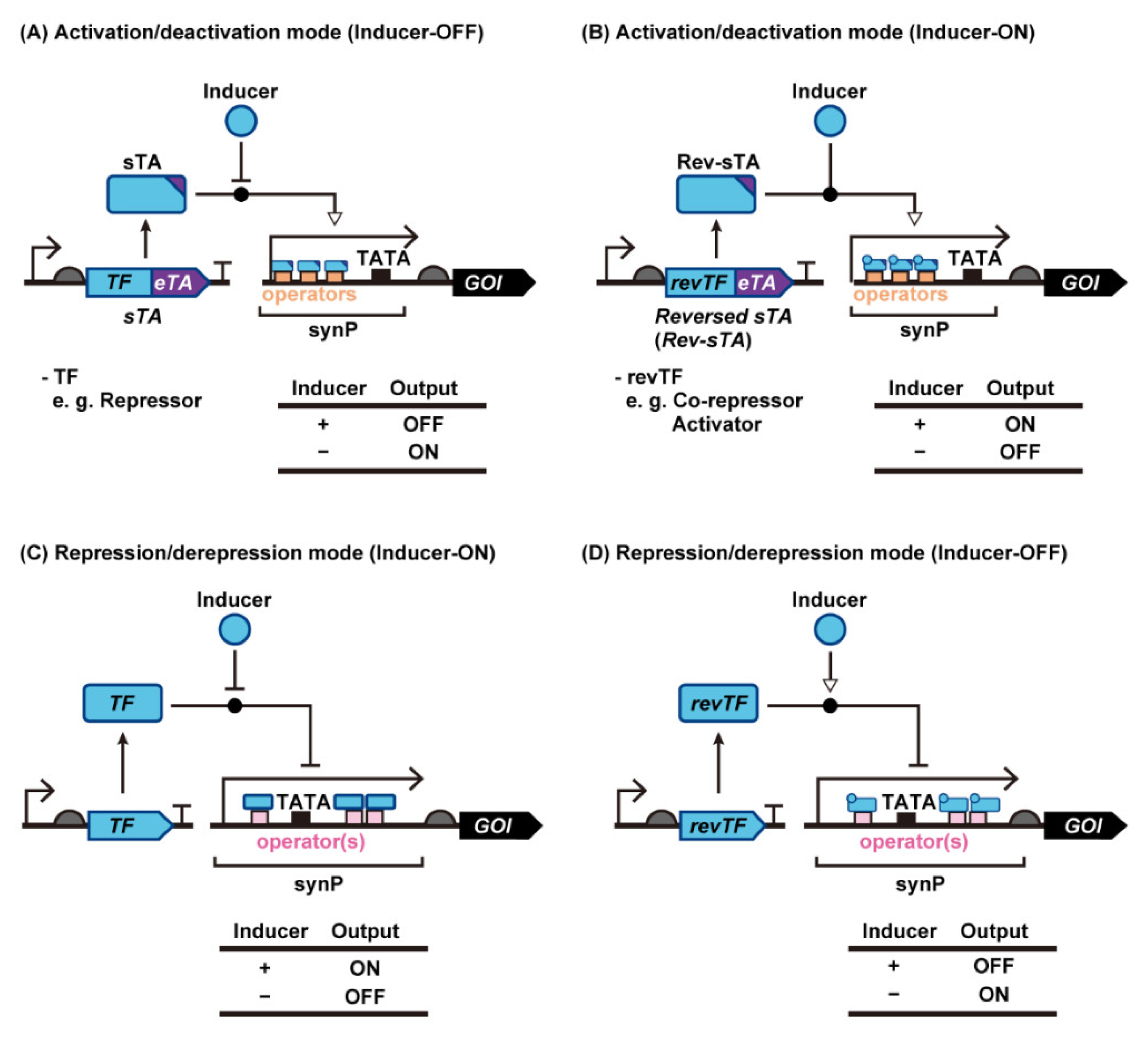
2. Synthetic Transcriptional Switches with Different Modes of Regulation in Yeast
2.1. Transcription Activation Mode
2.1.1. sTAs Based on Bacterial Transcriptional Repressors
2.1.2. sTAs Based on Bacterial Transcriptional Co-Repressors and Activators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inducer a | bTF | bTF Type c | Source | Additional Motif d | Operators e | CoreP f | Reference |
|---|---|---|---|---|---|---|---|
| Dox | Reversed TetR (rTetR) | Co-rep | Escherichia coli | VP16 × 3 | [tetO]7 | PGAL1(Sc) | [45] |
| Dox | rTetR | Co-rep | E. coli | VP16 × 3 | [tetO]7 | PCYC1(Sc) | [39] |
| Dox | rTetR | Co-rep | E. coli | VP16 × 3 | [tetO]3 or 4 | PGAL1(Sc) | [41] |
| Dox | TetR | Rep | E. coli | VP16ad × 1 or 2 | [tetO]1, 2 or 7 | PCYC1(Sc) | [24] |
| DAPG | PhlF | Rep | Pseudomonas fluorescens | VP16 × 3 | [phlO]7 | PCYC1(Sc) | [29] |
| NLS, VP16 × 3 | [phlO]1 | PGAL1(Sc) | [45] | ||||
| Camphor | CamR | Rep | P. putida | NLS, VP16 × 3 | [camO]6 | PCYC1(Sc) | [28,29] |
| [camO]1 | PGAL1(Sc) | [45] | |||||
| Cumate | CymR | Rep | P. putida | NLS, VP16 * | [cymO]6 | PCYC1(Sc) | [29] |
| DAPG | Reversed PhlF (rPhlF) | Rep | P. fluorescens | NLS, VP16 × 3 | [phlO]6 | PGAL1(Sc) | [45] |
| HSL | LuxR | Act | Vibrio fischeri | NLS, VP16 × 3 | [luxO]1 | PGAL1(Sc) | [45] |
| [luxO]5 | PGAL1(Sc) | ||||||
| [luxO]10 | PGAL1(Sc) | ||||||
| SAM | MetJ | Co-Rep | E. coli | NLS, B42 | [metO]1 | PCYC1(Sc) | [46] |
| Methylating compound | N-Ada b | Act | E. coli | GAL4-AD | [AdaOp]1, 3 or 8 | PCYC1(Sc) | [47] |
| Xylose | XylR | Act | E. coli | NLS, VPRH | PTEFup-[Pxo]1 | PTEF(Yl) | [49] |
| NLS, VPRH | [Pxo]1 | PTEF(Yl) | |||||
| NLS, VPR | ND | ND | |||||
| NLS, HSF | ND | ND | |||||
| NLS, VPRH | [Pxo]1 | PLEU(Yl) | |||||
| NLS, VPRH | [Pxo]1 | PYlACC1 | |||||
| NLS, VPRH | [Pxo]1 | PTEF(Sc) | |||||
| Malonyl-CoA | FapR | Rep | Bacillus subtilis | Prm1 | [fapO]1 | PAOX1(Kp) | [36] |
| Benzoate | HbaR | Act | Rhodopseudomonas palustris | B112 | [LexA binding site]8 | PCYC1(Sc) | [48] |
2.1.3. sTAs Based on Ligand-Dependent Nuclear Localization
2.1.4. sTAs Based on Ligand-Induced Protein-Protein Interactions
2.1.5. Transcription Activation without a Eukaryotic Activation Motif
2.2. Transcription Repression Mode
3. Strategies to Improve the Performance of Yeast Transcriptional Switches
3.1. Strategies for Improving Fold-Induction
3.1.1. sTA-Based Yeast Transcriptional Switches
3.1.2. LTTR-Based Yeast Transcriptional Switches
3.1.3. Repressor-Based Yeast Transcriptional Switches
3.1.4. Genetic Circuits: Assembly of Different Switches
3.2. Strategy to Modify Inducer Sensitivity and Specificity, and DNA-Binding Specificity
4. Directed Evolution of Transcriptional Switches in Yeast
4.1. Fluorescence-Based Screening
4.2. ON/OFF Selections Using FACS
4.3. ON/OFF Selections Using Genetic Selections
4.4. “Screening of Selection” Strategy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meyer, A.J.; Segall-Shapiro, T.H.; Glassey, E.; Zhang, J.; Voigt, C.A. Escherichia coli “Marionette” strains with 12 highly optimized small-molecule sensors. Nat. Chem. Biol. 2019, 15, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.A.; Voigt, C.A. Principles of genetic circuit design. Nat. Methods 2014, 11, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.M.; Mao, A.S.; de Puig, H.; Zhang, K.; Tippens, N.D.; Tan, X.; Ran, F.A.; Han, I.; Nguyen, P.Q.; Chory, E.J.; et al. RNA-responsive elements for eukaryotic translational control. Nat. Biotechnol. 2021, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.Q.; Soenksen, L.R.; Donghia, N.M.; Angenent-Mari, N.M.; de Puig, H.; Huang, A.; Lee, R.; Slomovic, S.; Galbersanini, T.; Lansberry, G.; et al. Wearable materials with embedded synthetic biology sensors for biomolecule detection. Nat. Biotechnol. 2021, 39, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.A. Genetic parts to program bacteria. Curr. Opin. Biotechnol. 2006, 17, 548–557. [Google Scholar] [CrossRef]
- Douglas, H.C.; Hawthorne, D.C. Enzymatic Expression and Genetic Linkage of Genes Controlling Galactose Utilization in Saccharomyces. Genetics 1964, 49, 837–844. [Google Scholar] [CrossRef]
- Bassel, J.; Mortimer, R. Genetic order of the galactose structural genes in Saccharomyces cerevisiae. J. Bacteriol. 1971, 108, 179–183. [Google Scholar] [CrossRef]
- Ahn, J.; Park, K.M.; Lee, H.; Son, Y.J.; Choi, E.S. GAL promoter-driven heterologous gene expression in Saccharomyces cerevisiae Delta strain at anaerobic alcoholic fermentation. FEMS Yeast Res. 2013, 13, 140–142. [Google Scholar] [CrossRef][Green Version]
- Vogl, T.; Glieder, A. Regulation of Pichia pastoris promoters and its consequences for protein production. Nat. Biotechnol. 2013, 30, 385–404. [Google Scholar] [CrossRef]
- Gander, M.W.; Vrana, J.D.; Voje, W.E.; Carothers, J.M.; Klavins, E. Digital logic circuits in yeast with CRISPR-dCas9 NOR gates. Nat. Commun. 2017, 8, 15459. [Google Scholar] [CrossRef]
- Ryo, S.; Ishii, J.; Matsuno, T.; Nakamura, Y.; Matsubara, D.; Tominaga, M.; Kondo, A. Positive Feedback Genetic Circuit Incorporating a Constitutively Active Mutant Gal3 into Yeast GAL Induction System. ACS Synth. Biol. 2017, 6, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Rantasalo, A.; Kuivanen, J.; Penttila, M.; Jantti, J.; Mojzita, D. Synthetic Toolkit for Complex Genetic Circuit Engineering in Saccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 1573–1587. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.; Young, E.M.; Jones, T.S.; Densmore, D.; Voigt, C.A. Genetic circuit design automation for yeast. Nat. Microbiol. 2020, 5, 1349–1360. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhao, H.; Zhang, H.; Wang, R.; Huang, J.; Tian, Y.; Luo, X.; Luo, X.; Wang, M.; Xiang, Y.; et al. De novo design of an intercellular signaling toolbox for multi-channel cell-cell communication and biological computation. Nat. Commun. 2020, 11, 4226. [Google Scholar] [CrossRef]
- Adeniran, A.; Sherer, M.; Tyo, K.E.J. Yeast-based biosensors: Design and applications. Fems Yeast Res. 2015, 15, 1–15. [Google Scholar] [CrossRef]
- D’Ambrosio, V.; Jensen, M.K. Lighting up yeast cell factories by transcription factor-based biosensors. FEMS Yeast Res. 2017, 17, fox076. [Google Scholar] [CrossRef]
- Monteiro, F.; Hubmann, G.; Takhaveev, V.; Vedelaar, S.R.; Norder, J.; Hekelaar, J.; Saldida, J.; Litsios, A.; Wijma, H.J.; Schmidt, A.; et al. Measuring glycolytic flux in single yeast cells with an orthogonal synthetic biosensor. Mol. Syst. Biol. 2019, 15, e9071. [Google Scholar] [CrossRef]
- D’Ambrosio, V.; Pramanik, S.; Goroncy, K.; Jakočiūnas, T.; Schönauer, D.; Davari, M.D.; Schwaneberg, U.; Keasling, J.D.; Jensen, M.K. Directed evolution of VanR biosensor specificity in yeast. Biotechnol. Notes 2020, 2020, 9–15. [Google Scholar] [CrossRef]
- Snoek, T.; Chaberski, E.K.; Ambri, F.; Kol, S.; Bjorn, S.P.; Pang, B.; Barajas, J.F.; Welner, D.H.; Jensen, M.K.; Keasling, J.D. Evolution-guided engineering of small-molecule biosensors. Nucleic Acids Res. 2020, 48, e3. [Google Scholar] [CrossRef]
- Shaw, W.M.; Yamauchi, H.; Mead, J.; Gowers, G.F.; Bell, D.J.; Oling, D.; Larsson, N.; Wigglesworth, M.; Ladds, G.; Ellis, T. Engineering a Model Cell for Rational Tuning of GPCR Signaling. Cell 2019, 177, 782–796.e27. [Google Scholar] [CrossRef]
- Gossen, M.; Bonin, A.L.; Freundlieb, S.; Bujard, H. Inducible gene expression systems for higher eukaryotic cells. Curr. Opin. Biotechnol. 1994, 5, 516–520. [Google Scholar] [CrossRef]
- Helmschrott, C.; Sasse, A.; Samantaray, S.; Krappmann, S.; Wagener, J. Upgrading fungal gene expression on demand: Improved systems for doxycycline-dependent silencing in Aspergillus fumigatus. Appl. Environ. Microbiol. 2013, 79, 1751–1754. [Google Scholar] [CrossRef]
- Belli, G.; Gari, E.; Piedrafita, L.; Aldea, M.; Herrero, E. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Res. 1998, 26, 942–947. [Google Scholar] [CrossRef]
- Gari, E.; Piedrafita, L.; Aldea, M.; Herrero, E. A set of vectors with a tetracycline-regulatable promoter system for modulated gene expression in Saccharomyces cerevisiae. Yeast 1997, 13, 837–848. [Google Scholar] [CrossRef]
- Ge, H.; Marchisio, M.A. Aptamers, Riboswitches, and Ribozymes in S. cerevisiae Synthetic Biology. Life 2021, 11, 248. [Google Scholar] [CrossRef]
- Caliando, B.J.; Voigt, C.A. Targeted DNA degradation using a CRISPR device stably carried in the host genome. Nat. Commun. 2015, 6, 6989. [Google Scholar] [CrossRef]
- Loew, R.; Heinz, N.; Hampf, M.; Bujard, H.; Gossen, M. Improved Tet-responsive promoters with minimized background expression. BMC Biotechnol. 2010, 10, 81. [Google Scholar] [CrossRef]
- Ikushima, S.; Zhao, Y.; Boeke, J.D. Development of a Tightly Controlled Off Switch for Saccharomyces cerevisiae Regulated by Camphor, a Low-Cost Natural Product. G3 Genes Genomes Genet. 2015, 5, 1983–1990. [Google Scholar] [CrossRef]
- Ikushima, S.; Boeke, J.D. New Orthogonal Transcriptional Switches Derived from Tet Repressor Homologues for Saccharomyces cerevisiae Regulated by 2,4-Diacetylphloroglucinol and Other Ligands. ACS Synth. Biol. 2017, 6, 497–506. [Google Scholar] [CrossRef]
- Gossen, M.; Freundlieb, S.; Bender, G.; Muller, G.; Hillen, W.; Bujard, H. Transcriptional activation by tetracyclines in mammalian cells. Science 1995, 268, 1766–1769. [Google Scholar] [CrossRef]
- Boyer, J.; Badis, G.; Fairhead, C.; Talla, E.; Hantraye, F.; Fabre, E.; Fischer, G.; Hennequin, C.; Koszul, R.; Lafontaine, I.; et al. Large-scale exploration of growth inhibition caused by overexpression of genomic fragments in Saccharomyces cerevisiae. Genome Biol. 2004, 5, R72. [Google Scholar] [CrossRef]
- Roemer, T.; Jiang, B.; Davison, J.; Ketela, T.; Veillette, K.; Breton, A.; Tandia, F.; Linteau, A.; Sillaots, S.; Marta, C.; et al. Large-scale essential gene identification in Candida albicans and applications to antifungal drug discovery. Mol. Microbiol. 2003, 50, 167–181. [Google Scholar] [CrossRef]
- Tabuchi, M.; Kawai, Y.; Nishie-Fujita, M.; Akada, R.; Izumi, T.; Yanatori, I.; Miyashita, N.; Ouchi, K.; Kishi, F. Development of a novel functional high-throughput screening system for pathogen effectors in the yeast Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 2009, 73, 2261–2267. [Google Scholar] [CrossRef][Green Version]
- Cuthbertson, L.; Nodwell, J.R. The TetR Family of Regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef]
- Qiu, C.; Chen, X.; Rexida, R.; Shen, Y.; Qi, Q.; Bao, X.; Hou, J. Engineering transcription factor-based biosensors for repressive regulation through transcriptional deactivation design in Saccharomyces cerevisiae. Microb. Cell Factories 2020, 19, 146. [Google Scholar] [CrossRef]
- Wen, J.; Tian, L.; Xu, M.; Zhou, X.; Zhang, Y.; Cai, M. A Synthetic Malonyl-CoA Metabolic Oscillator in Komagataella phaffii. ACS Synth. Biol. 2020, 9, 1059–1068. [Google Scholar] [CrossRef]
- Umeno, D.; Kimura, Y.; Kawai-Noma, S. Transcription Factors as Evolvable Biosensors. Anal. Sci. 2021, 37, 699–705. [Google Scholar] [CrossRef]
- Das, A.T.; Tenenbaum, L.; Berkhout, B. Tet-On Systems For Doxycycline-inducible Gene Expression. Curr. Gene Ther. 2016, 16, 156–167. [Google Scholar] [CrossRef]
- Urlinger, S.; Baron, U.; Thellmann, M.; Hasan, M.T.; Bujard, H.; Hillen, W. Exploring the sequence space for tetracycline-dependent transcriptional activators: Novel mutations yield expanded range and sensitivity. Proc. Natl. Acad. Sci. USA 2000, 97, 7963–7968. [Google Scholar] [CrossRef]
- Das, A.T.; Zhou, X.; Vink, M.; Klaver, B.; Verhoef, K.; Marzio, G.; Berkhout, B. Viral evolution as a tool to improve the tetracycline-regulated gene expression system. J. Biol. Chem. 2004, 279, 18776–18782. [Google Scholar] [CrossRef]
- Roney, I.J.; Rudner, A.D.; Couture, J.F.; Kaern, M. Improvement of the reverse tetracycline transactivator by single amino acid substitutions that reduce leaky target gene expression to undetectable levels. Sci. Rep. 2016, 6, 27697. [Google Scholar] [CrossRef] [PubMed]
- Becskei, A.; Seraphin, B.; Serrano, L. Positive feedback in eukaryotic gene networks: Cell differentiation by graded to binary response conversion. EMBO J. 2001, 20, 2528–2535. [Google Scholar] [CrossRef] [PubMed]
- Zhuravel, D.; Fraser, D.; St-Pierre, S.; Tepliakova, L.; Pang, W.L.; Hasty, J.; Kaern, M. Phenotypic impact of regulatory noise in cellular stress-response pathways. Syst. Synth. Biol. 2010, 4, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.; Ray, J.C.; Manhart, M.; Adams, R.M.; Nevozhay, D.; Morozov, A.V.; Balazsi, G. Stress-response balance drives the evolution of a network module and its host genome. Mol. Syst. Biol. 2015, 11, 827. [Google Scholar] [CrossRef]
- Tominaga, M.; Nozaki, K.; Umeno, D.; Ishii, J.; Kondo, A. Robust and flexible platform for directed evolution of yeast genetic switches. Nat. Commun. 2021, 12, 1846. [Google Scholar] [CrossRef]
- Umeyama, T.; Okada, S.; Ito, T. Synthetic gene circuit-mediated monitoring of endogenous metabolites: Identification of GAL11 as a novel multicopy enhancer of s-adenosylmethionine level in yeast. ACS Synth. Biol. 2013, 2, 425–430. [Google Scholar] [CrossRef]
- Moser, F.; Horwitz, A.; Chen, J.; Lim, W.; Voigt, C.A. Genetic sensor for strong methylating compounds. ACS Synth. Biol. 2013, 2, 614–624. [Google Scholar] [CrossRef]
- Castano-Cerezo, S.; Fournie, M.; Urban, P.; Faulon, J.L.; Truan, G. Development of a Biosensor for Detection of Benzoic Acid Derivatives in Saccharomyces cerevisiae. Front. Bioeng. Biotechnol. 2019, 7, 372. [Google Scholar] [CrossRef]
- Wei, W.; Shang, Y.; Zhang, P.; Liu, Y.; You, D.; Yin, B.; Ye, B. Engineering Prokaryotic Transcriptional Activator XylR as a Xylose-Inducible Biosensor for Transcription Activation in Yeast. ACS Synth. Biol. 2020, 9, 1022–1029. [Google Scholar] [CrossRef]
- Louvion, J.F.; Havaux-Copf, B.; Picard, D. Fusion of GAL4-VP16 to a steroid-binding domain provides a tool for gratuitous induction of galactose-responsive genes in yeast. Gene 1993, 131, 129–134. [Google Scholar] [CrossRef]
- McIsaac, R.S.; Silverman, S.J.; McClean, M.N.; Gibney, P.A.; Macinskas, J.; Hickman, M.J.; Petti, A.A.; Botstein, D. Fast-acting and nearly gratuitous induction of gene expression and protein depletion in Saccharomyces cerevisiae. Mol. Biol. Cell 2011, 22, 4447–4459. [Google Scholar] [CrossRef]
- McIsaac, R.S.; Gibney, P.A.; Chandran, S.S.; Benjamin, K.R.; Botstein, D. Synthetic biology tools for programming gene expression without nutritional perturbations in Saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, e48. [Google Scholar] [CrossRef]
- Gao, C.Y.; Pinkham, J.L. Tightly regulated, beta-estradiol dose-dependent expression system for yeast. Biotechniques 2000, 29, 1226–1231. [Google Scholar] [CrossRef]
- Quintero, M.J.; Maya, D.; Arevalo-Rodriguez, M.; Cebolla, A.; Chavez, S. An improved system for estradiol-dependent regulation of gene expression in yeast. Microb. Cell Fact. 2007, 6, 10. [Google Scholar] [CrossRef]
- Ottoz, D.S.; Rudolf, F.; Stelling, J. Inducible, tightly regulated and growth condition-independent transcription factor in Saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, e130. [Google Scholar] [CrossRef]
- Mormino, M.; Siewers, V.; Nygard, Y. Development of an Haa1-based biosensor for acetic acid sensing in Saccharomyces cerevisiae. FEMS Yeast Res. 2021, 21, foab049. [Google Scholar] [CrossRef]
- Niopek, D.; Benzinger, D.; Roensch, J.; Draebing, T.; Wehler, P.; Eils, R.; Di Ventura, B. Engineering light-inducible nuclear localization signals for precise spatiotemporal control of protein dynamics in living cells. Nat. Commun. 2014, 5, 4404. [Google Scholar] [CrossRef]
- Geller, S.H.; Antwi, E.B.; Di Ventura, B.; McClean, M.N. Optogenetic Repressors of Gene Expression in Yeasts Using Light-Controlled Nuclear Localization. Cell Mol. Bioeng. 2019, 12, 511–528. [Google Scholar] [CrossRef]
- Fields, S.; Song, O. A novel genetic system to detect protein-protein interactions. Nature 1989, 340, 245–246. [Google Scholar] [CrossRef]
- Chockalingam, K.; Chen, Z.; Katzenellenbogen, J.A.; Zhao, H. Directed evolution of specific receptor-ligand pairs for use in the creation of gene switches. Proc. Natl. Acad. Sci. USA 2005, 102, 5691–5696. [Google Scholar] [CrossRef]
- Salinas, F.; Rojas, V.; Delgado, V.; Lopez, J.; Agosin, E.; Larrondo, L.F. Fungal Light-Oxygen-Voltage Domains for Optogenetic Control of Gene Expression and Flocculation in Yeast. Mbio 2018, 9, e00626-18. [Google Scholar] [CrossRef]
- Xu, X.P.; Du, Z.X.; Liu, R.M.; Li, T.; Zhao, Y.Z.; Chen, X.J.; Yang, Y. A Single-Component Optogenetic System Allows Stringent Switch of Gene Expression in Yeast Cells. ACS Synth. Biol. 2018, 7, 2045–2053. [Google Scholar] [CrossRef]
- Zhao, E.M.; Lalwani, M.A.; Lovelett, R.J.; Garcia-Echauri, S.A.; Hoffman, S.M.; Gonzalez, C.L.; Toettcher, J.E.; Kevrekidis, I.G.; Avalos, J.L. Design and Characterization of Rapid Optogenetic Circuits for Dynamic Control in Yeast Metabolic Engineering. ACS Synth. Biol. 2020, 9, 3254–3266. [Google Scholar] [CrossRef]
- Romero, A.; Rojas, V.; Delgado, V.; Salinas, F.; Larrondo, L.F. Modular and Molecular Optimization of a LOV (Light-Oxygen-Voltage)-Based Optogenetic Switch in Yeast. Int J. Mol. Sci. 2021, 22, 8538. [Google Scholar] [CrossRef]
- Chou, H.H.; Keasling, J.D. Programming adaptive control to evolve increased metabolite production. Nat. Commun. 2013, 4, 2595. [Google Scholar] [CrossRef]
- Skjoedt, M.L.; Snoek, T.; Kildegaard, K.R.; Arsovska, D.; Eichenberger, M.; Goedecke, T.J.; Rajkumar, A.S.; Zhang, J.; Kristensen, M.; Lehka, B.J.; et al. Engineering prokaryotic transcriptional activators as metabolite biosensors in yeast. Nat. Chem. Biol. 2016, 12, 951–958. [Google Scholar] [CrossRef]
- Ambri, F.; D’Ambrosio, V.; Di Blasi, R.; Maury, J.; Jacobsen, S.A.B.; McCloskey, D.; Jensen, M.K.; Keasling, J.D. High-Resolution Scanning of Optimal Biosensor Reporter Promoters in Yeast. ACS Synth. Biol. 2020, 9, 218–226. [Google Scholar] [CrossRef]
- Ellis, T.; Wang, X.; Collins, J.J. Diversity-based, model-guided construction of synthetic gene networks with predicted functions. Nat. Biotechnol. 2009, 27, 465–471. [Google Scholar] [CrossRef]
- Blount, B.A.; Weenink, T.; Vasylechko, S.; Ellis, T. Rational diversification of a promoter providing fine-tuned expression and orthogonal regulation for synthetic biology. PLoS ONE 2012, 7, e33279. [Google Scholar] [CrossRef]
- Teo, W.S.; Chang, M.W. Bacterial XylRs and synthetic promoters function as genetically encoded xylose biosensors in Saccharomyces cerevisiae. Biotechnol. J. 2015, 10, 315–322. [Google Scholar] [CrossRef]
- Wang, M.; Li, S.; Zhao, H. Design and engineering of intracellular-metabolite-sensing/regulation gene circuits in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2016, 113, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Hector, R.E.; Mertens, J.A. A Synthetic Hybrid Promoter for Xylose-Regulated Control of Gene Expression in Saccharomyces Yeasts. Mol. Biotechnol. 2017, 59, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F.; Cress, B.F.; Yang, Z.; Hordines, J.C.; Zhao, S.J.; Jung, G.Y.; Wang, Z.T.; Koffas, M.A.G. Design and Characterization of Biosensors for the Screening of Modular Assembled Naringenin Biosynthetic Library in Saccharomyces cerevisiae. ACS Synth. Biol. 2019, 8, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Teo, W.S.; Hee, K.S.; Chang, M.W. Bacterial FadR and synthetic promoters function as modular fatty acid sensor- regulators in Saccharomyces cerevisiae. Eng. Life Sci. 2013, 13, 456–463. [Google Scholar] [CrossRef]
- David, F.; Nielsen, J.; Siewers, V. Flux Control at the Malonyl-CoA Node through Hierarchical Dynamic Pathway Regulation in Saccharomyces cerevisiae. ACS Synth. Biol. 2016, 5, 224–233. [Google Scholar] [CrossRef]
- Li, S.; Si, T.; Wang, M.; Zhao, H. Development of a Synthetic Malonyl-CoA Sensor in Saccharomyces cerevisiae for Intracellular Metabolite Monitoring and Genetic Screening. ACS Synth. Biol. 2015, 4, 1308–1315. [Google Scholar] [CrossRef]
- Cao, J.; Perez-Pinera, P.; Lowenhaupt, K.; Wu, M.R.; Purcell, O.; de la Fuente-Nunez, C.; Lu, T.K. Versatile and on-demand biologics co-production in yeast. Nat. Commun. 2018, 9, 77. [Google Scholar] [CrossRef]
- Kjaerulff, S.; Nielsen, O. An IPTG-inducible derivative of the fission yeast nmt promoter. Yeast 2015, 32, 469–478. [Google Scholar] [CrossRef]
- Jensen, M.K. Design principles for nuclease-deficient CRISPR-based transcriptional regulators. FEMS Yeast Res. 2018, 18, foy039. [Google Scholar] [CrossRef]
- Mazumder, M.; McMillen, D.R. Design and characterization of a dual-mode promoter with activation and repression capability for tuning gene expression in yeast. Nucleic Acids Res. 2014, 42, 9514–9522. [Google Scholar] [CrossRef]
- Naseri, G.; Balazadeh, S.; Machens, F.; Kamranfar, I.; Messerschmidt, K.; Mueller-Roeber, B. Plant-Derived Transcription Factors for Orthologous Regulation of Gene Expression in the Yeast Saccharomyces cerevisiae. ACS Synth. Biol. 2017, 6, 1742–1756. [Google Scholar] [CrossRef]
- Naseri, G.; Prause, K.; Hamdo, H.H.; Arenz, C. Artificial Transcription Factors for Tuneable Gene Expression in Pichia pastoris. Front. Bioeng. Biotechnol. 2021, 9, 676900. [Google Scholar] [CrossRef]
- Islam, K.M.D.; Dilcher, M.; Thurow, C.; Vock, C.; Krimmelbein, I.K.; Tietze, L.F.; Gonzalez, V.; Zhao, H.M.; Gatz, C. Directed evolution of estrogen receptor proteins with altered ligand-binding specificities. Protein Eng. Des. Sel. 2009, 22, 45–52. [Google Scholar] [CrossRef]
- Dimas, R.P.; Jordan, B.R.; Jiang, X.L.; Martini, C.; Glavy, J.S.; Patterson, D.P.; Morcos, F.; Chan, C.T.Y. Engineering DNA recognition and allosteric response properties of TetR family proteins by using a module-swapping strategy. Nucleic Acids Res. 2019, 47, 8913–8925. [Google Scholar] [CrossRef]
- Juarez, J.F.; Lecube-Azpeitia, B.; Brown, S.L.; Johnston, C.D.; Church, G.M. Biosensor libraries harness large classes of binding domains for construction of allosteric transcriptional regulators. Nat. Commun. 2018, 9, 3101. [Google Scholar] [CrossRef]
- Meinhardt, S.; Manley, M.W., Jr.; Becker, N.A.; Hessman, J.A.; Maher, L.J., 3rd; Swint-Kruse, L. Novel insights from hybrid LacI/GalR proteins: Family-wide functional attributes and biologically significant variation in transcription repression. Nucleic Acids Res. 2012, 40, 11139–11154. [Google Scholar] [CrossRef]
- Muranaka, N.; Sharma, V.; Nomura, Y.; Yokobayashi, Y. An efficient platform for genetic selection and screening of gene switches in Escherichia coli. Nucleic Acids Res. 2009, 37, e39. [Google Scholar] [CrossRef]
- Shong, J.; Huang, Y.M.; Bystroff, C.; Collins, C.H. Directed evolution of the quorum-sensing regulator EsaR for increased signal sensitivity. ACS Chem. Biol. 2013, 8, 789–795. [Google Scholar] [CrossRef]
- Ike, K.; Arasawa, Y.; Koizumi, S.; Mihashi, S.; Kawai-Noma, S.; Saito, K.; Umeno, D. Evolutionary Design of Choline-Inducible and -Repressible T7-Based Induction Systems. ACS Synth. Biol. 2015, 4, 1352–1360. [Google Scholar] [CrossRef]
- Tashiro, Y.; Kimura, Y.; Furubayashi, M.; Tanaka, A.; Terakubo, K.; Saito, K.; Kawai-Noma, S.; Umeno, D. Directed evolution of the autoinducer selectivity of Vibrio fischeri LuxR. J. Gen. Appl. Microbiol. 2016, 62, 240–247. [Google Scholar] [CrossRef]
- Saeki, K.; Tominaga, M.; Kawai-Noma, S.; Saito, K.; Umeno, D. Rapid Diversification of BetI-Based Transcriptional Switches for the Control of Biosynthetic Pathways and Genetic Circuits. ACS Synth. Biol. 2016, 5, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, B.; Peters, G.; Coussement, P.; Maertens, J.; De Mey, M. Tailor-made transcriptional biosensors for optimizing microbial cell factories. J. Ind. Microbiol. Biotechnol. 2017, 44, 623–645. [Google Scholar] [CrossRef] [PubMed]
- Ellefson, J.W.; Ledbetter, M.P.; Ellington, A.D. Directed evolution of a synthetic phylogeny of programmable Trp repressors. Nat. Chem. Biol. 2018, 14, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.J.; Parikh, R.Y.; Weitz, J.S.; Kim, H.D. Suppression of Expression Between Adjacent Genes Within Heterologous Modules in Yeast. G3 Genes Genomes Genet. 2014, 4, 109–116. [Google Scholar] [CrossRef]
- Redden, H.; Alper, H.S. The development and characterization of synthetic minimal yeast promoters. Nat. Commun. 2015, 6, 7810. [Google Scholar] [CrossRef]
- Schaerli, Y.; Isalan, M. Building synthetic gene circuits from combinatorial libraries: Screening and selection strategies. Mol. Biosyst. 2013, 9, 1559–1567. [Google Scholar] [CrossRef]
- Becker, S.; Schmoldt, H.U.; Adams, T.M.; Wilhelm, S.; Kolmar, H. Ultra-high-throughput screening based on cell-surface display and fluorescence-activated cell sorting for the identification of novel biocatalysts. Curr. Opin. Biotechnol. 2004, 15, 323–329. [Google Scholar] [CrossRef]
- Yang, G.; Withers, S.G. Ultrahigh-throughput FACS-based screening for directed enzyme evolution. ChemBioChem 2009, 10, 2704–2715. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef]
- Klauser, B.; Atanasov, J.; Siewert, L.K.; Hartig, J.S. Ribozyme-based aminoglycoside switches of gene expression engineered by genetic selection in S. cerevisiae. ACS Synth Biol 2015, 4, 516–525. [Google Scholar] [CrossRef]
- Johansson, B.; Hahn-Hagerdal, B. Overproduction of pentose phosphate pathway enzymes using a new CRE-loxP expression vector for repeated genomic integration in Saccharomyces cerevisiae. Yeast 2002, 19, 225–231. [Google Scholar] [CrossRef]
- Gnugge, R.; Rudolf, F. Saccharomyces cerevisiae Shuttle vectors. Yeast 2017, 34, 205–221. [Google Scholar] [CrossRef]
- Alexander, W.G.; Doering, D.T.; Hittinger, C.T. High-efficiency genome editing and allele replacement in prototrophic and wild strains of Saccharomyces. Genetics 2014, 198, 859–866. [Google Scholar] [CrossRef]
- Rugbjerg, P.; Genee, H.J.; Jensen, K.; Sarup-Lytzen, K.; Sommer, M.O. Molecular Buffers Permit Sensitivity Tuning and Inversion of Riboswitch Signals. ACS Synth. Biol. 2016, 5, 632–638. [Google Scholar] [CrossRef]
- Machado, L.F.M.; Currin, A.; Dixon, N. Directed evolution of the PcaV allosteric transcription factor to generate a biosensor for aromatic aldehydes. J. Biol. Eng. 2019, 13, 91. [Google Scholar] [CrossRef]
- Chouichit, P.; Whangsuk, W.; Sallabhan, R.; Mongkolsuk, S.; Loprasert, S. A highly sensitive biosensor with a single-copy evolved sensing cassette for chlorpyrifos pesticide detection. Microbiology 2020, 166, 1019–1024. [Google Scholar] [CrossRef]
- Della Corte, D.; van Beek, H.L.; Syberg, F.; Schallmey, M.; Tobola, F.; Cormann, K.U.; Schlicker, C.; Baumann, P.T.; Krumbach, K.; Sokolowsky, S.; et al. Engineering and application of a biosensor with focused ligand specificity. Nat. Commun. 2020, 11, 4851. [Google Scholar] [CrossRef]
- Jia, X.; Ma, Y.; Bu, R.; Zhao, T.; Wu, K. Directed evolution of a transcription factor PbrR to improve lead selectivity and reduce zinc interference through dual selection. AMB Express 2020, 10, 67. [Google Scholar] [CrossRef]
- Kwon, K.K.; Yeom, S.J.; Choi, S.L.; Rha, E.; Lee, H.; Kim, H.; Lee, D.H.; Lee, S.G. Acclimation of bacterial cell state for high-throughput enzyme engineering using a DmpR-dependent transcriptional activation system. Sci. Rep. 2020, 10, 6091. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, H.; Ang, E.L. A New Biosensor for Stilbenes and a Cannabinoid Enabled by Genome Mining of a Transcriptional Regulator. ACS Synth. Biol. 2020, 9, 698–705. [Google Scholar] [CrossRef]
- Tang, R.Q.; Wagner, J.M.; Alper, H.S.; Zhao, X.Q.; Bai, F.W. Design, Evolution, and Characterization of a Xylose Biosensor in Escherichia coli Using the XylR/xylO System with an Expanded Operating Range. ACS Synth. Biol. 2020, 9, 2714–2722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cao, Y.; Liu, Y.; Liu, L.; Li, J.; Du, G.; Chen, J. Development and optimization of N-acetylneuraminic acid biosensors in Bacillus subtilis. Biotechnol. Appl. Biochem. 2020, 67, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Unger, E.K.; Keller, J.P.; Altermatt, M.; Liang, R.Q.; Matsui, A.; Dong, C.Y.; Hon, O.J.; Yao, Z.; Sun, J.Q.; Banala, S.; et al. Directed Evolution of a Selective and Sensitive Serotonin Sensor via Machine Learning. Cell 2020, 183, 1986. [Google Scholar] [CrossRef] [PubMed]
- Crook, N.; Abatemarco, J.; Sun, J.; Wagner, J.M.; Schmitz, A.; Alper, H.S. In vivo continuous evolution of genes and pathways in yeast. Nat. Commun. 2016, 7, 13051. [Google Scholar] [CrossRef]
- Ravikumar, A.; Arzumanyan, G.A.; Obadi, M.K.A.; Javanpour, A.A.; Liu, C.C. Scalable, Continuous Evolution of Genes at Mutation Rates above Genomic Error Thresholds. Cell 2018, 175, 1946–1957.e13. [Google Scholar] [CrossRef]
- Tou, C.J.; Schaffer, D.V.; Dueber, J.E. Targeted Diversification in the S. cerevisiae Genome with CRISPR-Guided DNA Polymerase I. ACS Synth. Biol. 2020, 9, 1911–1916. [Google Scholar] [CrossRef]
- Zhong, Z.; Wong, B.G.; Ravikumar, A.; Arzumanyan, G.A.; Khalil, A.S.; Liu, C.C. Automated Continuous Evolution of Proteins in Vivo. ACS Synth. Biol. 2020, 9, 1270–1276. [Google Scholar] [CrossRef]
- Jensen, E.D.; Ambri, F.; Bendtsen, M.B.; Javanpour, A.A.; Liu, C.C.; Jensen, M.K.; Keasling, J.D. Integrating continuous hypermutation with high-throughput screening for optimization of cis,cis-muconic acid production in yeast. Microb. Biotechnol. 2021, 14, 2617–2626. [Google Scholar] [CrossRef]
- Marsafari, M.; Ma, J.B.; Koffas, M.; Xu, P. Genetically-encoded biosensors for analyzing and controlling cellular process in yeast. Curr. Opin. Biotechnol. 2020, 64, 175–182. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, S. Transcription Factor-Based Biosensor for Dynamic Control in Yeast for Natural Product Synthesis. Front. Bioeng Biotechnol 2021, 9, 635265. [Google Scholar] [CrossRef]
- Mullick, A.; Xu, Y.; Warren, R.; Koutroumanis, M.; Guilbault, C.; Broussau, S.; Malenfant, F.; Bourget, L.; Lamoureux, L.; Lo, R.; et al. The cumate gene-switch: A system for regulated expression in mammalian cells. BMC Biotechnol. 2006, 6, 43. [Google Scholar] [CrossRef]
- Nevozhay, D.; Zal, T.; Balazsi, G. Transferring a synthetic gene circuit from yeast to mammalian cells. Nat. Commun. 2013, 4, 1451. [Google Scholar] [CrossRef]
- Feng, J.; Jester, B.W.; Tinberg, C.E.; Mandell, D.J.; Antunes, M.S.; Chari, R.; Morey, K.J.; Rios, X.; Medford, J.I.; Church, G.M.; et al. A general strategy to construct small molecule biosensors in eukaryotes. Elife 2015, 4, e10606. [Google Scholar] [CrossRef]






| Inducer a | bTF | Source of bTF and Operator | Additional Motif b | Operators | Yeast Promoter d | Reference |
|---|---|---|---|---|---|---|
| aTc | TetR | Esherichia coli | NLS | [tetO]2 | Ptet(Sc) e | [13] |
| aTc | TetR | E. coli | – | [tetO]2 | PLibT f | [68] |
| aTc | TetR | E. coli | – | [tetO]2 | PPFY1(Sc) | [69] |
| DAPG | PhlF | Pseudomonas fluorescens | NLS | [phlO]1 or 2 | PADH1(Sc) | [29] |
| Vanilic acid | VanR | Caulobacter crescentus | – | [vanO]2 | PTEF1(Sc) orPCYC1(Sc) | [18] |
| Vanillin | ||||||
| Naringenin | FdeR | Herbaspirillum seropedicae | NLS | [fdeO]1 | PGPM1 (Sc) | [73] |
| Fatty acids | FadR | E. coli | NLS | [fadBA_EC]1 or 3 | PGAL1 (Sc) | [74] |
| Vibrio cholerae | – | [fadBA_VC]1 or 3 | ||||
| Malonyl-CoA | FapR | Bacillus subtilis | Prm1 | [fapO]1–3 | PGAP (Kp) | [36] |
| B. subtilis | NLS | [fapO]1 or 2 | PGPM1 (Sc) | [76] | ||
| B. subtilis | NLS | [fapO]1–3 | PTEF1 (Sc) | [75] | ||
| Xylose | XylR | B. licheniformis | NLS | [xylO]2 | Pxyl(Sc)e | [13] |
| B. subtilis | ||||||
| Tetragenococcus halophile | ||||||
| Clostridium difficile | ||||||
| Lactobacillus pentosus | ||||||
| Caulobacter crescentus | ||||||
| T. halophile | NLS | [xylO]1 | UEETEF1-PGAL1m(Sc) g | [70] | ||
| C. difficile | NLS | [xylO]1 | UEETEF1-PGAL1m(Sc) g | |||
| L. pentosus | NLS | [xylO]1 | UEETEF1-PGAL1m(Sc) g | |||
| C. crescentus | NLS, Ssn6 | [xylO]1 or 2 | PTEF(Ag) | [72] | ||
| Staphylococcus xylosus | NLS | [xylO]1 or 2 c | PGPM1 (Sc) | [71] | ||
| B. licheniformis | NLS | [xylO]1 or 2 c | PGPM1 (Sc) | |||
| B. subtilis | NLS | [xylO]1 or 2 c | PGPM1 (Sc) | |||
| IPTG | LacI | E. coli | NLS | [lacO]2 | Plac (Sc)e | [13] |
| NLS | [lacO]1 | Pnmt(Szp) | [78] | |||
| – | [lacO]2 | PGAP(Kp) | [77] | |||
| NLS | [lacO]1 | PLibL f | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tominaga, M.; Kondo, A.; Ishii, J. Engineering of Synthetic Transcriptional Switches in Yeast. Life 2022, 12, 557. https://doi.org/10.3390/life12040557
Tominaga M, Kondo A, Ishii J. Engineering of Synthetic Transcriptional Switches in Yeast. Life. 2022; 12(4):557. https://doi.org/10.3390/life12040557
Chicago/Turabian StyleTominaga, Masahiro, Akihiko Kondo, and Jun Ishii. 2022. "Engineering of Synthetic Transcriptional Switches in Yeast" Life 12, no. 4: 557. https://doi.org/10.3390/life12040557
APA StyleTominaga, M., Kondo, A., & Ishii, J. (2022). Engineering of Synthetic Transcriptional Switches in Yeast. Life, 12(4), 557. https://doi.org/10.3390/life12040557