
Blue Light-Induced Retinal Neuronal Injury and Amelioration by Commercially Available Blue Light-Blocking Lenses

 ,
,
Abstract
:1. Introduction
2. Methodology
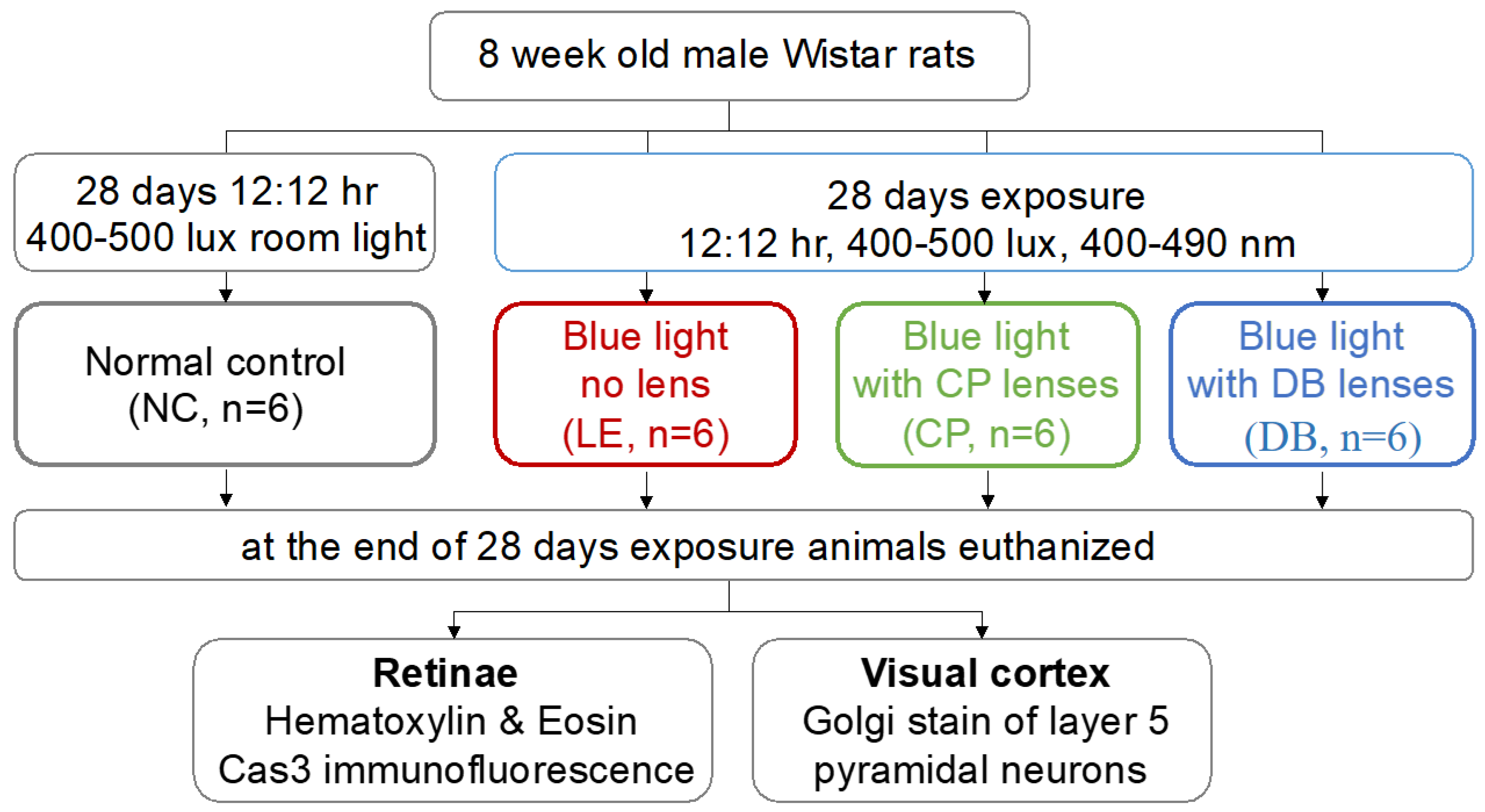
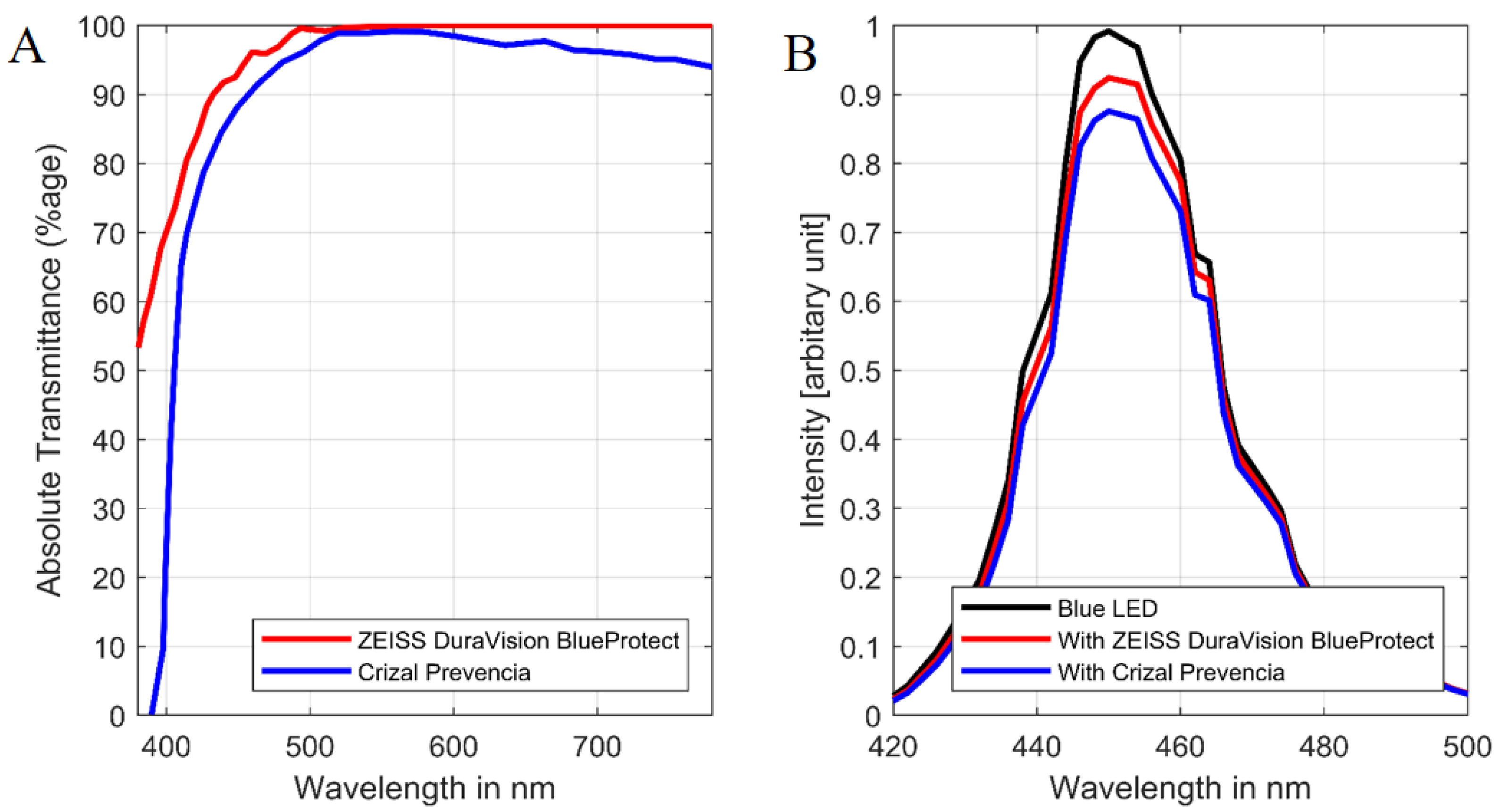
2.1. Experimental Setup and Animal Resources
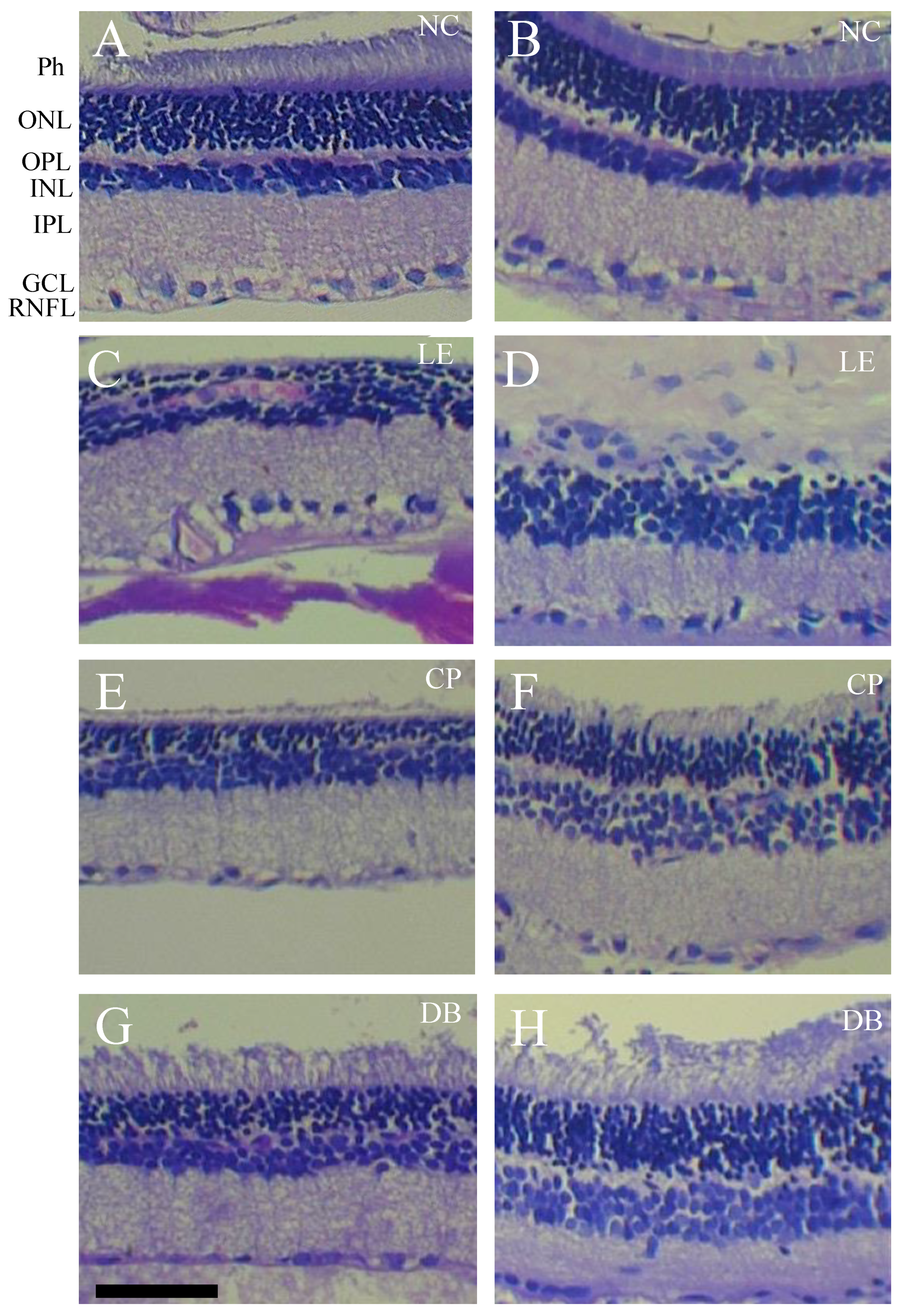
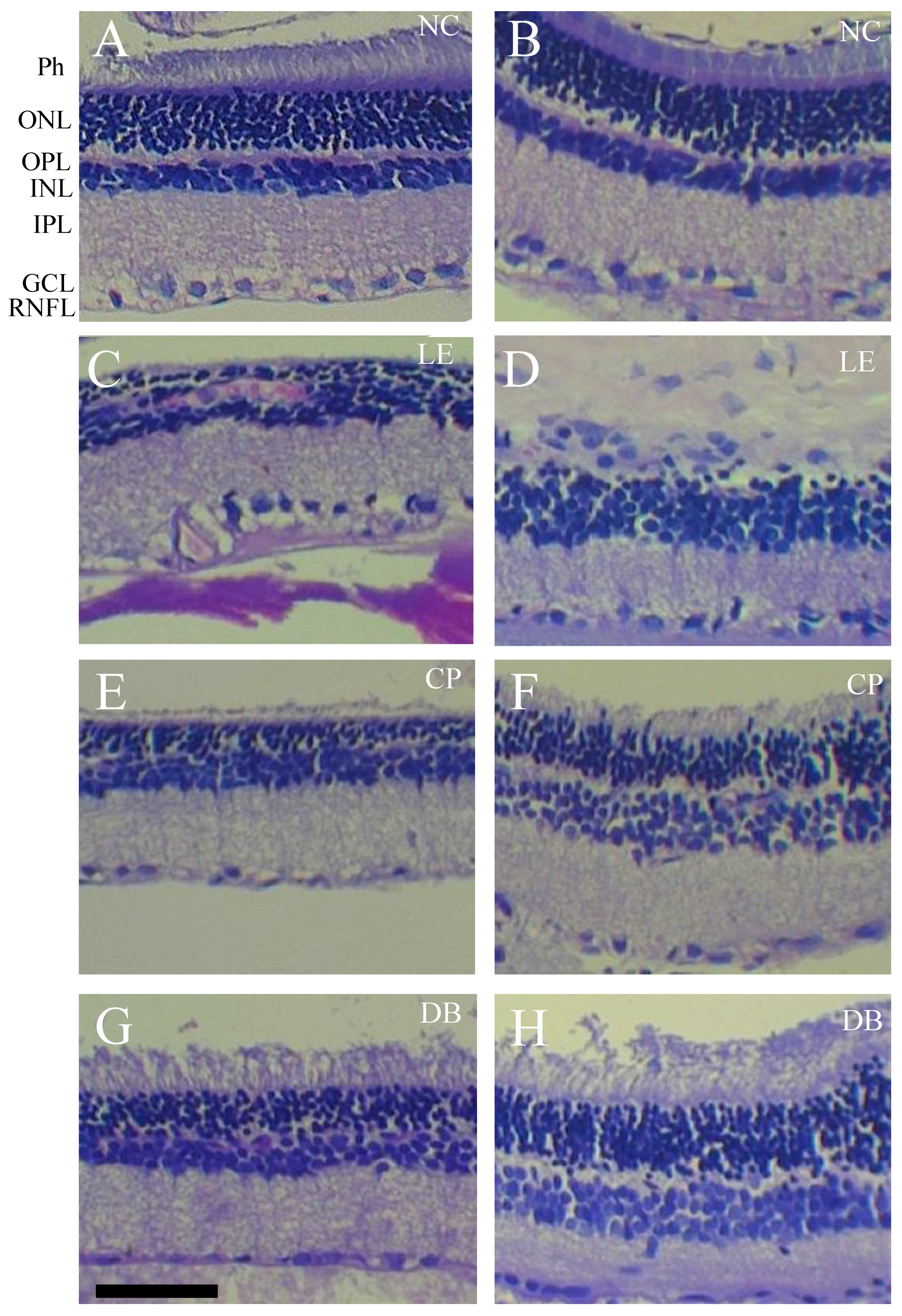
2.2. Histology
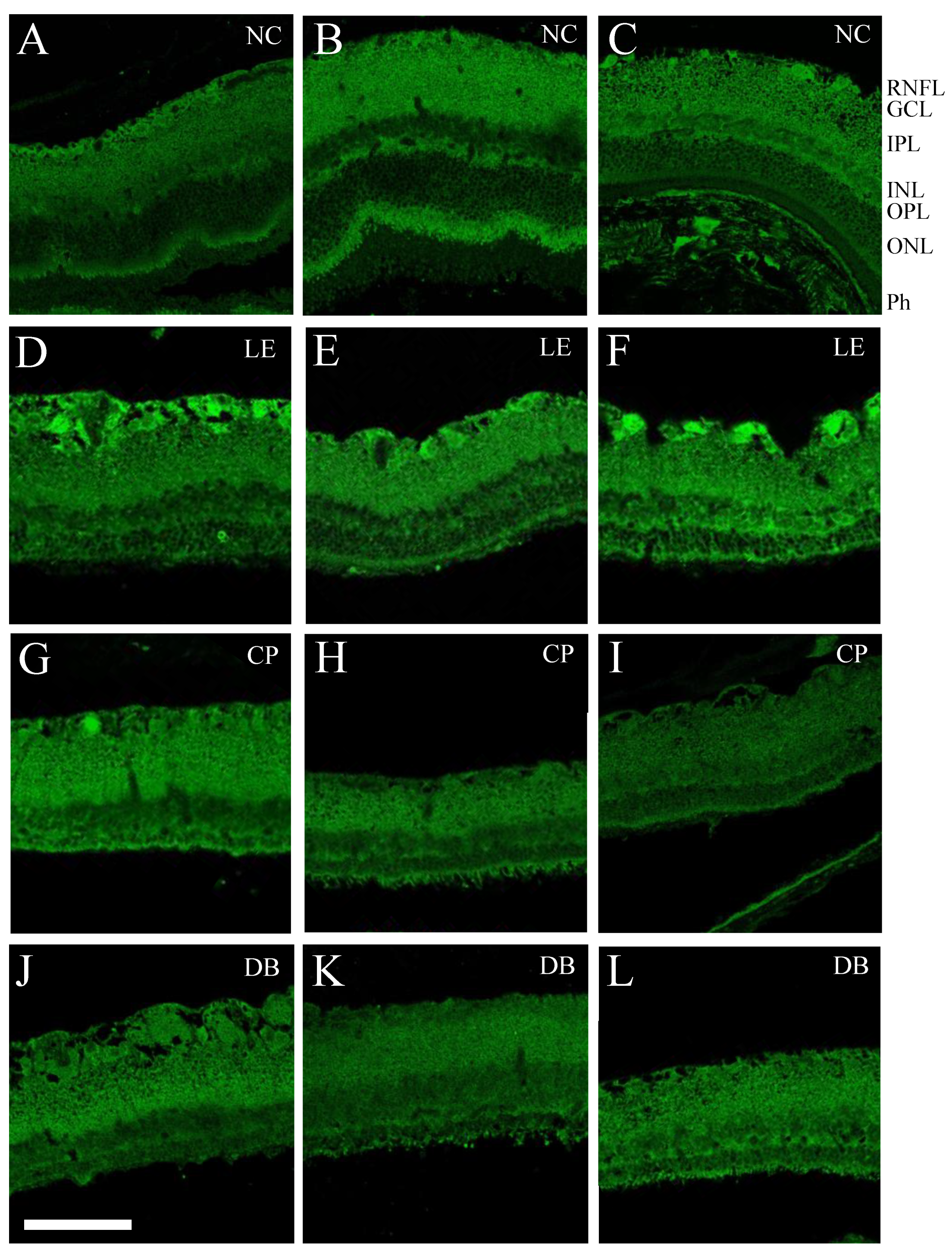
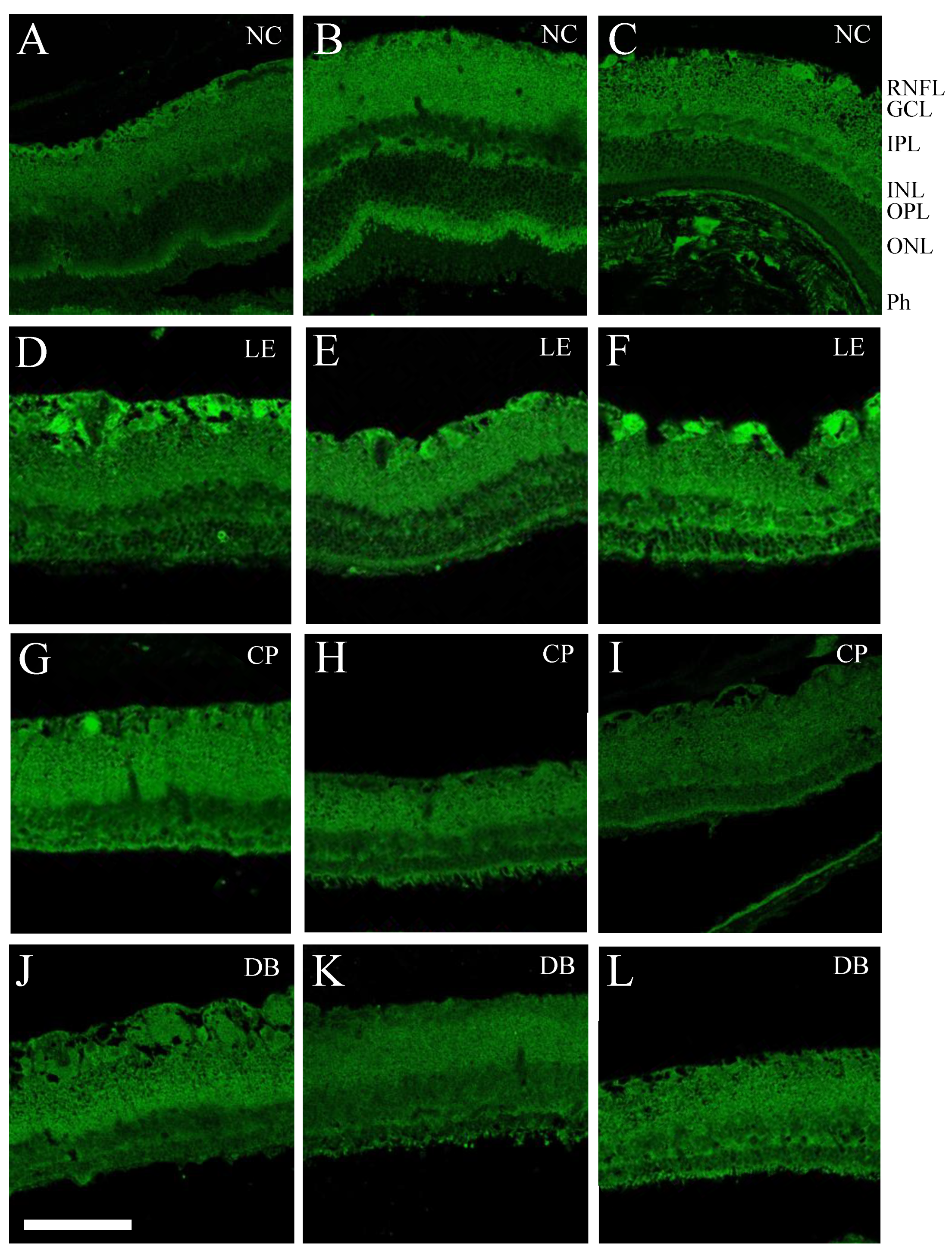
2.3. Immunofluorescence
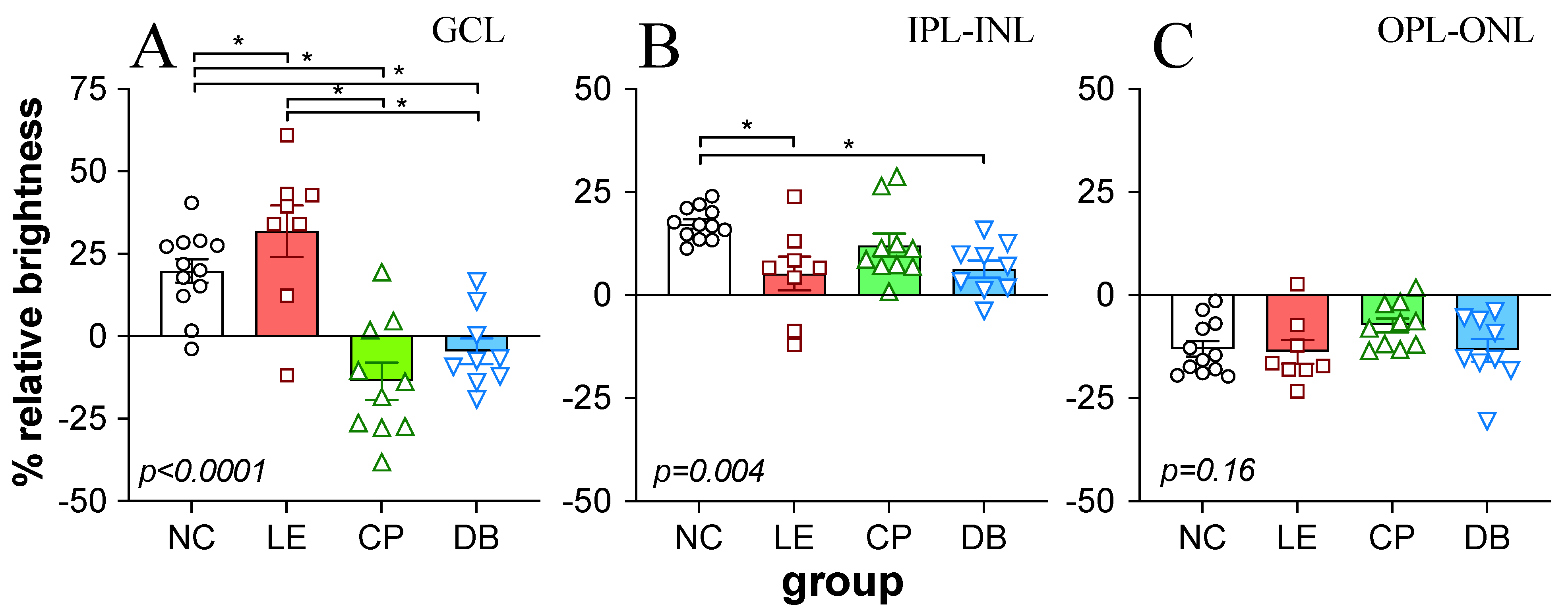
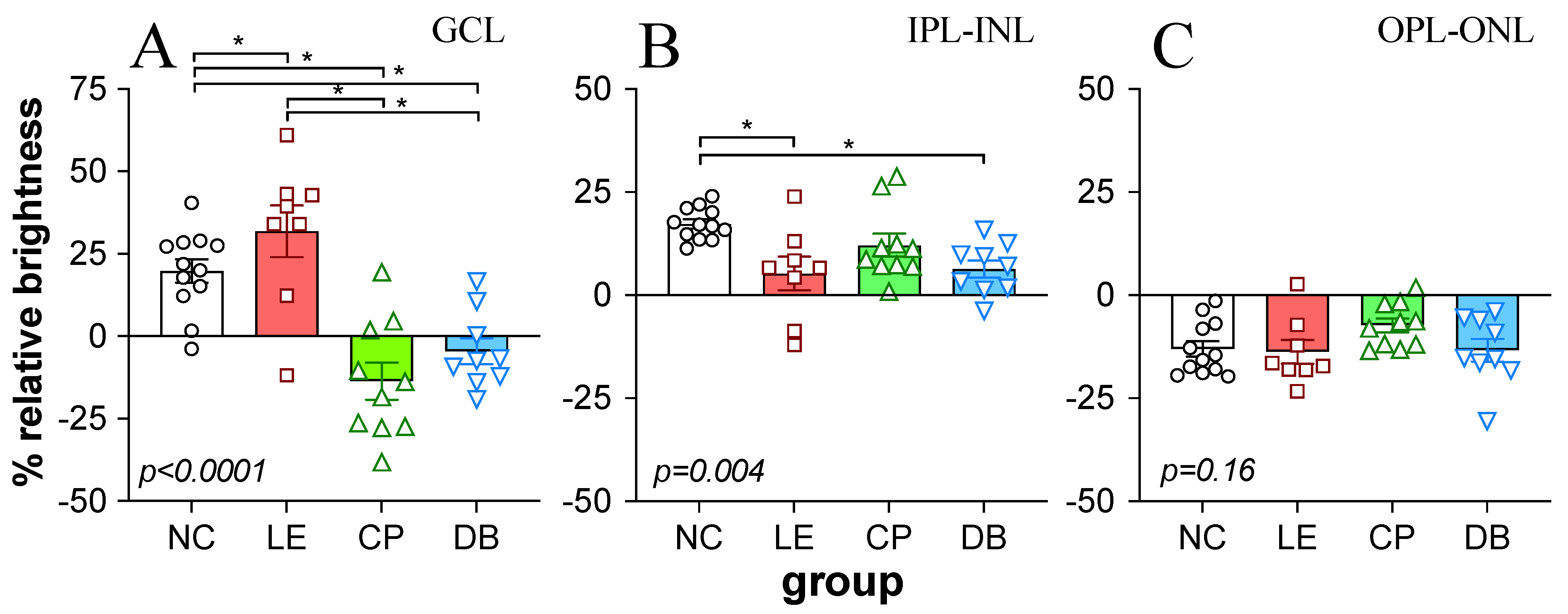
2.4. Retinal Immunofluorescence Imaging and Quantification
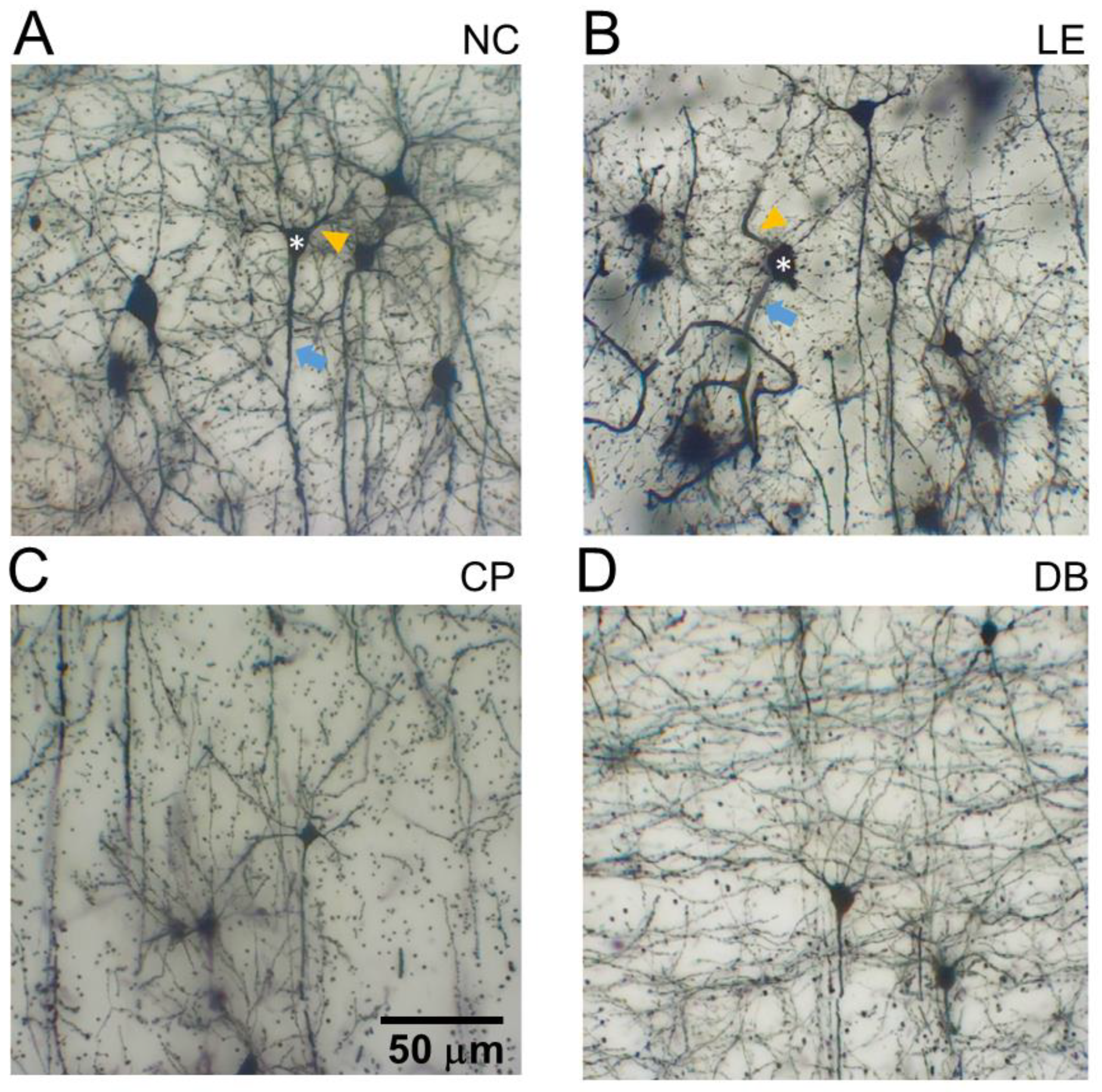
2.5. Golgi–Cox Stains for Visual Cortex Layer 5 Pyramidal Neurons (V1-L5PN)
2.6. V1-L5PN Image Capture Dendritic Quantification
2.7. Statistical Analysis
3. Results
3.1. Retinal Histology
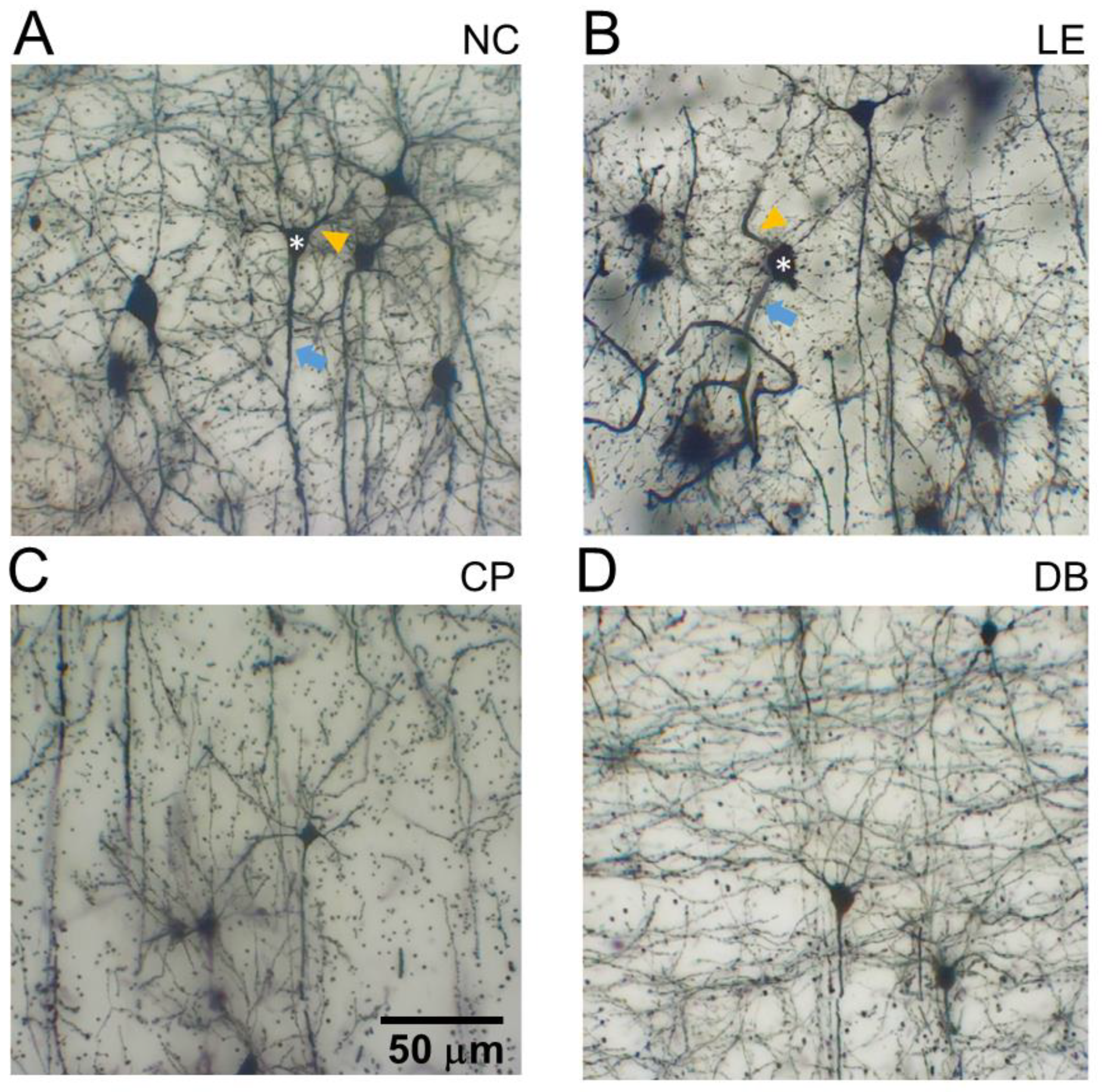
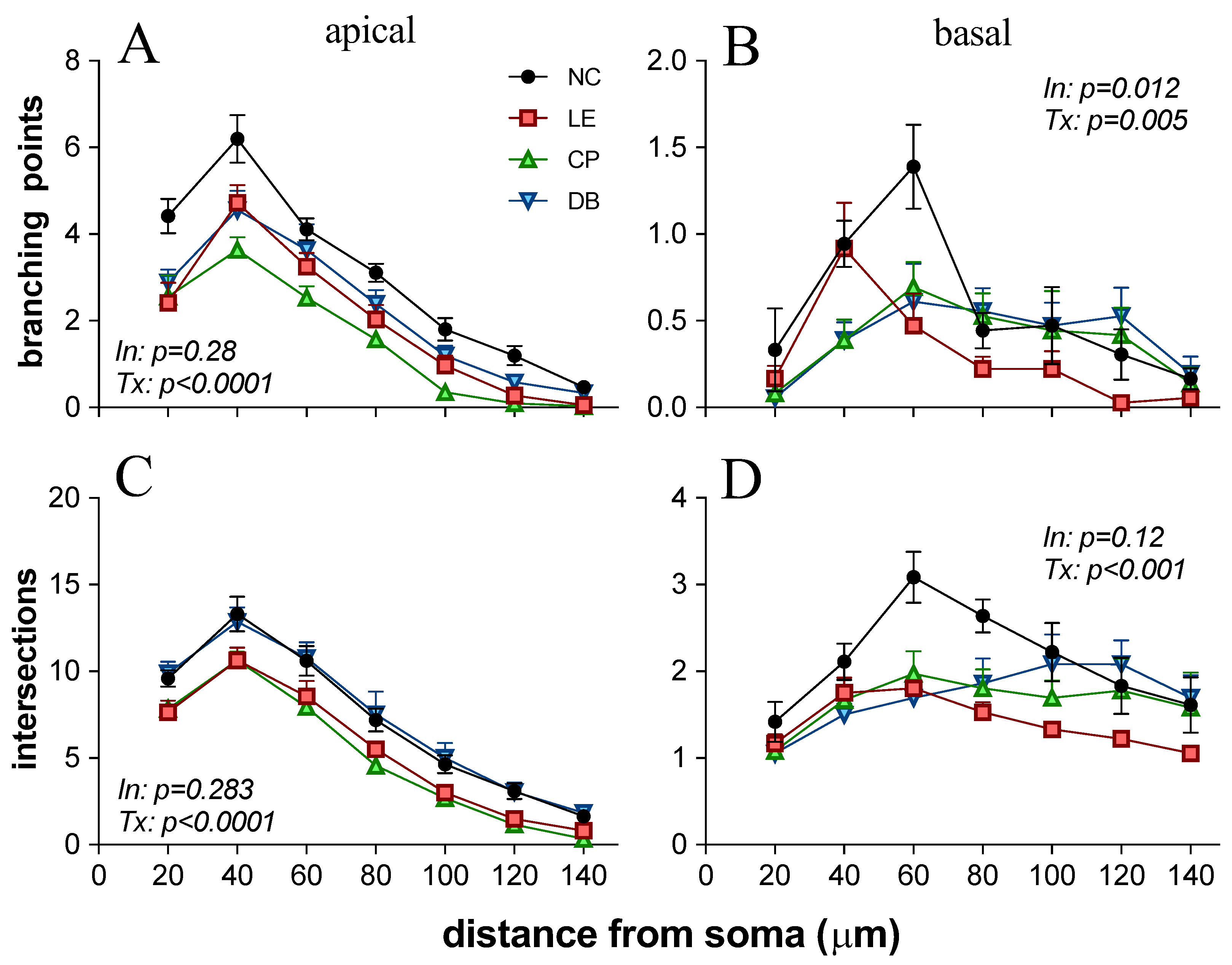
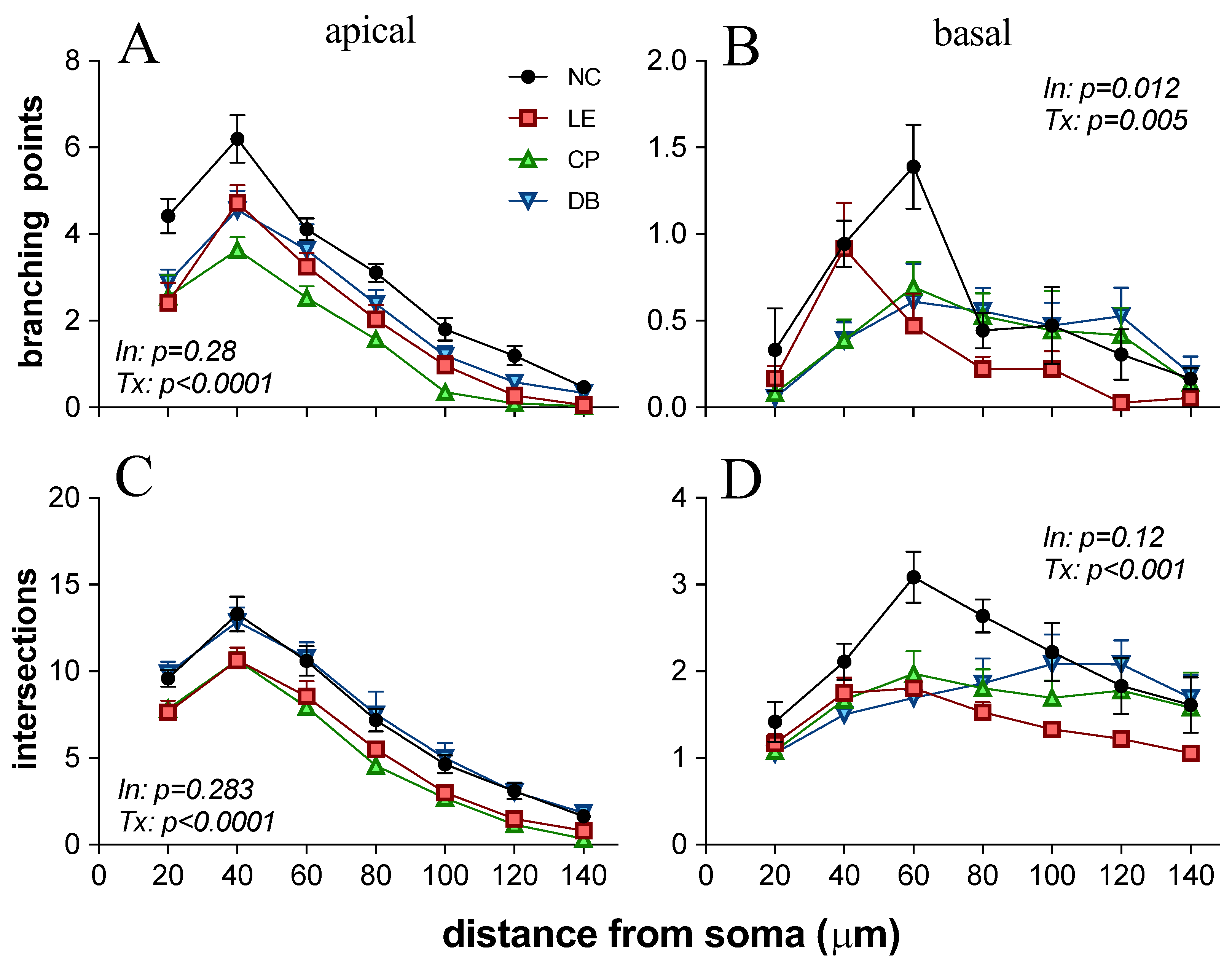
3.2. Blue Light-Induced Changes to Visual Cortex Neurons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Behar-Cohen, F.; Martinsons, C.; Viénot, F.; Zissis, G.; Barlier-Salsi, A.; Cesarini, J.P.; Enouf, O.; Garcia, M.; Picaud, S.; Attia, D. Light-emitting diodes (LED) for domestic lighting: Any risks for the eye? Prog. Retin. Eye Res. 2011, 30, 239–257. [Google Scholar] [CrossRef]
- Sheppard, A.L.; Wolffsohn, J.S. Digital eye strain: Prevalence, measurement and amelioration. BMJ Open Ophthalmol. 2018, 3, e000146. [Google Scholar] [CrossRef] [Green Version]
- Lawrenson, J.G.; Hull, C.C.; Downie, L.E. The effect of blue-light blocking spectacle lenses on visual performance, macular health and the sleep-wake cycle: A systematic review of the literature. Ophthalmic Physiol. Opt. 2017, 37, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiromoto, K.; Kuse, Y.; Tsuruma, K.; Tadokoro, N.; Kaneko, N.; Shimazawa, M.; Hara, H. Colored lenses suppress blue light-emitting diode light-induced damage in photoreceptor-derived cells. J. Biomed. Opt. 2016, 21, 035004. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, J.B.; Khazova, M.; Price, L.L.A. Low-energy light bulbs, computers, tablets and the blue light hazard. Eye 2016, 30, 230–233. [Google Scholar] [CrossRef] [Green Version]
- Delcourt, C.; Carrière, I.; Ponton-Sanchez, A.; Fourrey, S.; Lacroux, A.; Papoz, L. POLA Study Group Light exposure and the risk of age-related macular degeneration: The Pathologies Oculaires Liées à l’Age (POLA) study. Arch. Ophthalmol. 2001, 119, 1463–1468. [Google Scholar] [CrossRef]
- Ham, W.T.; Ruffolo, J.J.; Mueller, H.A.; Clarke, A.M.; Moon, M.E. Histologic analysis of photochemical lesions produced in rhesus retina by short-wave-length light. Invest. Ophthalmol. Vis. Sci. 1978, 17, 1029–1035. [Google Scholar]
- Wen, R.; Cheng, T.; Song, Y.; Matthes, M.T.; Yasumura, D.; LaVail, M.M.; Steinberg, R.H. Continuous exposure to bright light upregulates bFGF and CNTF expression in the rat retina. Curr. Eye Res. 1998, 17, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Yun, J.; Yoon, Y.D.; Park, S.I.; Seo, Y.J.; Park, W.S.; Chu, H.Y.; Park, K.H.; Lee, M.Y.; Lee, C.W.; et al. Blue light effect on retinal pigment epithelial cells by display devices. Integr. Biol. 2017, 9, 436–443. [Google Scholar] [CrossRef] [PubMed]
- van Norren, D.; Gorgels, T.G.M.F. The Action Spectrum of Photochemical Damage to the Retina: A Review of Monochromatic Threshold Data. Photochem. Photobiol. 2011, 87, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Van Norren, D.; Schellekens, P. Blue light hazard in rat. Vision Res. 1990, 30, 1517–1520. [Google Scholar] [CrossRef]
- Grimm, C.; Wenzel, A.; Williams, T.P.; Rol, P.O.; Hafezi, F.; Reme, C.E. Rhodopsin-Mediated Blue-Light Damage to the Rat Retina: Effect of Photoreversal of Bleaching. Investig. Ophthalmol. Vis. Sci. 2001, 42, 497–505. [Google Scholar]
- Contín, M.A.; Arietti, M.M.; Benedetto, M.M.; Bussi, C.; Guido, M.E. Photoreceptor damage induced by low-intensity light: Model of retinal degeneration in mammals. Mol. Vis. 2013, 19, 1614–1625. [Google Scholar] [PubMed]
- Shang, Y.M.; Wang, G.S.; Sliney, D.H.; Yang, C.H.; Lee, L.L. Light-emitting-diode induced retinal damage and its wavelength dependency in vivo. Int. J. Ophthalmol. 2017, 10, 191–202. [Google Scholar] [CrossRef]
- Hatori, M.; Gronfier, C.; Van Gelder, R.N.; Bernstein, P.S.; Carreras, J.; Panda, S.; Marks, F.; Sliney, D.; Hunt, C.E.; Hirota, T.; et al. Global rise of potential health hazards caused by blue light-induced circadian disruption in modern aging societies. NPJ Aging Mech. Dis. 2017, 3, 5–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, K.N.; Saafir, T.B.; Tosini, G. The role of retinal photoreceptors in the regulation of circadian rhythms. Rev. Endocr. Metab. Disord. 2009, 10, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, A.; Nordin, S.; Heiden, M.; Sandström, M. Symptoms, personality traits, and stress in people with mobile phone-related symptoms and electromagnetic hypersensitivity. J. Psychosom. Res. 2010, 68, 37–55. [Google Scholar] [CrossRef]
- Ostrin, L.A.; Abbott, K.S.; Queener, H.M. Attenuation of short wavelengths alters sleep and the ipRGC pupil response. Ophthalmic Physiol. Opt. 2017, 37, 440–450. [Google Scholar] [CrossRef]
- Weng, S.; Estevez, M.E.; Berson, D.M.; Barnes, S. Mouse Ganglion-Cell Photoreceptors Are Driven by the Most Sensitive Rod Pathway and by Both Types of Cones. PLoS ONE 2013, 8, e66480. [Google Scholar] [CrossRef]
- Tosini, G.; Ferguson, I.; Tsubota, K. Effects of blue light on the circadian system and eye physiology. Mol. Vis. 2016, 22, 61–72. [Google Scholar]
- Bauer, M.; Glenn, T.; Monteith, S.; Gottlieb, J.F.; Ritter, P.S.; Geddes, J.; Whybrow, P.C. The potential influence of LED lighting on mental illness. World J. Biol. Psychiatry 2018, 19, 59–73. [Google Scholar] [CrossRef] [PubMed]
- West, K.E.; Jablonski, M.R.; Warfield, B.; Cecil, K.S.; James, M.; Ayers, M.A.; Maida, J.; Bowen, C.; Sliney, D.H.; Rollag, M.D.; et al. Blue light from light-emitting diodes elicits a dose-dependent suppression of melatonin in humans. J. Appl. Physiol. 2011, 110, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2015, 112, 1232–1237. [Google Scholar] [CrossRef] [Green Version]
- Prayag, A.S.; Najjar, R.P.; Gronfier, C. Melatonin suppression is exquisitely sensitive to light and primarily driven by melanopsin in humans. J. Pineal Res. 2019, 66. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, J.R.; Miller, A.S.; Zhou, J. Blue light-absorbing intraocular lens and retinal pigment epithelium protection in vitro. J. Cataract Refract. Surg. 2004, 30, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Ham, W.T.; Mueller, H.A. The Photopathology and Nature of the Blue Light and Near-UV Retinal Lesions Produced by Lasers and Other Optical Sources. In Laser Applications in Medicine and Biology; Springer: Boston, MA, USA, 1989; pp. 191–246. [Google Scholar]
- Ayaki, M.; Yoshimura, M.; Nakano, M.; Maruyama, Y.; Negishi, K.; Tsubota, K.; Kitazawa, M.; Hattori, A. Protective effect of blue-light shield eyewear for adults against light pollution from self-luminous devices used at night. Chronobiol. Int. 2016, 33, 134–139. [Google Scholar] [CrossRef]
- Ide, T.; Toda, I.; Miki, E.; Tsubota, K. Effect of Blue Light–Reducing Eye Glasses on Critical Flicker Frequency. Asia-Pacific J. Ophthalmol. 2015, 4, 80–85. [Google Scholar] [CrossRef]
- McCarty, C.A.; Taylor, H.R. A Review of the Epidemiologic Evidence Linking Ultraviolet Radiation and Cataracts. In Progress in Lens and Cataract Research; KARGER: Basel, Switzerland, 2002; Volume 35, pp. 21–31. [Google Scholar]
- Leung, T.W.; Li, R.; Kee, C.-S. Blue-Light Filtering Spectacle Lenses: Optical and Clinical Performances. PLoS ONE 2017, 12, e0169114. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.E.; Procyk, C.A.; Howarth, M.; Walmsley, L.; Brown, T.M. Visual input to the mouse lateral posterior and posterior thalamic nuclei: Photoreceptive origins and retinotopic order. J. Physiol. Neurosci. C 2016, 594, 1911–1929. [Google Scholar] [CrossRef] [Green Version]
- Kiser, A.K.; Deschler, E.K.; Dagnelie, G. Visual function and performance with blue-light blocking filters in age-related macular degeneration. Clin. Exp. Ophthalmol. 2008, 36, 514–520. [Google Scholar] [CrossRef]
- Alzahrani, H.S.; Khuu, S.K.; Roy, M. Modelling the effect of commercially available blue-blocking lenses on visual and non-visual functions. Clin. Exp. Optom. 2019, 1–8. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, Q.; Lin, H.; Wu, J.; Wu, Z.; Qu, S.; Bi, Y. The protective effects of blue light-blocking films with different shielding rates: A rat model study. Transl. Vis. Sci. Technol. 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narimatsu, T.; Negishi, K.; Miyake, S.; Hirasawa, M.; Osada, H.; Kurihara, T.; Tsubota, K.; Ozawa, Y. Blue light-induced inflammatory marker expression in the retinal pigment epithelium-choroid of mice and the protective effect of a yellow intraocular lens material invivo. Exp. Eye Res. 2015, 132, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- R Core Development Team R: A Language and Environment for Statistical Computing, 3.2.1. 2015. Available online: http//www.r-project.org (accessed on 14 August 2021).
- Zhao, Z.-C.; Zhou, Y.; Tan, G.; Li, J. Research progress about the effect and prevention of blue light on eyes. Int. J. Ophthalmol. 2018, 11, 1999–2003. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Álvarez, C.; Osborne, N.N. Blue light exacerbates and red light counteracts negative insults to retinal ganglion cells in situ and R28 cells in vitro. Neurochem. Int. 2019, 125, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Ortín-Martínez, A.; Valiente-Soriano, F.J.; García-Ayuso, D.; Alarcón-Martínez, L.; Jiménez-López, M.; Bernal-Garro, J.M.; Nieto-López, L.; Nadal-Nicolás, F.M.; Villegas-Péez, M.P.; Wheeler, L.A.; et al. A novel in vivo model of focal light emitting diode-induced cone-photoreceptor phototoxicity: Neuroprotection afforded by brimonidine, BDNF, PEDF or bFGF. PLoS One 2014, 9, e113798. [Google Scholar] [CrossRef] [PubMed]
- Shahinfar, S.; Edward, D.P.; Tso, M.O.M. A pathologic study of photoreceptor cell death in retinal photic injury. Curr. Eye Res. 1991, 10, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Gorman, A.; Zhou, X.; Sandra, C.; Chen, E. Involvement of caspase-3 in photoreceptor cell apoptosis induced by in vivo blue light exposure. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3349–3354. [Google Scholar]
- Garcia-Ayuso, D.; Di Pierdomenico, J.; Agudo-Barriuso, M.; Vidal-Sanz, M.; Villegas-Pérez, M. Retinal remodeling following photoreceptor degeneration causes retinal ganglion cell death. Neural Regen. Res. 2018, 13, 1885–1886. [Google Scholar] [CrossRef]
- Niwano, Y.; Iwasawa, A.; Tsubota, K.; Ayaki, M.; Negishi, K. Protective effects of blue light-blocking shades on phototoxicity in human ocular surface cells. BMJ Open Ophthalmol. 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Tanito, M.; Kaidzu, S.; Anderson, R.E. Protective effects of soft acrylic yellow filter against blue light-induced retinal damage in rats. Exp. Eye Res. 2006, 83, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Vasalauskaite, A.; Morgan, J.E.; Sengpiel, F. Plasticity in Adult Mouse Visual Cortex Following Optic Nerve Injury. Cereb. Cortex 2019, 29, 1767–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, H.E.; Stoffler, D.J. Retinal ganglion cell degeneration following loss of postsynaptic target neurons in the dorsal lateral geniculate nucleus of the adult cat. Exp. Neurol. 1992, 116, 163–171. [Google Scholar] [CrossRef]
- Pearson, H.E.; Thompson, T.P. Atrophy and degeneration of ganglion cells in central retina following loss of postsynaptic target neurons in the dorsal lateral geniculate nucleus of the adult cat. Exp. Neurol. 1993, 119, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, Y. Glutamate release and neuronal damage in ischemia. Life Sci. 2001, 69, 369–381. [Google Scholar] [CrossRef]
- Witte, O.W.; Bidmon, H.J.; Schiene, K.; Redecker, C.; Hagemann, G. Functional Differentiation of Multiple Perilesional Zones after Focal Cerebral Ischemia. J. Cereb. Blood Flow Metab. 2000, 8, 1149–1165. [Google Scholar] [CrossRef] [Green Version]
- Crawford, M.L.J.; Harwerth, R.S.; Smith, E.L.; Mills, S.; Ewing, B. Experimental glaucoma in primates: Changes in cytochrome oxidase blobs in V1 cortex. Investig. Ophthalmol. Vis. Sci. 2001, 42, 358–364. [Google Scholar]
- Davis, B.M.; Crawley, L.; Pahlitzsch, M.; Javaid, F.; Cordeiro, M.F. Glaucoma: The retina and beyond. Acta Neuropathol. 2016, 132, 807–826. [Google Scholar] [CrossRef] [Green Version]
- Harwerth, R.S.; Quigley, H.A. Visual Field Defects and Retinal Ganglion Cell Losses in Human Glaucoma Patients. Arch Ophthalmol. 2006, 6, 853–859. [Google Scholar] [CrossRef] [Green Version]
- Rifai, K.; Hornauer, M.; Buechinger, R.; Schoen, R.; Barraza-Bernal, M.; Habtegiorgis, S.; Glasenapp, C.; Wahl, S.; Mappes, T. Efficiency of ocular UV protection by clear lenses. Biomed. Opt. Express 2018, 9, 1948–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The arrive guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]

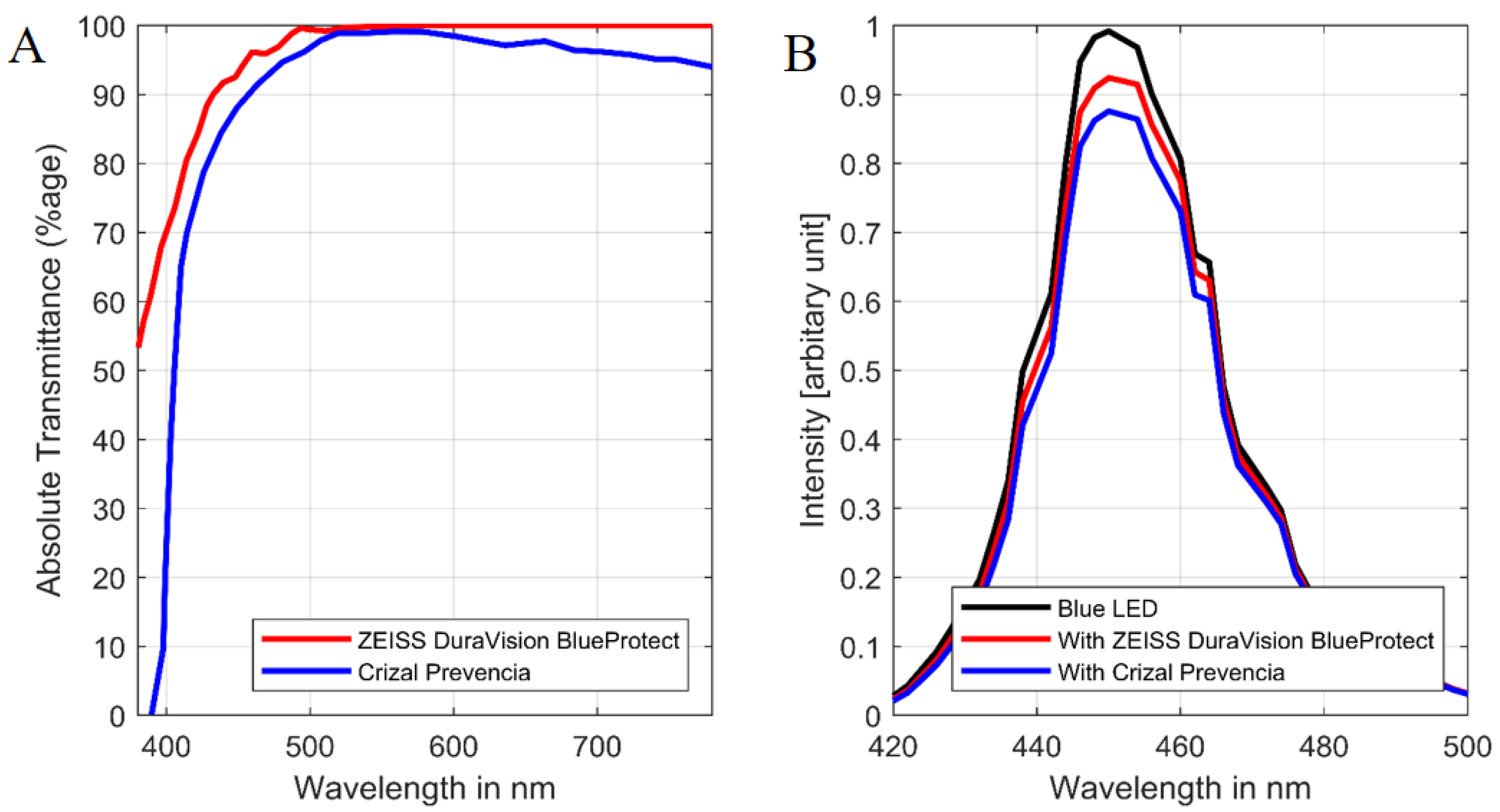

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer | Damage | NC n = 6 (12 Sections) | LE n = 6 (12 Sections) | CP n = 6 (12 Sections) | DB n = 6 (12 Sections) |
|---|---|---|---|---|---|
| Ganglion cell layer | Atrophy | 0 | 8 (66.0%) | 3 (25.0%) | 2 (16.0%) |
| Vacuolation | 0 | 1 (8.33%) | 4 (33.0%) | 4 (33%) | |
| Pyknosis | 0 | 1 (8.33%) | 1 (8.33%) | 0 | |
| Focal enlargement | 0 | 6 (50.0%) | 0 | 0 | |
| Inner plexiform layer | Atrophy | 0 | 2 (16.0%) | 0 | 2 (16%) |
| Inner nuclear nayer | Atrophy | 0 | 2 (16.0%) | 0 | 2 (16%) |
| Outer nuclear nayer | Atrophy | 0 | 0 | 2 (16.0%) | 0 |
| Decreased thickness | 0 | 3 (25.0%) | 0 | 0 | |
| Inner segment–outer Segment | Decreased thickness | 0 | 8 (66.0%) | 7 (58.0%) | 6 (50.0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Theruveethi, N.; Bui, B.V.; Joshi, M.B.; Valiathan, M.; Ganeshrao, S.B.; Gopalakrishnan, S.; Kabekkodu, S.P.; Bhat, S.S.; Surendran, S. Blue Light-Induced Retinal Neuronal Injury and Amelioration by Commercially Available Blue Light-Blocking Lenses. Life 2022, 12, 243. https://doi.org/10.3390/life12020243
Theruveethi N, Bui BV, Joshi MB, Valiathan M, Ganeshrao SB, Gopalakrishnan S, Kabekkodu SP, Bhat SS, Surendran S. Blue Light-Induced Retinal Neuronal Injury and Amelioration by Commercially Available Blue Light-Blocking Lenses. Life. 2022; 12(2):243. https://doi.org/10.3390/life12020243
Chicago/Turabian StyleTheruveethi, Nagarajan, Bang Viet Bui, Manjunath B. Joshi, Manna Valiathan, Shonraj Ballae Ganeshrao, Sivakumar Gopalakrishnan, Shama Prasada Kabekkodu, Shailaja S. Bhat, and Sudarshan Surendran. 2022. "Blue Light-Induced Retinal Neuronal Injury and Amelioration by Commercially Available Blue Light-Blocking Lenses" Life 12, no. 2: 243. https://doi.org/10.3390/life12020243
APA StyleTheruveethi, N., Bui, B. V., Joshi, M. B., Valiathan, M., Ganeshrao, S. B., Gopalakrishnan, S., Kabekkodu, S. P., Bhat, S. S., & Surendran, S. (2022). Blue Light-Induced Retinal Neuronal Injury and Amelioration by Commercially Available Blue Light-Blocking Lenses. Life, 12(2), 243. https://doi.org/10.3390/life12020243