
Mango “Ataulfo” Peel Extract Improves Metabolic Dysregulation in Prediabetic Wistar Rats

 ,
,  ,
,  , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Mango Peel Extract (MPE)
2.2.1. Extract Preparation
2.2.2. Total Phenolic Content
2.2.3. Total Flavonoids
2.2.4. Antioxidant Capacity
2.2.5. Chromatographic Identification of Phenolic Compounds
2.2.6. Reducing Sugars
2.3. Enzyme Inhibition
2.3.1. α-Amylase Activity Assay
2.3.2. α-Glucosidase Activity Assay
2.4. Bioassay
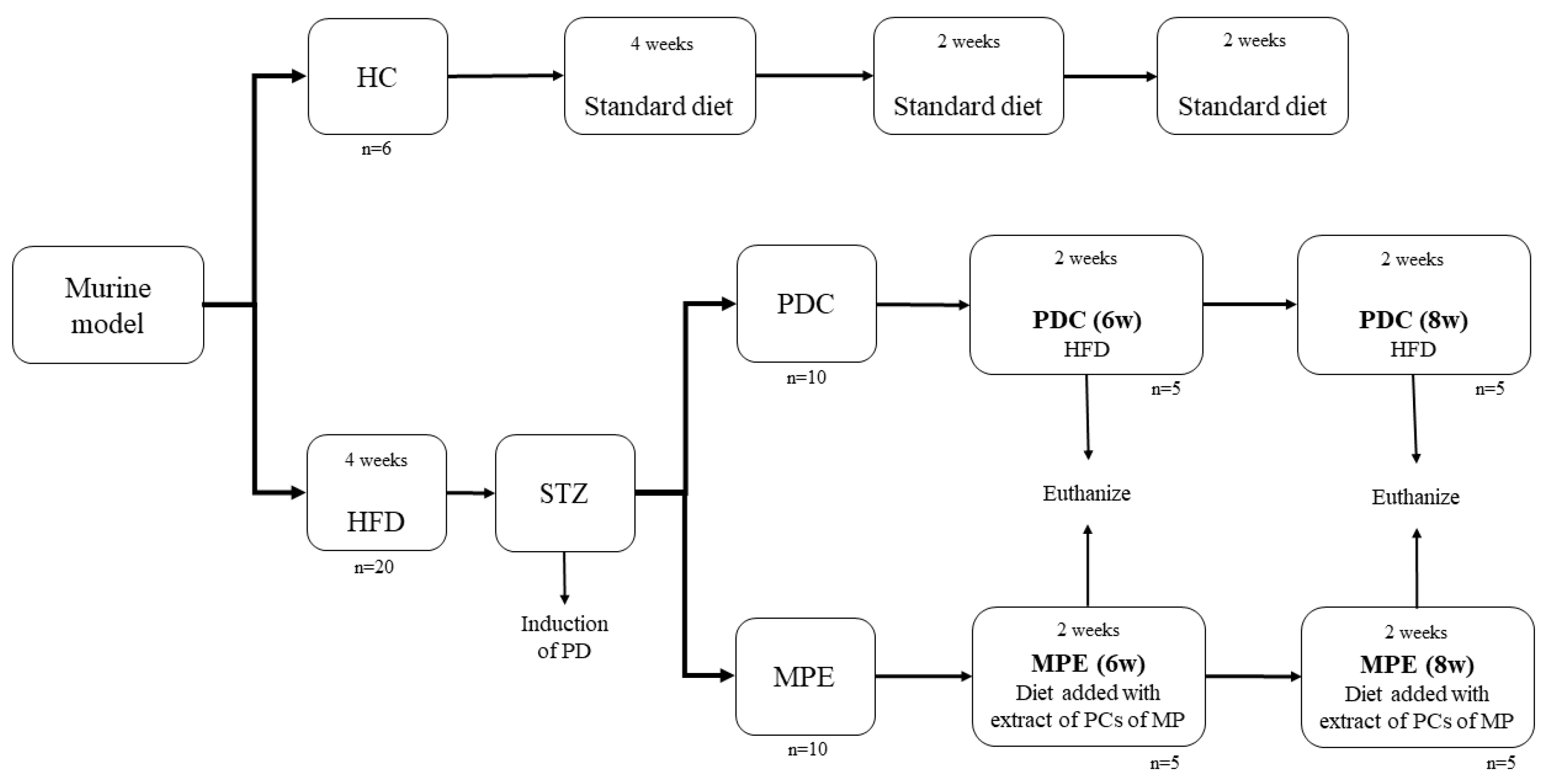
2.4.1. Animals and Diets
2.4.2. PD Induction
2.4.3. Biochemical Parameters
2.4.4. Oral Glucose Tolerance Test (OGTT)
2.4.5. Insulin Sensitivity
2.4.6. Statistical Analysis
3. Results and Discussion
3.1. Chemical Profile of MPE
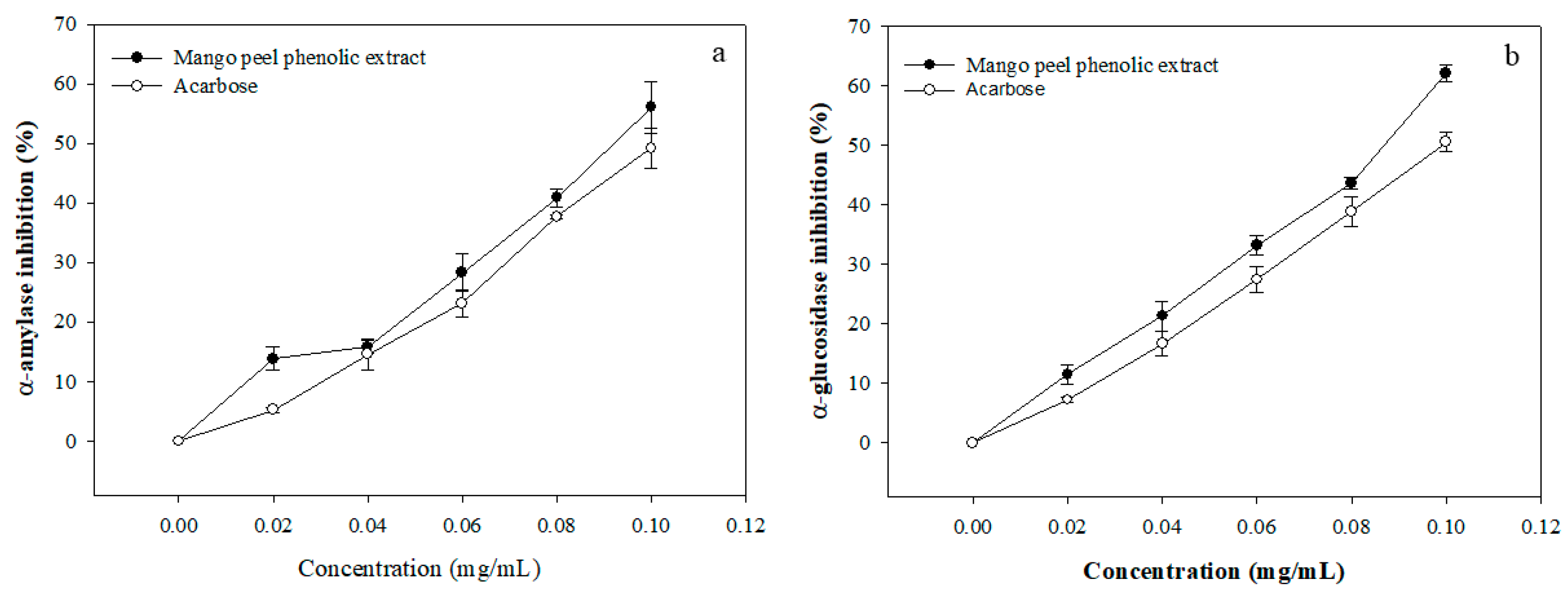
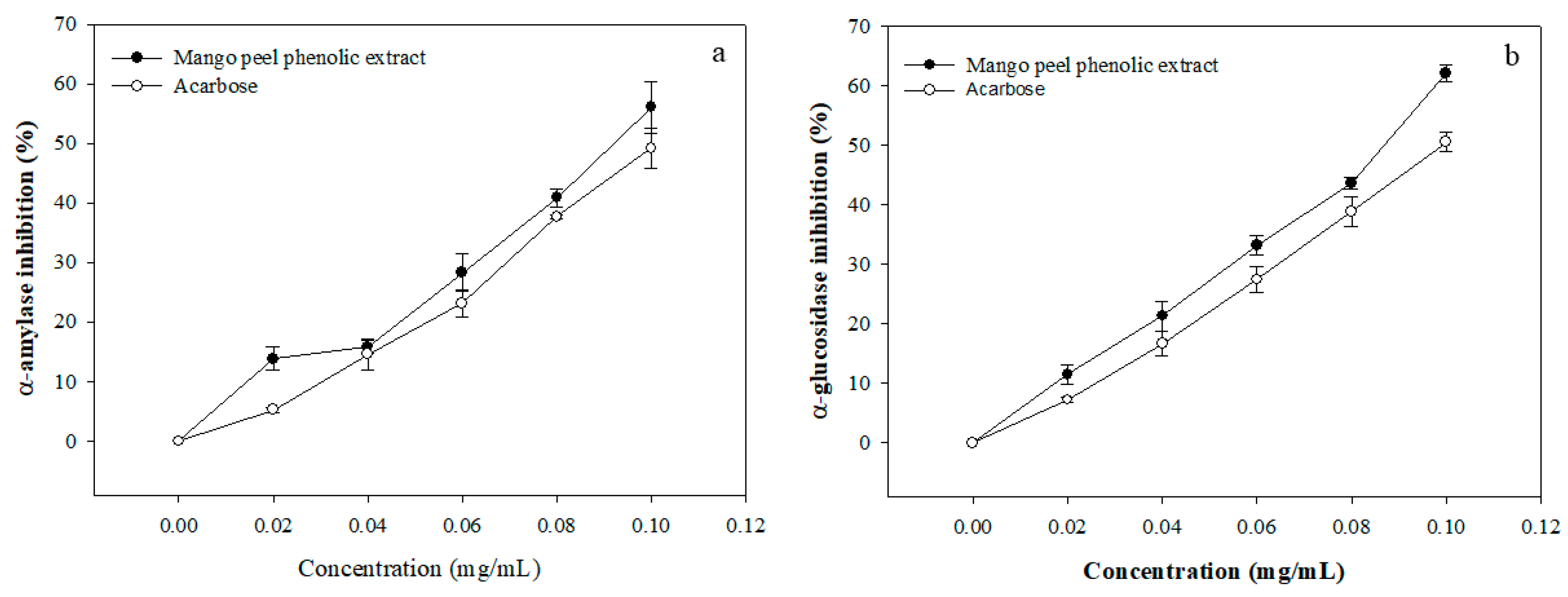
3.2. Enzyme Inhibitory Activity of MPE
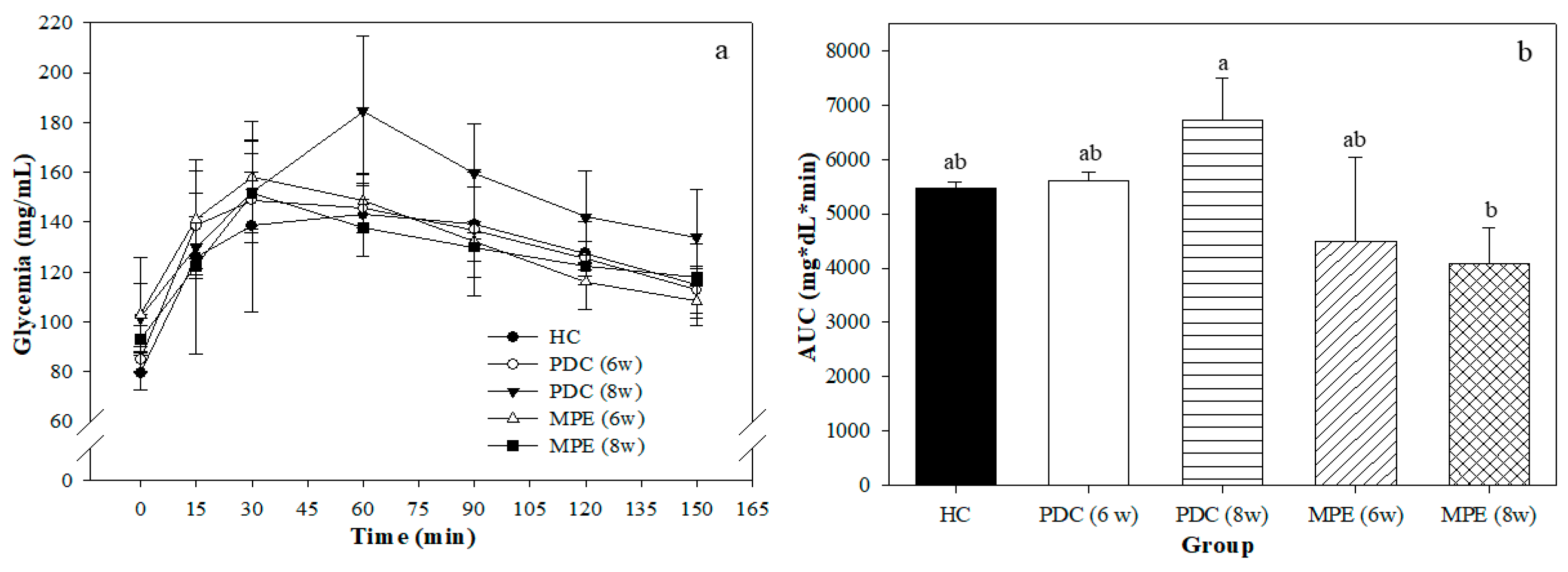
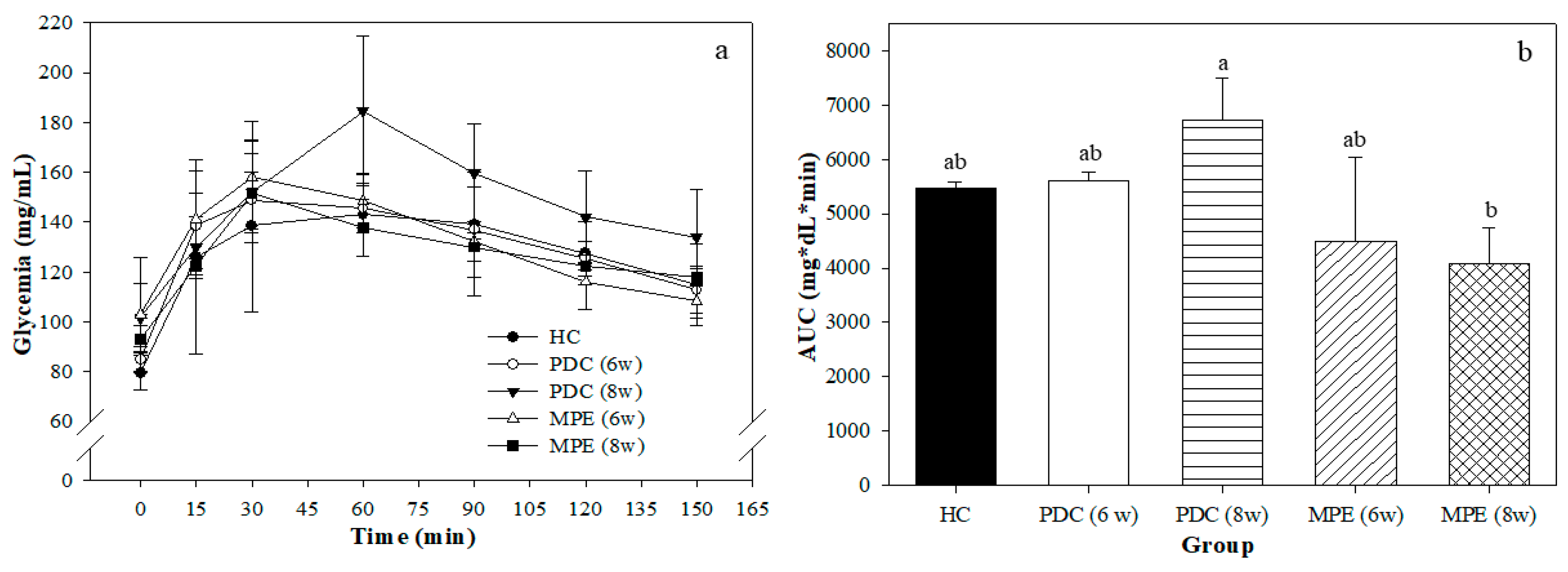
3.3. Effects of MPE in PD Rats
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Association, A.D. Classification and diagnosis of diabetes. Diabetes Care 2017, 40, S11–S24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brannick, B.; Wynn, A.; Dagogo-Jack, S. Prediabetes as a toxic environment for the initiation of microvascular and macrovascular complications. Exp. Biol. Med. 2016, 241, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Organization, W.H. Definition and diagnosis of diabetes mellitus and intermediate hyperglycaemia: Report of a WHO/IDF consultation. 2006, 1, 1–46. Available online: https://apps.who.int/iris/handle/10665/43588 (accessed on 15 March 2022).
- Tabák, A.G.; Herder, C.; Rathmann, W.; Brunner, E.J.; Kivimäki, M. Prediabetes: A high-risk state for developing diabetes. Lancet 2012, 379, 2279. [Google Scholar] [CrossRef] [Green Version]
- Perreault, L.; Temprosa, M.; Mather, K.; Horton, E.; Kitabchi, A.; Larkin, M.; Montez, M.; Thayer, D.; Orchard, T.; Hamman, R. Diabetes Prevention Program Research Group Regression from prediabetes to normal glucose regulation is associated with reduction in cardiovascular risk: Results from the diabetes prevention program outcomes study. Diabetes Care 2014, 37, 2622–2631. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.H. Dietary bioactive compounds and their health implications. J. Food Sci. 2013, 78, A18–A25. [Google Scholar] [CrossRef]
- Ali Asgar, M. Anti-diabetic potential of phenolic compounds: A review. Int. J. Food Prop. 2013, 16, 91–103. [Google Scholar] [CrossRef]
- Zaidan, U.H.; Zen, N.I.M.; Amran, N.A.; Shamsi, S.; Abd Gani, S.S. Biochemical evaluation of phenolic compounds and steviol glycoside from Stevia rebaudiana extracts associated with in vitro antidiabetic potential. Biocatal. Agric. Biotechnol. 2019, 18, 101049. [Google Scholar] [CrossRef]
- Beidokhti, M.N.; Jäger, A.K. Review of antidiabetic fruits, vegetables, beverages, oils and spices commonly consumed in the diet. J. Ethnopharmacol. 2017, 201, 26–41. [Google Scholar] [CrossRef]
- de Bem, G.F.; Costa, C.A.; Santos, I.B.; Cristino Cordeiro, V.d.S.; de Carvalho, L.C.R.M.; de Souza, M.A.V.; Soares, R.d.A.; Sousa, P.J.d.C.; Ognibene, D.T.; Resende, A.C. Antidiabetic effect of Euterpe oleracea Mart.(açaí) extract and exercise training on high-fat diet and streptozotocin-induced diabetic rats: A positive interaction. PLoS ONE 2018, 13, e0199207. [Google Scholar] [CrossRef]
- Domínguez Avila, J.A.; Rodrigo García, J.; González Aguilar, G.A.; De la Rosa, L.A. The antidiabetic mechanisms of polyphenols related to increased glucagon-like peptide-1 (GLP1) and insulin signaling. Molecules 2017, 22, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guasch-Ferré, M.; Merino, J.; Sun, Q.; Fitó, M.; Salas-Salvadó, J. Dietary polyphenols, Mediterranean diet, prediabetes, and type 2 diabetes: A narrative review of the evidence. Oxidative Med. Cell. Longev. 2017, 2017, 6723931. [Google Scholar] [CrossRef]
- Javidi, A.; Mozaffari-Khosravi, H.; Nadjarzadeh, A.; Dehghani, A.; Eftekhari, M.H. The effect of flaxseed powder on insulin resistance indices and blood pressure in prediabetic individuals: A randomized controlled clinical trial. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2016, 21, 70. [Google Scholar]
- Trojan-Rodrigues, M.; Alves, T.; Soares, G.; Ritter, M. Plants used as antidiabetics in popular medicine in Rio Grande do Sul, southern Brazil. J. Ethnopharmacol. 2012, 139, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Ordaz, R.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; González-Aguilar, G.A. Intestinal Permeability and Cellular Antioxidant Activity of Phenolic Compounds from Mango (Mangifera indica cv. Ataulfo) Peels. Int. J. Mol. Sci. 2018, 19, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahurul, M.H.A.; Zaidul, I.S.M.; Ghafoor, K.; Al-Juhaimi, F.Y.; Nyam, K.-L.; Norulaini, N.A.N.; Sahena, F.; Mohd Omar, A.K. Mango (Mangifera indica L.) by-products and their valuable components: A review. Food Chem. 2015, 183, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Yi, X.; Zhang, S.; Cheng, J.; Wang, Y.; Liu, C.; He, X. Bioactive phenolics from mango leaves (Mangifera indica L.). Ind. Crops Prod. 2018, 111, 400–406. [Google Scholar] [CrossRef]
- Gondi, M.; Basha, S.A.; Bhaskar, J.J.; Salimath, P.V.; Prasada Rao, U.J. Anti-diabetic effect of dietary mango (Mangifera indica L.) peel in streptozotocin-induced diabetic rats. J. Sci. Food Agric. 2015, 95, 991–999. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Yahia, E.; Islas-Osuna, M.; Gutierrez-Martinez, P.; Robles-Sánchez, M.; González-Aguilar, G. Effect of ripeness stage of mango fruit (Mangifera indica L., cv. Ataulfo) on physiological parameters and antioxidant activity. Sci. Hortic. 2012, 135, 7–13. [Google Scholar] [CrossRef]
- Quirós-Sauceda, A.E.; Ayala-Zavala, J.F.; Sáyago-Ayerdi, S.G.; Vélez-de La Rocha, R.; Sañudo-Barajas, A.; González-Aguilar, G.A. Added dietary fiber reduces the antioxidant capacity of phenolic compounds extracted from tropical fruit. J. Appl. Bot. Food Qual. 2014, 87, 227–233. [Google Scholar]
- Quirós-Sauceda, A.E.; Chen, C.-Y.O.; Blumberg, J.B.; Astiazaran-Garcia, H.; Wall-Medrano, A.; González-Aguilar, G.A. Processing ‘ataulfo’mango into juice preserves the bioavailability and antioxidant capacity of its phenolic compounds. Nutrients 2017, 9, 1082. [Google Scholar] [CrossRef]
- Velderrain-Rodríguez, G.; Torres-Moreno, H.; Villegas-Ochoa, M.A.; Ayala-Zavala, J.F.; Robles-Zepeda, R.E.; Wall-Medrano, A.; González-Aguilar, G.A. Gallic acid content and an antioxidant mechanism are responsible for the antiproliferative activity of ‘Ataulfo’ mango peel on LS180 cells. Molecules 2018, 23, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Jácquez, V.; Valenzuela-Vázquez, U.; Grajales-Hernández, D.A.; Mateos-Díaz, J.C.; Arrellano-Plaza, M.; Jara-Marini, M.E.; Asaff-Torres, A. Pilot-Scale Integrated Membrane System for the Separation and Concentration of Compounds of Industrial Interest from Tortilla Industry Wastewater (Nejayote). Waste Biomass Valorization 2022, 13, 345–360. [Google Scholar] [CrossRef]
- Ren, S.; Li, K.; Liu, Z. Research on the influences of five food-borne polyphenols on in vitro slow starch digestion and the mechanism of action. J. Agric. Food Chem. 2019, 67, 8617–8625. [Google Scholar] [CrossRef]
- Macho-González, A.; Garcimartín, A.; López-Oliva, M.E.; Ruiz-Roso, B.; Martín de la Torre, I.; Bastida, S.; Benedí, J.; Sánchez-Muniz, F.J. Can carob-fruit-extract-enriched meat improve the lipoprotein profile, VLDL-oxidation, and LDL receptor levels induced by an atherogenic diet in STZ-NAD-diabetic rats? Nutrients 2019, 11, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gondi, M.; Prasada Rao, U. Ethanol extract of mango (Mangifera indica L.) peel inhibits α-amylase and α-glucosidase activities, and ameliorates diabetes related biochemical parameters in streptozotocin (STZ)-induced diabetic rats. J. Food Sci. Technol. 2015, 52, 7883–7893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Shen, Y.; Qiao, Z.; Wang, Y.; Zhang, P.; Yu, B. Comparison of diabetes remission and micronutrient deficiency in a mildly obese diabetic rat model undergoing SADI-S versus RYGB. Obes. Surg. 2019, 29, 1174–1184. [Google Scholar] [CrossRef] [PubMed]
- Nurdiana, S.; Goh, Y.M.; Ahmad, H.; Dom, S.M.; Syimal’ain Azmi, N.; Noor Mohamad Zin, N.S.; Ebrahimi, M. Changes in pancreatic histology, insulin secretion and oxidative status in diabetic rats following treatment with Ficus deltoidea and vitexin. BMC Complement. Altern. Med. 2017, 17, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barman, S.; Srinivasan, K. Zinc supplementation alleviates hyperglycemia and associated metabolic abnormalities in streptozotocin-induced diabetic rats. Can. J. Physiol. Pharmacol. 2016, 94, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Nawaz, H.; Pohorly, J.; Mittal, G.; Kakuda, Y.; Jiang, Y. Extraction of polyphenolics from plant material for functional foods—Engineering and technology. Food Rev. Int. 2005, 21, 139–166. [Google Scholar] [CrossRef]
- Ruiz-Montañez, G.; Ragazzo-Sánchez, J.; Calderón-Santoyo, M.; Velazquez-De La Cruz, G.; De León, J.R.; Navarro-Ocaña, A. Evaluation of extraction methods for preparative scale obtention of mangiferin and lupeol from mango peels (Mangifera indica L.). Food Chem. 2014, 159, 267–272. [Google Scholar] [CrossRef]
- Dineshkumar, B.; Mitra, A.; Manjunatha, M. Studies on the anti-diabetic and hypolipidemic potentials of mangiferin (xanthone glucoside) in streptozotocin-induced type 1 and type 2 diabetic model rats. Int. J. Adv. Pharm. Sci. 2010, 1, 75–85. [Google Scholar]
- Kumar, B.D.; Krishnakumar, K.; Jaganathan, S.K.; Mandal, M. Effect of mangiferin and mahanimbine on glucose utilization in 3T3-L1 cells. Pharmacogn. Mag. 2013, 9, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Chen, S.; Chen, H.; Wang, Y.; Wang, Y.; Hochstetter, D.; Xu, P. Studies on the bioactivity of aqueous extract of pu-erh tea and its fractions: In vitro antioxidant activity and α-glycosidase inhibitory property, and their effect on postprandial hyperglycemia in diabetic mice. Food Chem. Toxicol. 2013, 53, 75–83. [Google Scholar] [CrossRef]
- Irondi, E.A.; Oboh, G.; Akindahunsi, A.A.; Boligon, A.A.; Athayde, M.L. Phenolic composition and inhibitory activity of Mangifera indica and Mucuna urens seeds extracts against key enzymes linked to the pathology and complications of type 2 diabetes. Asian Pac. J. Trop. Biomed. 2014, 4, 903–910. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Yao, F.; Du, J.; Deng, X.; Li, C. Persimmon tannin decreased the glycemic response through decreasing the digestibility of starch and inhibiting α-amylase, α-glucosidase, and intestinal glucose uptake. J. Agric. Food Chem. 2018, 66, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Haque, A.R.; Kabir, M.; Hasan, M.; Khushe, K.J.; Hasan, S. Fruit by-products: The potential natural sources of antioxidants and α-glucosidase inhibitors. J. Food Sci. Technol. 2021, 58, 1715–1726. [Google Scholar] [CrossRef]
- Sellamuthu, P.S.; Arulselvan, P.; Muniappan, B.P.; Fakurazi, S.; Kandasamy, M. Influence of mangiferin on membrane bound phosphatases and lysosomal hydrolases in streptozotocin induced diabetic rats. Lat. Am. J. Pharm. 2012, 31, 1013–1020. [Google Scholar]
- Zhao, C.; Yang, C.; Wai, S.T.C.; Zhang, Y.; Portillo, M.P.; Paoli, P.; Wu, Y.; San Cheang, W.; Liu, B.; Carpéné, C. Regulation of glucose metabolism by bioactive phytochemicals for the management of type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2019, 59, 830–847. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Prajapati, A. Quercetin in anti-diabetic research and strategies for improved quercetin bioavailability using polymer-based carriers—A review. RSC Adv. 2015, 5, 97547–97562. [Google Scholar] [CrossRef]
- Anhê, G.F.; Okamoto, M.M.; Kinote, A.; Sollon, C.; Lellis-Santos, C.; Anhê, F.F.; Lima, G.A.; Hirabara, S.M.; Velloso, L.A.; Bordin, S. Quercetin decreases inflammatory response and increases insulin action in skeletal muscle of ob/ob mice and in L6 myotubes. Eur. J. Pharmacol. 2012, 689, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Prasad, C.V.; Anjana, T.; Banerji, A.; Gopalakrishnapillai, A. Gallic acid induces GLUT4 translocation and glucose uptake activity in 3T3-L1 cells. FEBS Lett. 2010, 584, 531–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latha, R.C.R.; Daisy, P. Insulin-secretagogue, antihyperlipidemic and other protective effects of gallic acid isolated from Terminalia bellerica Roxb. in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2011, 189, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Cázares-Camacho, R.; Domínguez-Avila, J.A.; Astiazarán-García, H.; Montiel-Herrera, M.; González-Aguilar, G.A. Neuroprotective effects of mango cv. ‘Ataulfo’ peel and pulp against oxidative stress in streptozotocin-induced diabetic rats. J. Sci. Food Agric. 2021, 101, 497–504. [Google Scholar] [CrossRef]
- Morsi, R.M.; El-Tahan, N.; El-Hadad, A.M. Effect of aqueous extract Mangifera indica leaves, as functional foods. J. Appl. Sci. Res. 2010, 6, 712–721. [Google Scholar]
- Dra, L.A.; Sellami, S.; Rais, H.; Aziz, F.; Aghraz, A.; Bekkouche, K.; Markouk, M.; Larhsini, M. Antidiabetic potential of Caralluma europaea against alloxan-induced diabetes in mice. Saudi J. Biol. Sci. 2019, 26, 1171–1178. [Google Scholar] [CrossRef]
- Anhê, F.F.; Desjardins, Y.; Pilon, G.; Dudonné, S.; Genovese, M.I.; Lajolo, F.M.; Marette, A. Polyphenols and type 2 diabetes: A prospective review. PharmaNutrition 2013, 1, 105–114. [Google Scholar] [CrossRef]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef]
- Safdar, M.N.; Kausar, T.; Nadeem, M.; Jabbar, S.; Ahmed, S.; Taj, T.; Tufail, F.; Mehmood, W. Cardioprotective Effect of Mango and Kinnow Peel Extracts on Doxorubicin-induced Cardiotoxicity in Albino Rats: Cardioprotective Effect of Mango and Kinnow Peel Extracts on Doxorubicin. Proc. Pak. Acad. Sci. B Life Environ. Sci. 2017, 54, 219–235. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Parameter | HC | PDC (6 w) | PDC (8 w) | MPE (6 w) | MPE (8 w) |
|---|---|---|---|---|---|
| Food intake (g·d−1) | 23.2 ± 0.7 a | 14.6 ± 2.2 b | 14.6 ± 1.4 b | 17.8 ± 3.0 b | 17.5 ± 2.6 b |
| Body weight (g) | 344.6 ± 20.2 a | 267.4 ± 33.2 b | 308.0 ± 64.3 a | 326.4 ± 10.7 a | 299.0 ± 18.8 a |
| FG (mg·dL−1)Basal | 82.0 ± 2.6 b | 91.8 ± 9.0 ab | 111.8 ± 12.3 a | 90.0 ± 9.2 ab | 84.8 ± 2.2 b |
| FG (mg·dL−1)End | 83.8 ± 3.1 b | 84.6 ± 4.6 b | 114.4 ± 13.5 a | 89.2 ± 9.6 ab | 94.0 ± 5.2 ab |
| FI (µUI·mL−1)End | 14.2 ± 2.6 b | 18.5 ± 2.6 ab | 19.1 ± 6.1 a | 14.5 ± 3.0 b | 15.8 ± 2.0 b |
| HOMA-IR | 3.2 ± 0.7 b | 4.2 ± 0.5 ab | 5.8 ± 2.0 a | 3.4 ± 0.6 b | 4.0 ± 0.3 b |
| HOMA-β | 185.2 ± 15.8 b | 242.9 ± 64.8 a | 118.0 ± 43.3 c | 174.2 ± 71.2 bc | 152.6 ± 43.1 bc |
| QUICKI | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.0 |
| TC (mg·dL−1) | 64.0 ± 13.1 c | 81.9 ± 11.1 b | 96.7 ± 10.8 a | 67.0 ± 4.0 c | 76.6 ± 5.0 b |
| HDL-c (mg·dL−1) | 11.8 ± 1.1 a | 7.6 ± 0.7 bc | 5.5 ± 1.2 c | 8.5 ± 0.6 b | 8.0 ± 0.3 b |
| LDL-c (mg·dL−1) | 21.5 ± 13.0 c | 46.9 ± 7.5 b | 63.1 ± 8.9 a | 42.1 ± 7.1 bc | 50.5 ± 4.4 ab |
| VLDL-c (mg·dL−1) | 17.1 ± 4.1 b | 27.5 ± 5.6 a | 28.2 ± 3.6 a | 16.4 ± 5.1 b | 18.1 ± 1.0 b |
| TAG (mg·dL−1) | 86.7 ± 12.0 b | 137.2 ± 22.7 a | 140.8 ± 14.3 a | 81.8 ± 6.7 b | 90.6 ± 5.5 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preciado-Saldaña, A.M.; Domínguez-Avila, J.A.; Ayala-Zavala, J.F.; Astiazaran-Garcia, H.F.; Montiel-Herrera, M.; Villegas-Ochoa, M.A.; González-Aguilar, G.A.; Wall-Medrano, A. Mango “Ataulfo” Peel Extract Improves Metabolic Dysregulation in Prediabetic Wistar Rats. Life 2022, 12, 532. https://doi.org/10.3390/life12040532
Preciado-Saldaña AM, Domínguez-Avila JA, Ayala-Zavala JF, Astiazaran-Garcia HF, Montiel-Herrera M, Villegas-Ochoa MA, González-Aguilar GA, Wall-Medrano A. Mango “Ataulfo” Peel Extract Improves Metabolic Dysregulation in Prediabetic Wistar Rats. Life. 2022; 12(4):532. https://doi.org/10.3390/life12040532
Chicago/Turabian StylePreciado-Saldaña, Alejandra M., Jesús Abraham Domínguez-Avila, Jesús Fernando Ayala-Zavala, Humberto F. Astiazaran-Garcia, Marcelino Montiel-Herrera, Mónica A. Villegas-Ochoa, Gustavo A. González-Aguilar, and Abraham Wall-Medrano. 2022. "Mango “Ataulfo” Peel Extract Improves Metabolic Dysregulation in Prediabetic Wistar Rats" Life 12, no. 4: 532. https://doi.org/10.3390/life12040532
APA StylePreciado-Saldaña, A. M., Domínguez-Avila, J. A., Ayala-Zavala, J. F., Astiazaran-Garcia, H. F., Montiel-Herrera, M., Villegas-Ochoa, M. A., González-Aguilar, G. A., & Wall-Medrano, A. (2022). Mango “Ataulfo” Peel Extract Improves Metabolic Dysregulation in Prediabetic Wistar Rats. Life, 12(4), 532. https://doi.org/10.3390/life12040532