Abstract
Canine distemper is a highly contagious viral disease in carnivores and represents a serious threat for both wild and domestic animals. The aim of our study was to monitor the occurrence of the canine distemper virus in wildlife from the Czech Republic, reveal the H gene heterogeneity in positive samples and perform subsequent phylogenetic analysis. In total, 412 wild animals of 10 species were included in the study: 219 red foxes (Vulpes vulpes), 79 European badgers (Meles meles), 47 European otters (Lutra lutra), 40 stone martens (Martes foina), 10 pine martens (M. martes), 7 raccoons (Procyon lotor), 5 undetermined martens (Martes sp.), 2 wolves (Canis lupus), 1 European polecat (Mustela putorius), 1 free-ranging ferret (Mustela putorius furo), and 1 free-ranging American mink (Neovison vison). Most animals were found dead or were killed by hunters during hunting seasons in the years 2012–2020 and came from all 14 regions of the Czech Republic. In the animals that were hunted, symptoms such as apathy, loss of shyness or disorientation were reported. Canine distemper virus (CDV) was detected by real-time RT-PCR in the tissues of 74 (18%) of the animals, including 62 (28%) red foxes, 4 (10%) stone martens, 3 (43%) raccoons, 2 (20%) pine martens, 2 (2.5%) European badgers and 1 (20%) undetermined marten. There was a statistical difference in positivity among animal species (p < 0.0001), regions (p = 0.0057), and the years of sampling (p = 0.0005). To determine the genetic characteristics of circulating variants of CDV in wildlife, 23 of 74 CDV variants were partially sequenced. Phylogenetic analysis showed that 21 variants belonged to the European lineage and two strains belonged to the European-Wildlife lineage. This study provides the first comprehensive overview of the prevalence and spatial distribution of CDV in wildlife in the Czech Republic, including molecular phylogenetic analysis of currently circulating CDV lineages.
1. Introduction
Canine distemper is a disease of dogs and wild carnivores caused by the canine morbillivirus (previously called distemper virus, CDV), belonging to the genus Morbillivirus, family Paramyxoviridae [1]. Closely related paramyxoviruses, such as the phocine morbillivirus (phocid distemper virus), rinderpest virus in animals and measles virus in humans are important pathogens [2]. Canine distemper virus is spread worldwide and is associated with highly contagious infection in susceptible species. The main hosts of the virus are carnivores, especially canids [3], in which the infection is clinically manifested with respiratory, gastrointestinal, dermatologic, and neurological disorders [4]. The CDV may also infect animals from the families Ailuridae, Felidae, Hyaenidae, Mephitidae, Mustelidae, Odobenidae, Otariidae, Phocidae, Procyonidae, Ursidae and Viveridae of the order Carnivora and in animals from species of the orders Artiodactyla, Primates, Proboscidea, Rodentia and recently Pilosa [4,5].
The CDV virion contains a 15,690 bp negative-sense single-stranded RNA genome with the hemagglutinin (H) gene as one of the most variable genes [6]. The H gene is therefore a suitable target for molecular epidemiological studies [7,8]. Based on phylogenetic analysis of H gene sequences, CDV is classified into 19 genetic lineages distributed worldwide including America-1 (vaccine strains), America-2 through -5, Europe (also called Europe/South America-1), Europe-Wildlife, Arctic-like, America-1/Europe, Rockborn-like, Asia-1 through -4, India-1/Asia-5, Asia-6, South Africa, and South America-2 and -3 [9,10,11,12].
In Europe, CDV circulates mainly in the wild red fox population. Canine distemper was previously described in northern Italy and in nearby Germany (the federal states of Bavaria, Saxony, and Anhalt) [13,14,15]. In the Czech Republic, only sporadic cases of CDV have been proven in wild mustelids [16]. The last wave of canine distemper attacked the domestic dog population in the Czech Republic between the years 1990 and 1993 [17], however a massive vaccination of domestic dogs prevented the spread of the virus.
The aims of our study were to assess the prevalence of the CDV in wildlife from the Czech Republic and to use phylogenetic analysis of H gene sequence to determine their evolutionary relationships with the European lineages.
2. Results
In total, 74 (18%) of 412 wild animals were positive for the CDV, including 62 (28%) foxes, 4 (10%) stone martens, 2 (20%) pine martens, 2 (2.5%) European badgers, 2 (43%) raccoons and 1 (20%) undetermined marten; animals belonging to the other species were negative (Table 1). There was a statistical difference in positivity among species (p < 0.0001). Animal species that had only one representative were excluded from statistical analyses. Scheffé’s method of multiple comparisons showed that at the significance level of 0.05 the pairs (red fox, European badger) and (red fox, European otter) differed. Positive animals were found in 12 of 14 regions with statistical differences among regions (p = 0.0057); the highest prevalence 37.5% was in the region Ústí nad Labem. Depending on the year, the prevalence ranged from 41.6% in 2012 to 0% in 2014 with significant statistical differences (p = 0.0005).

Table 1.
Characteristics of 412 wild animals tested for canine distemper virus.
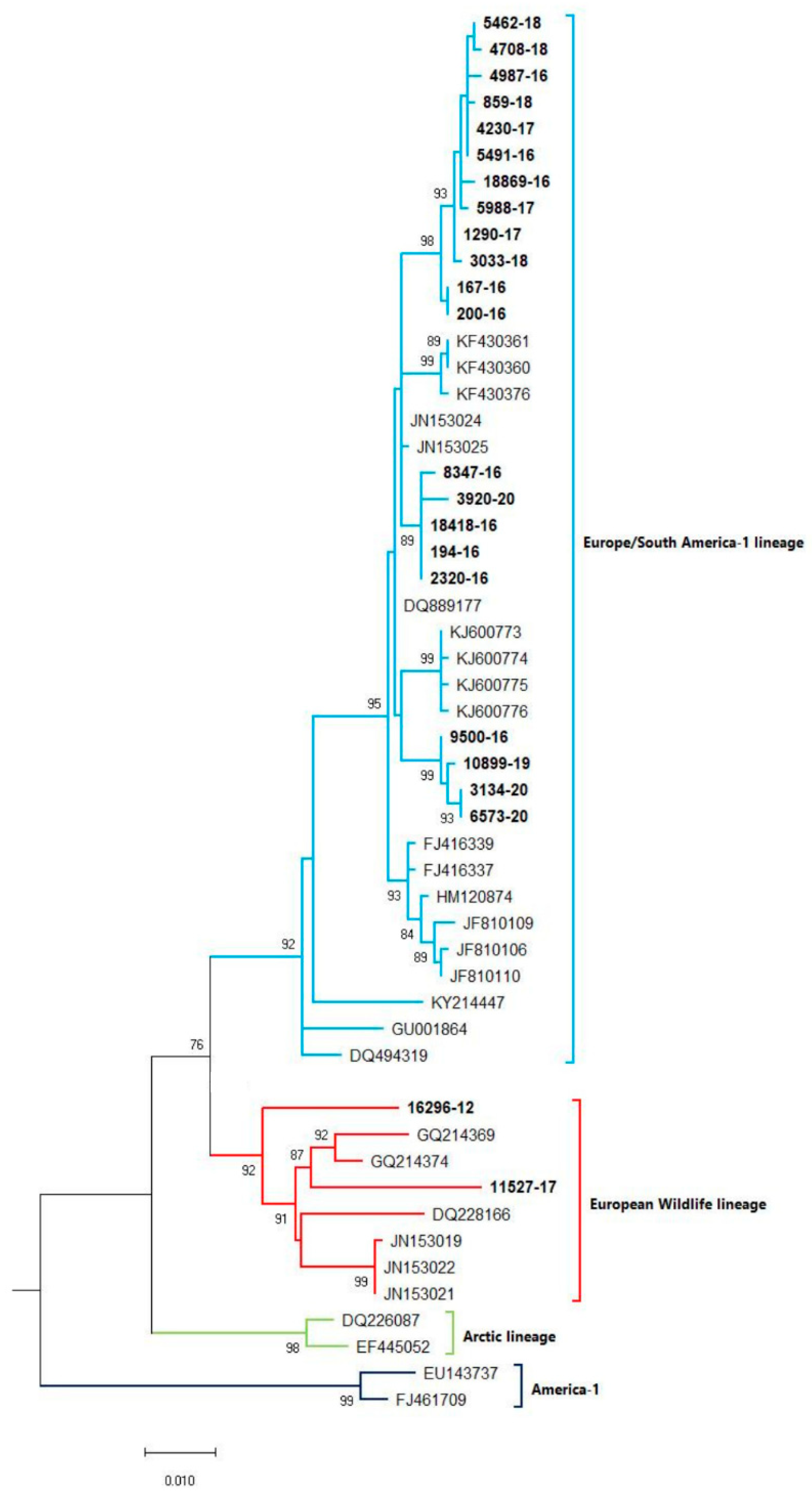
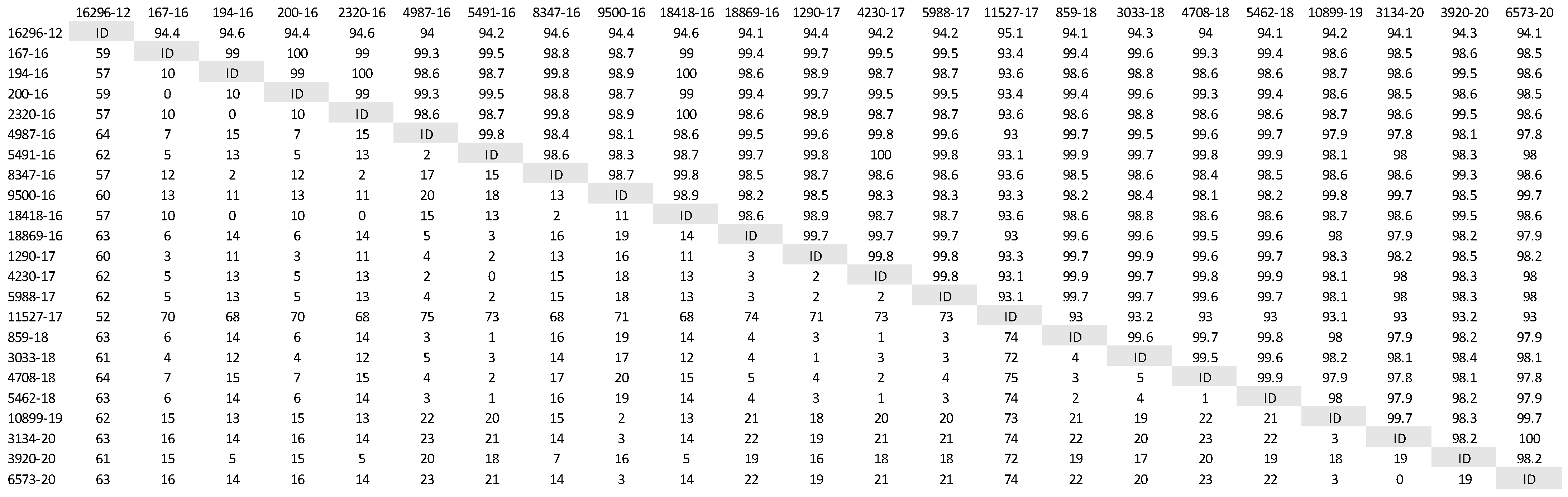
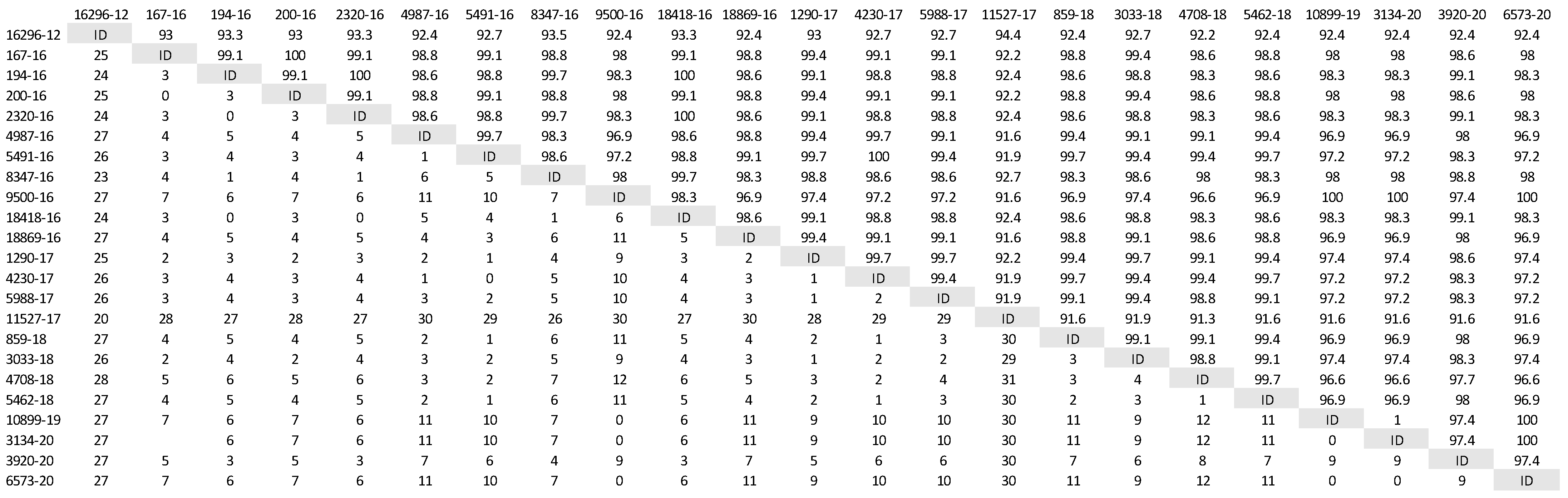
A phylogenetic tree was constructed (Figure 1) based on partial H gene sequences from 23 animals that were positive for CDV (Table 2). Twenty-one variants were assigned to the Europe/South America -1 lineage, suggesting that this lineage is the most prevalent among wild animals in the Czech Republic. Two other variants isolated from the stone marten (11527/17) and the pine marten (16296/12) were characterized as European Wildlife lineage. The genetic relatedness of CDV variants circulating in wildlife population in the Czech Republic with other European variants of CDV is shown in nucleotide (Figure 2) and amino acids variability matrices (Figure 3). The nucleotide sequence differences within 21 Europe/South America -1 lineage reached up to 23 nucleotides, which represents 97.8% similarity. The difference in 23 nucleotides led to the substitution of 12 amino acids, which means that approximately half of the substitutions were non-synonymous. Two variants (no. 9500/16 from a marten and no. 10899/16 from a fox) from the Pilsen region showed high identities of 99.44% and 99.22%, respectively, with the CDV variants isolated from dogs in Hungary during the years 2005–2006 (GenBank accession no. DQ889177). The rest of the 19 variants showed high identities of 98.80–99.73% with the CDV variants isolated from the two foxes in Germany in 2008 (GenBank accession no. JN153024.1 and JN153025.1). Two variants characterized as European Wildlife lineage showed a difference in 52 nucleotides, which represent substitutions in 20 amino acids. The variants no. 16296/12 and 11527/17, isolated from martens, showed a high identity of 97.01% with the CDV variant isolated from the badger from Austria (GenBank accession no. GQ214374.2). These results suggest cross-border transmission of the CDV between the Czech Republic and neighboring countries.
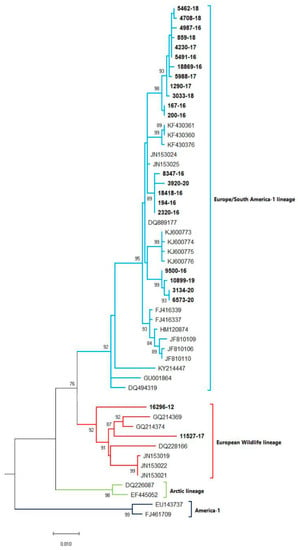
Figure 1.
Phylogenetic tree constructed from the Czech CDV (canine distemper virus) variants isolated from wild animals and variants, representing different CDV lineages, available from the NCBI. The sequences from our study are in bold. The evolutionary history was inferred by using the Maximum Likelihood method and Tamura 3-parameter model [18]. The percentage of trees, in which the associated taxa clustered together, is shown next to the branches (bootstrap values 1000 replicants). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Evolutionary analyses were conducted in MEGA X [19].
Table 2.
Characteristics of 23 samples, randomly selected from 74 samples positive for canine distemper virus, used for partial H gene sequencing to characterize CDV lineages circulating among wildlife in the Czech Republic.
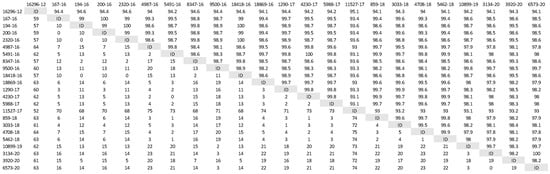
Figure 2.
Nucleotide variability of Czech CDV (canine distemper virus) variants isolated from wild animals. The matrix was generated by plotting the CDV variants against each other. The difference in the number of nucleotides is expressed below the diagonal, the nucleotide similarity of the CDV variants in percentages is expressed above the diagonal.
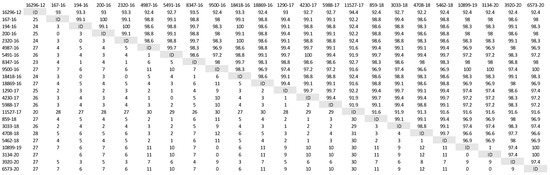
Figure 3.
Amino acids variability of Czech CDV (canine distemper virus) variants isolated from wild animals. The matrix was generated by plotting the CDV variants against each other. The difference in the number of amino acids is expressed below the diagonal, the amino acids similarity of the CDV variants in percentages is expressed above the diagonal.
3. Discussion
Canine distemper disease was reported in Europe in the second half of the 18th century [20]. In the Czech Republic, the virus was first reported in the 1930s and in the following decades caused the deaths of domestic dogs and farm silver foxes (Vulpes vulpes), polar foxes (Vulpes lagopus), and American minks (Neovison vison) [21]. The last outbreak of canine distemper disease in domestic dogs in the Czech Republic occurred between the years 1990 and 1993. At that time, it affected about 40% of the population of dogs [17]. In the case of foxes, great attention was focused on rabies, that is why there was not any monitoring of the CDV in foxes in the past. There is only one article published on canine distemper in wild mustelids in the Czech Republic [16]. Authors collected 194 samples of brain tissues from wild mustelids in the years 2001–2003 and tested them for the presence of CDV by a direct immunofluorescence test. They detected CDV in stone martens and badgers and observed clinical signs of distemper in three stone marten pup siblings.
The widespread vaccination of dogs in the first half of the 1990s prevented the spread of CDV and is probably the reason for the eradication of infection in dogs in the Czech Republic [17]. Although several fatal cases of post-vaccination canine distemper were demonstrated in the years 2002–2004, e.g., in domestic ferrets (Mustela putorius furo) bred as pets and in the South American coati (Nasua nasua) at a zoo (unpublished results), no autochthonous clinical case of distemper in dogs has been published in the last 25 years. Due to the absence of clinical cases in dogs, the occurrence of canine distemper has not been systematically monitored even in red foxes and other wild carnivores in the Czech Republic.
In our study, we reported CDV predominantly in red foxes, less in both species of martens, badgers, and raccoons, while the virus was not detected in other species. The composition of the species investigated was influenced by the population density of carnivores in the Czech Republic [22]. The red fox has been overpopulated in the Czech Republic for a long time, and it has been regularly hunted and examined, mainly to prevent the spread of rabies (the last case was proven in the Czech Republic in 2002). It appears likely that the red fox plays a major role in the circulation of the CDV in Central Europe. Our results are consistent with previous studies in neighboring Germany [23,24]. The population of stone martens is more numerous than the pine marten population, however both are only rarely hunted and examined in the laboratory. The badger is rarely hunted in the Czech Republic and examined for trichinellosis. The wolf is a rare and protected species in the Czech Republic, but has been increasing in numbers in recent years. Similarly, the European otter is a protected species with an estimated population size of 2500. Raccoons and the American mink are invasive species in the Czech countryside, both of which are occasionally hunted and sporadically examined [22].
In our study, we focused on the long-term collection of samples. In total, 23 partially sequenced variants of the CDV were characterized with respect to their lineages, diversity, the geographical origin, and year of collection. We were limited to the passive monitoring of the CDV in the wild carnivore population (just sick or dead carnivores). However, the results showed that during the period observed (2012–2020), CDV infection showed some dynamism. A total of 47 CDV variants were detected in animals from the Pilsen region, which was also a region where the infection was detected both at the beginning (2012) and at the end (2020) of our study. The Pilsen region is the western region of the Czech Republic, which is directly adjacent to Bavaria (Germany), where the occurrence of the CDV in wild carnivores has been described shortly before [13].
It is probable that the situation in the Czech Republic was like that in Germany, with the difference that the CDV in foxes in the Czech Republic escaped the attention of professional observers for a long time. One of the reasons for a such speculation may be the fact that rabies was a more significant and monitored disease until around 2005. The red fox is an adaptable species that has virtually no enemy in the Czech Republic, and therefore, as a successful species, its numbers have increased for several decades. Rabies has been involved to some extent in regulating the numbers of the fox population in the 1990s. After oral vaccination, the fox population recovered, and a gradual increase in the density of fox populations in the Czech Republic continued. The reluctance of hunters to hunt foxes has also contributed to this increase. In a large population of foxes, there is a higher frequency of contacts and therefore easier transmission of infection. The CDV circulated in wild carnivores in varying degrees in different regions of the Czech Republic, as evidenced in mustelids [16].
The sequence analysis of 23 CDV variants, randomly selected from the various animal species, regions, and years, showed little genetic variability (heterogeneity) with a predominant lineage Europe/South America-1 in 21 variants. Only two variants were characterized as European Wildlife lineage. The occurrence of these lineages was previously published in wild animals from neighboring countries Austria and Germany [25,26]. The CDV variants selected for sequencing analyses were highly identical to the CDV variants from Germany, Austria, and Hungary [27,28,29]. Similar CDV variants, circulating in wildlife in Central Europe, confirm the cross-border transmission of the CDV between Germany, Austria, and the Czech Republic. With regard to this fact, it cannot be ruled out that, in the future, other CDV lineages may appear in the population of Czech wild carnivores.
4. Materials and Methods
4.1. Animals
Samples (brain, lung, and diaphragm) from 412 wild animals representing 10 species were collected in all 14 Czech regions from January 2012 to December 2020 (Table 1). The following species were sampled: 219 red foxes (Vulpes vulpes), 79 European badgers (Meles meles), 47 European otters (Lutra lutra), 40 stone martens (Martes foina), 10 pine martens (Martes martes), 7 raccoons (Procyon lotor), 5 undetermined martens (Martes sp.), 2 wolves (Canis lupus), 1 European polecat (Mustela putorius), 1 free-ranging ferret (Mustela putorius furo), and 1 free-ranging American mink (Neovison vison).
A large number of the animals (foxes and badgers) were hunted during the hunting season and their carcasses were sent for examination to the State Veterinary Institute (SVÚ) Prague, SVÚ Olomouc or SVÚ Jihlava. In the case of overpopulated foxes, hunting is legal throughout the year. Foxes showing symptoms such as apathy, loss of shyness or disorientation are monitored for rabies according to the current regulations. The same animals were tested in our study for the presence of CDV. A smaller group of animals including ferret, mink, otter, polecat, wolf and 23 foxes were found dead. Three animals (1 fox, 1 raccoon, and 1 wolf), found in the wild, showed loss of shyness and impaired orientation and were therefore euthanized.
4.2. Molecular Methods
Canine distemper virus (CDV) was detected in the tissues (brain and lung) of animals by real-time RT-PCR. In the case of badgers, CDV was detected in samples of the diaphragm that was collected by hunters and sent to the SVI Prague for detection of Trichinella spp. Total nucleic acids were extracted from tissues with the MagNA Pure Compact Nucleic Isolation Kit I (Roche, Basel, Switzerland) and machine TissueLyser II (Qiagen), with input sample volumes of 200 µL and elution volumes of 50 µL. Isolated nucleic acids were stored at −80 °C until they were assayed. For the initial detection of the CDV, a real-time polymerase chain reaction (RT-PCR) targeting N-protein was performed according to the procedure described by Elia et al. [28].
Not all samples positive by RT-PCR were suitable for sequencing. Some of them were weakly positive and thus with a small chance to be successfully sequenced and most of the positive samples were collected in the years 2016 and 2017 from a relatively limited area around Pilsen. That is why positive samples from different species, different regions and years were used together with a representative number of samples from the Pilsen region. Partial Sanger sequencing of the hemagglutinin (H) gene was thus carried out in 23 of 74 CDV variants (Table 2).
First, a reverse transcription and amplification of the H gene, described by Harder et al. [29], was performed using the OneStep RT-PCR kit (Qiagen, Hilden, Germany). The final volume of 50 µL of the master mix contained 0.6 µM of each primer (Table 3) and 5 µL of nucleic acid extract. The reaction was performed under the following conditions: 50 °C 30 min, 95 °C 15 min, 35 cycles (94 °C 30 s, 50 °C 30 s and 68 °C 3 min) and 72 °C 10 min. Then, a nested PCR was carried out using the GoTaq Colorless Master Mix (Promega, Madison, WI, USA) with the final volume of 20 µL containing 0.6 µM of each inner primers and 0.4 mM dNTP. Then 1 µL of the PCR product was added. The characteristics of the two sets of inner primers used in the nested PCR [30,31] are summarized in Table 3. The PCR reaction was performed under the following conditions: 95 °C 5 min, 30 cycles (94 °C 30 s, 50 °C 30 s and 72 °C 1 min) and 72 °C 10 min. The sequencing reactions were prepared using the Big Dye Terminator Cycle Sequencing Kit v3.1 (Thermo Fisher Scientific, MA, USA) and evaluated on the 3130 and 3500 Genetic Analyzers (Life Technologies, Carlsbad, CA, USA). Partial nucleotide sequences 1071 bp long were used for phylogenetic analysis. The partial sequences were aligned to 24 hemagglutinin sequences obtained from the GenBank database using MAFFT [32]. A phylogenetic tree was constructed using the MEGA X software [19]. To reveal nucleotide and amino acid differences in the isolates, the partial sequences were compared in a Sequence Identity Matrix and a Sequence Difference Count Matrix using BioEdit Software [33]. These 23 sequences were deposited into the NCBI GenBank under the accession’s numbers MW828731-MW288753 (Table 2).
Table 3.
Characteristics of inner primers used in nested PCR used for the amplification of H genes of the canine distemper virus.
4.3. Statistical Analysis
The results were statistically analyzed with Pearson’s chi-square test for independence using STATISTICA Cz 12 [34]. We tested the null hypothesis that the occurrence of CDV does not differ among species, regions, or years of sampling. The differences were considered statistically significant if the p-value was <0.05. In the case of a statistically significant difference of CDV in some of the variables, Scheffé’s multiple comparison method [34] was subsequently applied. This method was used to identify a statistically significant difference between pairs of species, regions, and years of sampling. Animal species that had only one representative were excluded from statistical analyses.
Author Contributions
Conceptualization, E.K., K.S. and E.B.; methodology, L.Č., A.D.; software, L.Č., M.B.; validation, E.K., L.Č. and A.D.; formal analysis, E.K., L.Č., A.D. and M.B.; investigation, E.K., L.Č.; resources, E.K.; data curation, E.K.; writing—original draft preparation E.K., L.Č. and K.S., writing—review and editing, K.S., E.B.; visualization, E.K., L.Č.; supervision, E.K., K.S. and E.B.; project administration, E.K., E.B.; funding acquisition, K.S., E.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
Acknowledgments
We would like to thank K. Makoň (DES OP, Animal Rescue Station, Plzeň), J. Vadlejch (Czech University of Life Sciences), ALKA Wildlife, Public Service Company, P. Barták (SVI Jihlava), J. Bardoň (SVI Olomouc), and O. Máca (SVI Prague) for providing samples, V. Paurová (University of Veterinary Sciences Brno) for sample preparation and A. Nagy (SVI Prague) for helping with the sequence analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- ICTV, International Committee on Taxonomy of Viruses. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/negative-sense-rna-viruses/w/paramyxoviridae/1183/genus-morbillivirus (accessed on 11 October 2021).
- Quintero-Gil, C.; Rendon-Marin, S.; Martinez-Gutierrez, M.; Ruiz-Saenz, J. Origin of Canine Distemper Virus: Consolidating evidence to understand potential zoonoses. Front. Microbiol. 2019, 10, 1982. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, M.; Ruiz-Saenz, J. Diversity of susceptible hosts in canine distemper virus infection: A systemic review and data synthesis. BMC Vet. Res. 2016, 12, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martella, V.; Elia, G.; Buonavoglia, C. Canine distemper virus. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Granjeiro, M.D.B.; Kavasaki, M.L.; Morgado, T.O.; Pavelegini, L.A.D.; De Barros, M.A.; Fontana, C.; De Assis Bianchini, M.; De Oliveira Souza, A.; Santos, A.R.G.L.O.; Lunardi, M.; et al. First report of a canine morbillivirus infection in a giant anteater (Myrmecophaga tridactyla) in Brazil. Vet. Med. Sci. 2020, 6, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Parardo, I.D.R.; Johnson, G.C.; Kleiboeker, S.B. Phylogenetic characterization of canine distemper viruses detected in naturally infected dogs in North America. J. Clin. Microbiol. 2005, 43, 5009–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Une, Y.; Mochizuki, M. Hemagglutinin genotype profiles of canine distemper virus from domestic dogs in Japan. Arch. Virol. 2001, 146, 149–155. [Google Scholar] [CrossRef]
- Von Messling, V.; Zimmer, G.; Herrler, G.; Haas, L.; Cattaneo, R. The hemagglutinin of canine distemper virus determines tropism and cytopathogenicity. J. Virol. 2001, 75, 6418–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Sabatino, D.; Lorusso, A.; Di Francesco, C.E.; Gentile, L.; Di Pirro, V.; Bellacicco, A.L.; Giovannini, A.; Di Francesco, G.; Marruchella, G.; Marsilio, F.; et al. Arctic-lineage canine distemper virus as a cause of death in Apennine wolves (Canis lupus) in Italy. PLoS ONE 2014, 9, e82356. [Google Scholar] [CrossRef]
- Piewbang, C.H.; Radtanakatikanon, A.; Puenpa, J.; Poovorawan, Y.; Techangamsuwan, S. Genetic and evolutionary analysis of a new Asia-4 lineage and naturally recombinant canine distemper virus strains from Thailand. Sci. Rep. 2019, 9, 3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abirami, M.; Srinivas, M.V.; Vasu, J.; Antony, P.X.; Thanislass, J.; Muthaiah, M.; Mukhopadhyay, H.K. Genotyping of Canine Distemper Virus Lineage in Clinically Infected Dogs in Puducherry, Southern India. Microbiol. Res. J. Int. 2020, 30, 17–30. [Google Scholar] [CrossRef]
- Wang, R.; Wang, X.; Zhai, J.; Zhang, P.; Irwin, D.M.; Shen, X.; Chen, W.; Shen, Y. A new canine distemper virus lineage identified from red pandas in China. Transbound. Emerg. Dis. 2021; epub ahead of print. [Google Scholar]
- Sekulin, K.; Hafner-Marx, A.; Kolodziejek, J.; Janik, D.; Schmidt, P.; Nowotny, N. Emergence of Canine Distemper in Bavarian wildlife associated with a specific amino acid exchange in the haemagglutinin protein. Vet. J. 2011, 187, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Denzin, N.; Herwig, V.; Van Der Grinten, E. Occurrence and geographical distribution of Canine Distemper Virus infection in red foxes (Vulpes vulpes) of Saxony-Anhalt, Germany. Vet. Microbiol. 2013, 162, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Nouvellet, P.; Donnelly, C.A.; De Nardi, M.; Rhodes, C.J.; De Benedictis, P.; Citterio, C.; Obber, F.; Lorenzetto, M.; Pozza, M.D.; Cauchemez, S.; et al. Rabies and canine distemper virus epidemics in the red fox population of northern Italy (2006–2010). PLoS ONE 2013, 8, e61588. [Google Scholar] [CrossRef] [PubMed]
- Pavlacik, L.; Celer, V.; Koubek, P.; Literak, I. Prevalence of canine distemper virus in wild mustelids in the Czech Republic and a case of canine distemper in young stone martens. Vet. Med. 2007, 52, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Svoboda, M.; Srenk, P. Distemper. In Infectious Diseases of Dogs and Cats; Svoboda, M., Pospíšil, Z., Eds.; Czech Association of Veterinarians of Small Animals: Brno, Czech, 1996; pp. 123–135. (In Czech) [Google Scholar]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Uhl, E.W.; Kelderhouse, C.H.; Buikstra, J.; Blick, J.P.; Bolon, B.; Hogan, R.J. New world origin of canine distemper: Interdisciplinary insights. Int. J. Paleopathol. 2019, 24, 266–278. [Google Scholar] [CrossRef]
- Konrád, J. Nemoci kožešinových zvířat (Diseases of Furred Animals); Státní zemědělské nakladatelství: Praha, Czech, 1989; p. 368. (In Czech) [Google Scholar]
- Anděra, M.; Červený, J. Velcí savci v České republice. Rozšíření, historie a Ochrana. 2. Šelmy (Carnivora) [Large Mammals in the Czech Republic. Distribution, History and Protection. 2. Carnivores (Carnivora)]; Národní museum: Praha, Czech, 2009; p. 215. (In Czech) [Google Scholar]
- Frölich, K.; Czupalla, O.; Haas, L.; Hentschke, J.; Dedek, J.; Fickel, J. Epizootiological investigations of canine distemper virus in free-ranging carnivores from Germany. Vet. Microbiol. 2000, 74, 283–292. [Google Scholar] [CrossRef]
- Lempp, C.; Jungwirth, N.; Grilo, M.L.; Reckendorf, A.; Ulrich, A.; Van Neer, A.; Bodewes, R.; Pfankuche, V.M.; Bauer, C.; Osterhaus, A.D.; et al. Pathological findings in the red fox (Vulpes vulpes), stone marten (Martes foina) and raccoon dog (Nyctereutes procyonoides), with special emphasis on infectious and zoonotic agents in Northern Germany. PLoS ONE 2017, 12, e0175469. [Google Scholar] [CrossRef]
- Benetka, V.; Leschnik, M.; Affenzeller, N.; Möstl, K. Phylogenetic analysis of Austrian canine distemper virus strains from clinical samples from dogs and wild carnivores. Vet. Rec. 2011, 168, 377. [Google Scholar] [CrossRef]
- Nikolin, V.M.; Wibbelt, G.; Michler, F.U.; Wolf, P.; East, M.L. Susceptibility of carnivore hosts to strains of canine distemper virus from distinct genetic lineages. Vet. Microbiol. 2012, 156, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Demeter, Z.; Lakatos, B.; Palade, E.A.; Kozma, T.; Forgách, P.; Rusvai, M. Genetic diversity of Hungarian canine distemper virus strains. Vet. Microbiol. 2007, 122, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Elia, G.; Decaro, N.; Martella, V.; Cirone, F.; Lucente, M.S.; Lorusso, E.; Di Trani, L.; Buonavoglia, C. Detection of canine distemper virus in dogs by real-time RT-PCR. J. Virol. Methods 2006, 136, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Harder, T.C.; Kenter, M.; Vos, H.; Siebelink, K.; Huisman, W.; Van Amerongen, G.; Orvell, C.; Barrett, T.; Appel, M.J.; Osterhaus, A.D. Canine distemper virus from diseased large felids: Biological properties and phylogenetic relationships. J. Gen. Virol. 1996, 77, 397–405. [Google Scholar] [CrossRef] [PubMed]
- An, D.J.; Yoon, S.H.; Park, J.Y.; No, Y.S.; Park, B.K. Phylogenetic characterization of canine distemper virus isolates from naturally infected dogs and a marten in Korea. Vet. Microbiol. 2008, 132, 389–395. [Google Scholar] [CrossRef] [PubMed]
- da Fontoura Budaszewski, F.; Pinto, L.D.; Weber, M.N.; Caldart, E.T.; Alves, C.D.; Martella, V.; Ikuta, N.; Lunge, V.R.; Canal, C.W. Genotyping of canine distemper virus strains circulating in Brazil from 2008 to 2012. Virus Res. 2014, 13, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment bases on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Statsoft, Inc. STATISTICA (Data Analysis Software System), Version 12, 2013. Available online: www.statsoft.com (accessed on 26 August 2021).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).