
Plant-Derived Recombinant Vaccines against Zoonotic Viruses

 ,
,  ,
,
Abstract
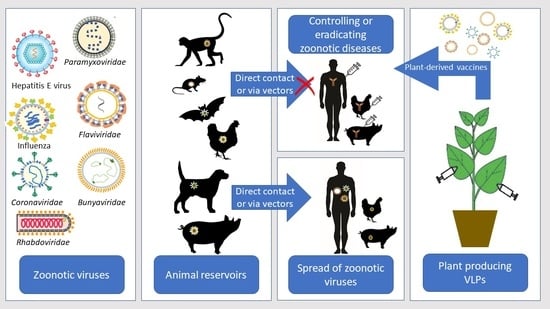
1. Introduction
2. Plants as an Expression System for Vaccine Production
3. Virus-like Particles (VLPs) for Vaccine Development
4. Influenza Viruses
5. Emerging Coronaviruses
6. Filoviruses
6.1. Ebola Virus (EBOV)
6.2. Marburg Virus (MV)
7. Bunyavirales
7.1. Crimean–Congo Hemorrhagic Fever Virus (CCHFV)
7.2. Rift Valley Fever Virus (RVFV)
7.3. Hantaviruses
8. Togaviruses
Chikungunya Virus (CHIKV)
9. Flaviviruses
9.1. West Nile Virus (WNV)
9.2. Dengue Virus (DENV)
9.3. Zika Virus (ZIKV)
9.4. Yellow Fever Virus (YFV)
9.5. Japanese Encephalitis Virus (JEV)
9.6. Tick-Borne Encephalitis Virus (TBEV)
10. Human Immunodeficiency Viruses (HIV)
11. Rabdoviruses
Rabies Virus (RABV)
12. Hepatitis E Virus
13. Newcastle Disease Virus (NDV)
14. Henipaviruses
15. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reed, K.D. Viral Zoonoses. Ref. Module Biomed. Sci. 2018. [Google Scholar] [CrossRef]
- Martins, S.B.; Häsler, B.; Rushton, J. Economic Aspects of Zoonoses: Impact of Zoonoses on the Food Industry. In Zoonoses—Infections Affecting Humans and Animals: Focus on Public Health Aspects; Sing, A., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 1107–1126. ISBN 978-94-017-9457-2. [Google Scholar]
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; Ievy, S.; Hossain, M.J.; El Zowalaty, M.E.; Rahman, A.T.; Ashour, H.M. Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.L.; Daszak, P. Host and Viral Traits Predict Zoonotic Spillover from Mammals. Nature 2017, 546, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D.; Watson, B.; Togami, E.; Daszak, P.; Mazet, J.A.; Chrisman, C.J.; Rubin, E.M.; Wolfe, N.; Morel, C.M.; Gao, G.F.; et al. Building a Global Atlas of Zoonotic Viruses. Bull. World Health Organ. 2018, 96, 292–294. [Google Scholar] [CrossRef]
- Belay, E.D.; Kile, J.C.; Hall, A.J.; Barton-Behravesh, C.; Parsons, M.B.; Salyer, S.J.; Walke, H. Zoonotic Disease Programs for Enhancing Global Health Security. Emerg. Infect. Dis. J. 2017, 23, S65. [Google Scholar] [CrossRef] [PubMed]
- Rantsios, A.T. Zoonoses. Encycl. Food Health 2016, 645–653. [Google Scholar] [CrossRef]
- Institute of Medicine (US) Forum on Microbial Threats. Vector-Borne Diseases: Understanding the Environmental, Human Health, and Ecological Connections, Workshop Summary; National Academies Press: Washington, DC, USA, 2008. [Google Scholar]
- Gebreyes, W.A.; Dupouy-Camet, J.; Newport, M.J.; Oliveira, C.J.B.; Schlesinger, L.S.; Saif, Y.M.; Kariuki, S.; Saif, L.J.; Saville, W.; Wittum, T.; et al. The Global One Health Paradigm: Challenges and Opportunities for Tackling Infectious Diseases at the Human, Animal, and Environment Interface in Low-Resource Settings. PLoS Negl. Trop. Dis. 2014, 8, e3257. [Google Scholar] [CrossRef] [PubMed]
- Schillberg, S.; Finnern, R. Plant Molecular Farming for the Production of Valuable Proteins—Critical Evaluation of Achievements and Future Challenges. J. Plant Physiol. 2021, 258–259, 153359. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant Molecular Farming of Virus-like Nanoparticles as Vaccines and Reagents. WIREs Nanomed. Nanobiotechnol. 2020, 12, e1587. [Google Scholar] [CrossRef] [PubMed]
- Shanmugaraj, B.; Bulaon, C.J.I.; Phoolcharoen, W. Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production. Plants 2020, 9, 842. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant-Based Vaccines against Viruses. Virol. J. 2014, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Buyel, J.F. Molecular Farming—The Slope of Enlightenment. Biotechnol. Adv. 2020, 40, 107519. [Google Scholar] [CrossRef]
- Gómez, M.L.; Huang, X.; Alvarez, D.; He, W.; Baysal, C.; Zhu, C.; Armario-Najera, V.; Perera, A.B.; Bennasser, P.C.; Saba-Mayoral, A.; et al. Contributions of the International Plant Science Community to the Fight against Human Infectious Diseases–Part 1: Epidemic and Pandemic Diseases. Plant Biotechnol. J. 2021, 19, 1901–1920. [Google Scholar] [CrossRef]
- Ma, J.K.-C.; Drake, P.M.W.; Christou, P. The Production of Recombinant Pharmaceutical Proteins in Plants. Nat. Rev. Genet. 2003, 4, 794–805. [Google Scholar] [CrossRef]
- Ward, B.J.; Makarkov, A.; Séguin, A.; Pillet, S.; Trépanier, S.; Dhaliwall, J.; Libman, M.D.; Vesikari, T.; Landry, N. Efficacy, Immunogenicity, and Safety of a Plant-Derived, Quadrivalent, Virus-like Particle Influenza Vaccine in Adults (18–64 Years) and Older Adults (≥65 Years): Two Multicentre, Randomised Phase 3 Trials. Lancet 2020, 396, 1491–1503. [Google Scholar] [CrossRef]
- Vermij, P.; Waltz, E. USDA Approves the First Plant-Based Vaccine. Nat. Biotechnol. 2006, 24, 234. [Google Scholar]
- Lai, K.S.; Yusoff, K.; Mahmood, M. Functional Ectodomain of the Hemagglutinin-Neuraminidase Protein Is Expressed in Transgenic Tobacco Cells as a Candidate Vaccine against Newcastle Disease Virus. Plant Cell Tissue Organ Cult. 2013, 1, 117–121. [Google Scholar] [CrossRef]
- Elelyso® for Gaucher Disease. Available online: https://protalix.com/about/elelyso/ (accessed on 19 November 2021).
- Peyret, H.; Brown, J.K.M.; Lomonossoff, G.P. Improving Plant Transient Expression through the Rational Design of Synthetic 5′ and 3′ Untranslated Regions. Plant Methods 2019, 15, 108. [Google Scholar] [CrossRef] [PubMed]
- Montero-Morales, L.; Steinkellner, H. Advanced Plant-Based Glycan Engineering. Front. Bioeng. Biotechnol. 2018, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Drossard, J. Downstream Processing of Plant-Derived Recombinant Therapeutic Proteins. In Molecular Farming; John Wiley & Sons: Hoboken, NJ, USA, 2004; pp. 217–231. ISBN 978-3-527-60363-3. [Google Scholar]
- Streatfield, S.J. Approaches to Achieve High-Level Heterologous Protein Production in Plants. Plant Biotechnol. J. 2007, 5, 2–15. [Google Scholar] [CrossRef]
- Jutras, P.V.; D’Aoust, M.-A.; Couture, M.M.-J.; Vézina, L.-P.; Goulet, M.-C.; Michaud, D.; Sainsbury, F. Modulating Secretory Pathway PH by Proton Channel Co-Expression Can Increase Recombinant Protein Stability in Plants. Biotechnol. J. 2015, 10, 1478–1486. [Google Scholar] [CrossRef]
- Jutras, P.V.; Dodds, I.; van der Hoorn, R.A. Proteases of Nicotiana Benthamiana: An Emerging Battle for Molecular Farming. Curr. Opin. Biotechnol. 2020, 61, 60–65. [Google Scholar] [CrossRef]
- Chung, Y.H.; Church, D.; Koellhoffer, E.C.; Osota, E.; Shukla, S.; Rybicki, E.P.; Pokorski, J.K.; Steinmetz, N.F. Integrating Plant Molecular Farming and Materials Research for Next-Generation Vaccines. Nat. Rev. Mater. 2021, 1–17. [Google Scholar] [CrossRef]
- LeBlanc, Z.; Waterhouse, P.; Bally, J. Plant-Based Vaccines: The Way Ahead? Viruses 2021, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Schillberg, S. Molecular Farming: Plant-Made Pharmaceuticals and Technical Proteins; Wiley-VCH: Weinheim, Germany, 2004; ISBN 978-3-527-60363-3. [Google Scholar]
- Arntzen, C. Plant-Made Pharmaceuticals: From “Edible Vaccines” to Ebola Therapeutics. Plant Biotechnol. J. 2015, 13, 1013–1016. [Google Scholar] [CrossRef]
- Tacket, C.O.; Mason, H.S.; Losonsky, G.; Estes, M.K.; Levine, M.M.; Arntzen, C.J. Human Immune Responses to a Novel Norwalk Virus Vaccine Delivered in Transgenic Potatoes. J. Infect. Dis. 2000, 182, 302–305. [Google Scholar] [CrossRef]
- Tacket, C.O.; Mason, H.S.; Losonsky, G.; Clements, J.D.; Levine, M.M.; Arntzen, C.J. Immunogenicity in Humans of a Recombinant Bacterial Antigen Delivered in a Transgenic Potato. Nat. Med. 1998, 4, 607–609. [Google Scholar] [CrossRef]
- Thanavala, Y.; Mahoney, M.; Pal, S.; Scott, A.; Richter, L.; Natarajan, N.; Goodwin, P.; Arntzen, C.J.; Mason, H.S. Immunogenicity in Humans of an Edible Vaccine for Hepatitis B. Proc. Natl. Acad. Sci. USA 2005, 102, 3378–3382. [Google Scholar] [CrossRef] [PubMed]
- Kolotilin, I.; Topp, E.; Cox, E.; Devriendt, B.; Conrad, U.; Joensuu, J.; Stöger, E.; Warzecha, H.; McAllister, T.; Potter, A.; et al. Plant-Based Solutions for Veterinary Immunotherapeutics and Prophylactics. Vet. Res. 2014, 45, 117. [Google Scholar] [CrossRef]
- Liew, P.S.; Hair-Bejo, M. Farming of Plant-Based Veterinary Vaccines and Their Applications for Disease Prevention in Animals. Adv. Virol. 2015, 2015, 936940. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E. History and Promise of Plant-Made Vaccines for Animals. In Prospects of Plant-Based Vaccines in Veterinary Medicine; MacDonald, J., Ed.; Springer: Cham, Switzerland, 2018; pp. 1–22. ISBN 978-3-319-90137-4. [Google Scholar]
- Keshavareddy, G.; Kumar, A.R.V.; Ramu, V.S. Methods of Plant Transformation—A Review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2656–2668. [Google Scholar] [CrossRef]
- Barampuram, S.; Zhang, Z.J. Recent Advances in Plant Transformation. Methods Mol. Biol. Clifton 2011, 701, 1–35. [Google Scholar] [CrossRef]
- Krenek, P.; Samajova, O.; Luptovciak, I.; Doskocilova, A.; Komis, G.; Samaj, J. Transient Plant Transformation Mediated by Agrobacterium Tumefaciens: Principles, Methods and Applications. Biotechnol. Adv. 2015, 33, 1024–1042. [Google Scholar] [CrossRef]
- Mardanova, E.S.; Blokhina, E.A.; Tsybalova, L.M.; Peyret, H.; Lomonossoff, G.P.; Ravin, N.V. Efficient Transient Expression of Recombinant Proteins in Plants by the Novel PEff Vector Based on the Genome of Potato Virus X. Front. Plant Sci. 2017, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Naseri, Z.; Khezri, G.; Davarpanah, S.J.; Ofoghi, H. Virus-Based Vectors: A New Approach for Production of Recombinant Proteins. J. Appl. Biotechnol. Rep. 2019, 6, 6–14. [Google Scholar] [CrossRef]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Viral Vectors for the Expression of Proteins in Plants. Curr. Opin. Biotechnol. 2007, 18, 134–141. [Google Scholar] [CrossRef]
- Sainsbury, F.; Thuenemann, E.C.; Lomonossoff, G.P. PEAQ: Versatile Expression Vectors for Easy and Quick Transient Expression of Heterologous Proteins in Plants. Plant Biotechnol. J. 2009, 7, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Hefferon, K. Application of Plant Viruses in Biotechnology, Medicine, and Human Health. Viruses 2021, 13, 1697. [Google Scholar] [CrossRef]
- Peyret, H.; Lomonossoff, G.P. When Plant Virology Met Agrobacterium: The Rise of the Deconstructed Clones. Plant Biotechnol. J. 2015, 13, 1121–1135. [Google Scholar] [CrossRef]
- Sainsbury, F. Innovation in Plant-Based Transient Protein Expression for Infectious Disease Prevention and Preparedness. Curr. Opin. Biotechnol. 2020, 61, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Bally, J.; Jung, H.; Mortimer, C.; Naim, F.; Philips, J.G.; Hellens, R.; Bombarely, A.; Goodin, M.M.; Waterhouse, P.M. The Rise and Rise of Nicotiana Benthamiana: A Plant for All Reasons. Annu. Rev. Phytopathol. 2018, 56, 405–426. [Google Scholar] [CrossRef] [PubMed]
- Shahid, N.; Daniell, H. Plant-Based Oral Vaccines against Zoonotic and Non-Zoonotic Diseases. Plant Biotechnol. J. 2016, 14, 2079–2099. [Google Scholar] [CrossRef] [PubMed]
- Twyman, R.M.; Christou, P. Plant Transformation Technology: Particle Bombardment. In Handbook of Plant Biotechnology; John Wiley & Sons: Hoboken, NJ, USA, 2004; ISBN 978-0-470-86914-7. [Google Scholar]
- Xu, J.; Zhang, N. On the Way to Commercializing Plant Cell Culture Platform for Biopharmaceuticals: Present Status and Prospect. Pharm. Bioprocess. 2014, 2, 499–518. [Google Scholar] [CrossRef]
- Marsian, J.; Lomonossoff, G.P. Molecular Pharming—VLPs Made in Plants. Curr. Opin. Biotechnol. 2016, 37, 201–206. [Google Scholar] [CrossRef]
- Zeltins, A. Protein Complexes and Virus-Like Particle Technology. Subcell. Biochem. 2018, 88, 379–405. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, X.; Gai, W.; Wong, G.; Wang, H.; Jin, H.; Feng, N.; Zhao, Y.; Zhang, W.; Li, N.; et al. Novel Chimeric Virus-like Particles Vaccine Displaying MERS-CoV Receptor-Binding Domain Induce Specific Humoral and Cellular Immune Response in Mice. Antivir. Res. 2017, 140, 55–61. [Google Scholar] [CrossRef]
- Chen, Q.; Lai, H. Plant-Derived Virus-like Particles as Vaccines. Hum. Vaccines Immunother. 2013, 9, 26–49. [Google Scholar] [CrossRef]
- Rutkowska, D.A.; Mokoena, N.B.; Tsekoa, T.L.; Dibakwane, V.S.; O’Kennedy, M.M. Plant-Produced Chimeric Virus-like Particles—A New Generation Vaccine against African Horse Sickness. BMC Vet. Res. 2019, 15, 432. [Google Scholar] [CrossRef]
- Zahmanova, G.; Mazalovska, M.; Takova, K.; Toneva, V.; Minkov, I.; Peyret, H.; Lomonossoff, G. Efficient Production of Chimeric Hepatitis B Virus-Like Particles Bearing an Epitope of Hepatitis E Virus Capsid by Transient Expression in Nicotiana Benthamiana. Life 2021, 11, 64. [Google Scholar] [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like Particles: Preparation, Immunogenicity and Their Roles as Nanovaccines and Drug Nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Lomonossoff, G.P. Virus Particles and the Uses of Such Particles in Bio- and Nanotechnology. In Recent Advances in Plant Virology; Caister Academic Press: Poole, UK, 2011. [Google Scholar]
- Mardanova, E.S.; Kotlyarov, R.Y.; Kuprianov, V.V.; Stepanova, L.A.; Tsybalova, L.M.; Lomonosoff, G.P.; Ravin, N.V. Rapid High-Yield Expression of a Candidate Influenza Vaccine Based on the Ectodomain of M2 Protein Linked to Flagellin in Plants Using Viral Vectors. BMC Biotechnol. 2015, 15, 42. [Google Scholar] [CrossRef]
- Matić, S.; Rinaldi, R.; Masenga, V.; Noris, E. Efficient Production of Chimeric Human Papillomavirus 16 L1 Protein Bearing the M2e Influenza Epitope in Nicotiana Benthamiana Plants. BMC Biotechnol. 2011, 11, 106. [Google Scholar] [CrossRef]
- Medicago Inc. Plant-Derived Vaccines. Biopharma Dealmakers (Biopharm Deal). 2018. Available online: https://www.nature.com/articles/d43747-020-00537-y (accessed on 19 November 2021).
- Pillet, S.; Aubin, É.; Trépanier, S.; Bussière, D.; Dargis, M.; Poulin, J.-F.; Yassine-Diab, B.; Ward, B.J.; Landry, N. A Plant-Derived Quadrivalent Virus like Particle Influenza Vaccine Induces Cross-Reactive Antibody and T Cell Response in Healthy Adults. Clin. Immunol. 2016, 168, 72–87. [Google Scholar] [CrossRef]
- Balke, I.; Zeltins, A. Recent Advances in the Use of Plant Virus-Like Particles as Vaccines. Viruses 2020, 12, 270. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus Taxonomy: The Database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef]
- Hay, A.J.; Gregory, V.; Douglas, A.R.; Lin, Y.P. The Evolution of Human Influenza Viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1861–1870. [Google Scholar] [CrossRef]
- Cox, N.J.; Subbarao, K. Global Epidemiology of Influenza: Past and Present. Annu. Rev. Med. 2000, 51, 407–421. [Google Scholar] [CrossRef]
- Krammer, F. The Human Antibody Response to Influenza A Virus Infection and Vaccination. Nat. Rev. Immunol. 2019, 19, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Li, X.; Goraya, M.U.; Wang, S.; Chen, J.-L. Evolution of Influenza A Virus by Mutation and Re-Assortment. Int. J. Mol. Sci. 2017, 18, 1650. [Google Scholar] [CrossRef] [PubMed]
- Proença-Módena, J.L.; Macedo, I.S.; Arruda, E. H5N1 Avian Influenza Virus: An Overview. Braz. J. Infect. Dis. 2007, 11, 125–133. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Paget, J.; Spreeuwenberg, P.; Charu, V.; Taylor, R.J.; Iuliano, A.D.; Bresee, J.; Simonsen, L.; Viboud, C. Global Seasonal Influenza-associated Mortality Collaborator Network and GLaMOR Collaborating Teams* Global Mortality Associated with Seasonal Influenza Epidemics: New Burden Estimates and Predictors from the GLaMOR Project. J. Glob. Health 2019, 9, 020421. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, C. DARPA Effort Speeds Bio-Threat Response. Available online: https://www.army.mil/article/47617/darpa_effort_speeds_bio_threat_response (accessed on 19 November 2021).
- Medicago Inc. Final Short Form Prospectus. Available online: https://sec.report/otc/financial-report/93315 (accessed on 19 November 2021).
- D’Aoust, M.-A.; Lavoie, P.-O.; Couture, M.M.-J.; Trépanier, S.; Guay, J.-M.; Dargis, M.; Mongrand, S.; Landry, N.; Ward, B.J.; Vézina, L.-P. Influenza Virus-like Particles Produced by Transient Expression in Nicotiana Benthamiana Induce a Protective Immune Response against a Lethal Viral Challenge in Mice. Plant Biotechnol. J. 2008, 6, 930–940. [Google Scholar] [CrossRef]
- Landry, N.; Ward, B.J.; Trépanier, S.; Montomoli, E.; Dargis, M.; Lapini, G.; Vézina, L.-P. Preclinical and Clinical Development of Plant-Made Virus-Like Particle Vaccine against Avian H5N1 Influenza. PLoS ONE 2010, 5, e15559. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Aubin, É.; Trépanier, S.; Poulin, J.-F.; Yassine-Diab, B.; ter Meulen, J.; Ward, B.J.; Landry, N. Humoral and Cell-Mediated Immune Responses to H5N1 Plant-Made Virus-like Particle Vaccine Are Differentially Impacted by Alum and GLA-SE Adjuvants in a Phase 2 Clinical Trial. npj Vaccines 2018, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Couillard, J.; Trépanier, S.; Poulin, J.-F.; Yassine-Diab, B.; Guy, B.; Ward, B.J.; Landry, N. Immunogenicity and Safety of a Quadrivalent Plant-Derived Virus like Particle Influenza Vaccine Candidate—Two Randomized Phase II Clinical Trials in 18 to 49 and ≥50 Years Old Adults. PLoS ONE 2019, 14, e0216533. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, S.; Dassarma, B.; Bhattacharya, M.; Matsabisa, M.G. Plant-Based Vaccine Research Development against Viral Diseases with Emphasis on Ebola Virus Disease: A Review Study. Curr. Opin. Pharmacol. 2021, 60, 261–267. [Google Scholar] [CrossRef]
- Home—ClinicalTrials.Gov. Available online: https://www.clinicaltrials.gov/ (accessed on 19 November 2021).
- Smith, T.; O’Kennedy, M.M.; Wandrag, D.B.R.; Adeyemi, M.; Abolnik, C. Efficacy of a Plant-Produced Virus-like Particle Vaccine in Chickens Challenged with Influenza A H6N2 Virus. Plant Biotechnol. J. 2020, 18, 502–512. [Google Scholar] [CrossRef]
- Shoji, Y.; Bi, H.; Musiychuk, K.; Rhee, A.; Horsey, A.; Roy, G.; Green, B.; Shamloul, M.; Farrance, C.E.; Taggart, B.; et al. Plant-Derived Hemagglutinin Protects Ferrets against Challenge Infection with the A/Indonesia/05/05 Strain of Avian Influenza. Vaccine 2009, 27, 1087–1092. [Google Scholar] [CrossRef]
- Shoji, Y.; Chichester, J.A.; Bi, H.; Musiychuk, K.; de la Rosa, P.; Goldschmidt, L.; Horsey, A.; Ugulava, N.; Palmer, G.A.; Mett, V.; et al. Plant-Expressed HA as a Seasonal Influenza Vaccine Candidate. Vaccine 2008, 26, 2930–2934. [Google Scholar] [CrossRef]
- Shoji, Y.; Farrance, C.E.; Bautista, J.; Bi, H.; Musiychuk, K.; Horsey, A.; Park, H.; Jaje, J.; Green, B.J.; Shamloul, M.; et al. A Plant-Based System for Rapid Production of Influenza Vaccine Antigens. Influenza Other Respir. Viruses 2012, 6, 204–210. [Google Scholar] [CrossRef]
- Shoji, Y.; Jones, R.M.; Mett, V.; Chichester, J.A.; Musiychuk, K.; Sun, X.; Tumpey, T.M.; Green, B.J.; Shamloul, M.; Norikane, J.; et al. A Plant-Produced H1N1 Trimeric Hemagglutinin Protects Mice from a Lethal Influenza Virus Challenge. Hum. Vaccines Immunother. 2013, 9, 553–560. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Phan, H.T.; Pham, V.T.; Ho, T.T.; Pham, N.B.; Chu, H.H.; Vu, T.H.; Abdelwhab, E.M.; Scheibner, D.; Mettenleiter, T.C.; Hanh, T.X.; et al. Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1. Vaccines 2020, 8, 593. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.; Ho, T.T.; Chu, H.H.; Vu, T.H.; Gresch, U.; Conrad, U. Neutralizing Immune Responses Induced by Oligomeric H5N1-Hemagglutinins from Plants. Vet. Res. 2017, 48, 53. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Phan, H.T.; Hause, B.; Hause, G.; Arcalis, E.; Stoger, E.; Maresch, D.; Altmann, F.; Joensuu, J.; Conrad, U. Influence of Elastin-Like Polypeptide and Hydrophobin on Recombinant Hemagglutinin Accumulations in Transgenic Tobacco Plants. PLoS ONE 2014, 9, e99347. [Google Scholar] [CrossRef]
- Phan, H.T.; Gresch, U.; Conrad, U. In Vitro-Formulated Oligomers of Strep-Tagged Avian Influenza Haemagglutinin Produced in Plants Cause Neutralizing Immune Responses. Front. Bioeng. Biotechnol. 2018, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.; Pohl, J.; Floss, D.M.; Rabenstein, F.; Veits, J.; Le, B.T.; Chu, H.H.; Hause, G.; Mettenleiter, T.; Conrad, U. ELPylated Haemagglutinins Produced in Tobacco Plants Induce Potentially Neutralizing Antibodies against H5N1 Viruses in Mice. Plant Biotechnol. J. 2013, 11, 582–593. [Google Scholar] [CrossRef]
- Mett, V.; Musiychuk, K.; Bi, H.; Farrance, C.E.; Horsey, A.; Ugulava, N.; Shoji, Y.; de la Rosa, P.; Palmer, G.A.; Rabindran, S.; et al. A Plant-Produced Influenza Subunit Vaccine Protects Ferrets against Virus Challenge. Influenza Other Respir. Viruses 2008, 2, 33–40. [Google Scholar] [CrossRef]
- Musiychuk, K.; Stephenson, N.; Bi, H.; Farrance, C.E.; Orozovic, G.; Brodelius, M.; Brodelius, P.; Horsey, A.; Ugulava, N.; Shamloul, A.-M.; et al. A Launch Vector for the Production of Vaccine Antigens in Plants. Influenza Other Respir. Viruses 2007, 1, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Redkiewicz, P.; Sirko, A.; Kamel, K.A.; Góra-Sochacka, A. Plant Expression Systems for Production of Hemagglutinin as a Vaccine against Influenza Virus. Acta Biochim. Pol. 2014, 61, 551–560. [Google Scholar] [CrossRef]
- Ceballo, Y.; Tiel, K.; López, A.; Cabrera, G.; Pérez, M.; Ramos, O.; Rosabal, Y.; Montero, C.; Menassa, R.; Depicker, A.; et al. High Accumulation in Tobacco Seeds of Hemagglutinin Antigen from Avian (H5N1) Influenza. Transgenic Res. 2017, 26, 775–789. [Google Scholar] [CrossRef]
- Nahampun, H.N.; Bosworth, B.; Cunnick, J.; Mogler, M.; Wang, K. Expression of H3N2 Nucleoprotein in Maize Seeds and Immunogenicity in Mice. Plant Cell Rep. 2015, 34, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Thuenemann, E.C.; Lenzi, P.; Love, A.J.; Taliansky, M.; Bécares, M.; Zuñiga, S.; Enjuanes, L.; Zahmanova, G.G.; Minkov, I.N.; Matić, S.; et al. The Use of Transient Expression Systems for the Rapid Production of Virus-like Particles in Plants. Curr. Pharm. Des. 2013, 19, 5564–5573. [Google Scholar] [CrossRef]
- Zahmanova, G.G.; Mazalovska, M.; Takova, K.H.; Toneva, V.T.; Minkov, I.N.; Mardanova, E.S.; Ravin, N.V.; Lomonossoff, G.P. Rapid High-Yield Transient Expression of Swine Hepatitis E ORF2 Capsid Proteins in Nicotiana Benthamiana Plants and Production of Chimeric Hepatitis E Virus-Like Particles Bearing the M2e Influenza Epitope. Plants 2019, 9, 29. [Google Scholar] [CrossRef]
- Meshcheryakova, Y.A.; Eldarov, M.A.; Migunov, A.I.; Stepanova, L.A.; Repko, I.A.; Kiselev, C.I.; Lomonossoff, G.P.; Skryabin, K.G. Cowpea Mosaic Virus Chimeric Particles Bearing the Ectodomain of Matrix Protein 2 (M2E) of the Influenza a Virus: Production and Characterization. Mol. Biol. 2009, 43, 685–694. [Google Scholar] [CrossRef]
- Petukhova, N.V.; Gasanova, T.V.; Ivanov, P.A.; Atabekov, J.G. High-Level Systemic Expression of Conserved Influenza Epitope in Plants on the Surface of Rod-Shaped Chimeric Particles. Viruses 2014, 6, 1789–1800. [Google Scholar] [CrossRef]
- Ravin, N.V.; Kotlyarov, R.Y.; Mardanova, E.S.; Kuprianov, V.V.; Migunov, A.I.; Stepanova, L.A.; Tsybalova, L.M.; Kiselev, O.I.; Skryabin, K.G. Plant-Produced Recombinant Influenza Vaccine Based on Virus-like HBc Particles Carrying an Extracellular Domain of M2 Protein. Biochem. Mosc. 2012, 77, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Petukhova, N.V.; Gasanova, T.V.; Stepanova, L.A.; Rusova, O.A.; Potapchuk, M.V.; Korotkov, A.V.; Skurat, E.V.; Tsybalova, L.M.; Kiselev, O.I.; Ivanov, P.A.; et al. Immunogenicity and Protective Efficacy of Candidate Universal Influenza A Nanovaccines Produced in Plants by Tobacco Mosaic Virus-Based Vectors. Curr. Pharm. Des. 2013, 19, 5587–5600. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, E.A.; Mardanova, E.S.; Stepanova, L.A.; Tsybalova, L.M.; Ravin, N.V. Plant-Produced Recombinant Influenza A Virus Candidate Vaccine Based on Flagellin Linked to Conservative Fragments of M2 Protein and Hemagglutintin. Plants 2020, 9, 162. [Google Scholar] [CrossRef]
- Pal, M.; Berhanu, G.; Desalegn, C.; Kandi, V. Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2): An Update. Cureus 2020, 12, e7423. [Google Scholar] [CrossRef]
- Liu, D.X.; Liang, J.Q.; Fung, T.S. Human Coronavirus-229E, -OC43, -NL63, and -HKU1 (Coronaviridae). Encycl. Virol. 2021, 428–440. [Google Scholar] [CrossRef]
- Ying, W.; Hao, Y.; Zhang, Y.; Peng, W.; Qin, E.; Cai, Y.; Wei, K.; Wang, J.; Chang, G.; Sun, W.; et al. Proteomic Analysis on Structural Proteins of Severe Acute Respiratory Syndrome Coronavirus. Proteomics 2004, 4, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, L. SARS-CoV-2 Spike Protein: A Key Target for Eliciting Persistent Neutralizing Antibodies. Signal Transduct. Target. Ther. 2021, 6, 95. [Google Scholar] [CrossRef]
- Tusé, D.; Nandi, S.; McDonald, K.A.; Buyel, J.F. The Emergency Response Capacity of Plant-Based Biopharmaceutical Manufacturing-What It Is and What It Could Be. Front. Plant Sci. 2020, 11, 1573. [Google Scholar] [CrossRef] [PubMed]
- Capell, T.; Twyman, R.M.; Armario-Najera, V.; Ma, J.K.-C.; Schillberg, S.; Christou, P. Potential Applications of Plant Biotechnology against SARS-CoV-2. Trends Plant Sci. 2020, 25, 635–643. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S. Will Plant-Made Biopharmaceuticals Play a Role in the Fight against COVID-19? Expert Opin. Biol. Ther. 2020, 20, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.J.; Gobeil, P.; Séguin, A.; Atkins, J.; Boulay, I.; Charbonneau, P.-Y.; Couture, M.; D’Aoust, M.-A.; Dhaliwall, J.; Finkle, C.; et al. Phase 1 Randomized Trial of a Plant-Derived Virus-like Particle Vaccine for COVID-19. Nat. Med. 2021, 27, 1071–1078. [Google Scholar] [CrossRef]
- Medicago Inc. Positive Results from Phase 3 Study of Medicago’s Plant-Based COVID-19 Vaccine Candidate Announced. Available online: https://www.news-medical.net/news/20211209/Positive-results-from-Phase-3-study-of-Medicagoe28099s-plant-based-COVID-19-vaccine-candidate-announced.aspx (accessed on 14 January 2022).
- Kentucky BioProcessing, Inc. A Phase I/II, First-in-Human, Observer-Blinded, Randomized, Placebo-Controlled, Parallel Group Study to Evaluate the Safety and Immunogenicity of TAP-COVID-19 SARS-CoV-2 Vaccine with CpG Adjuvant in Healthy Adults Aged 18–49 and 50–85. Clinicaltrials.gov. 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT04473690 (accessed on 14 January 2022).
- Medicago Inc. Medicago and GSK Start Phase 3 Trial of Adjuvanted COVID-19 Vaccine Candidate. Available online: https://medicago.com/en/press-release/medicago-and-gsk-start-phase-3-trial-of-adjuvanted-covid-19-vaccine-candidate/ (accessed on 12 January 2022).
- iBio Inc. IBio Reports Successful Preclinical Immunization Studies with Next-Gen Nucleocapsid COVID-19 Vaccine Candidate. Available online: https://ibioinc.com/ibio-reports-successful-preclinical-immunization-studies-with-next-gen-nucleocapsid-covid-19-vaccine-candidate/ (accessed on 20 November 2021).
- British American Tobacco. BAT Makes Progress on COVID-19 Vaccine Provides Community Support. Available online: https://www.bat.com/group/sites/UK__9D9KCY.nsf/vwPagesWebLive/DOBPMBZC# (accessed on 20 November 2021).
- Hardy, A. IBIO-201 Demonstrates Ability to Elicit Anti-SARS-CoV-2 Immune Response in Preclinical Studies. Available online: https://biotuesdays.com/2020/08/10/ibio-updates-ibio-201-covid-19-vaccine-candidate/ (accessed on 20 November 2021).
- Baiya Phytopharm Co Ltd. Baiya SARS-CoV-2 Vax 1 Vaccine–COVID-19 Vaccine Tracker. Available online: https://covid19.trackvaccines.org/vaccines/130/ (accessed on 20 November 2021).
- Peyret, H.; Steele, J.F.C.; Jung, J.-W.; Thuenemann, E.C.; Meshcheriakova, Y.; Lomonossoff, G.P. Producing Vaccines against Enveloped Viruses in Plants: Making the Impossible, Difficult. Vaccines 2021, 9, 780. [Google Scholar] [CrossRef]
- Pogrebnyak, N.; Golovkin, M.; Andrianov, V.; Spitsin, S.; Smirnov, Y.; Egolf, R.; Koprowski, H. Severe Acute Respiratory Syndrome (SARS) S Protein Production in Plants: Development of Recombinant Vaccine. Proc. Natl. Acad. Sci. USA 2005, 102, 9062–9067. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Ramalingam, S.; Chye, M.-L. Accumulation of Recombinant SARS-CoV Spike Protein in Plant Cytosol and Chloroplasts Indicate Potential for Development of Plant-Derived Oral Vaccines. Exp. Biol. Med. 2006, 231, 1346–1352. [Google Scholar] [CrossRef]
- Zheng, N.; Xia, R.; Yang, C.; Yin, B.; Li, Y.; Duan, C.; Liang, L.; Guo, H.; Xie, Q. Boosted Expression of the SARS-CoV Nucleocapsid Protein in Tobacco and Its Immunogenicity in Mice. Vaccine 2009, 27, 5001–5007. [Google Scholar] [CrossRef] [PubMed]
- Demurtas, O.C.; Massa, S.; Illiano, E.; De Martinis, D.; Chan, P.K.S.; Di Bonito, P.; Franconi, R. Antigen Production in Plant to Tackle Infectious Diseases Flare Up: The Case of SARS. Front. Plant Sci. 2016, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Mamedov, T.; Yuksel, D.; Ilgın, M.; Gürbüzaslan, I.; Gulec, B.; Mammadova, G.; Ozdarendeli, A.; Yetiskin, H.; Kaplan, B.; Islam Pavel, S.T.; et al. Production and Characterization of Nucleocapsid and RBD Cocktail Antigens of SARS-CoV-2 in Nicotiana Benthamiana Plant as a Vaccine Candidate against COVID-19. Vaccines 2021, 9, 1337. [Google Scholar] [CrossRef]
- Maharjan, P.M.; Cheon, J.; Jung, J.; Kim, H.; Lee, J.; Song, M.; Jeong, G.U.; Kwon, Y.; Shim, B.; Choe, S. Plant-Expressed Receptor Binding Domain of the SARS-CoV-2 Spike Protein Elicits Humoral Immunity in Mice. Vaccines 2021, 9, 978. [Google Scholar] [CrossRef]
- Hemmati, F.; Hemmati-Dinarvand, M.; Karimzade, M.; Rutkowska, D.; Eskandari, M.H.; Khanizadeh, S.; Afsharifar, A. Plant-Derived VLP: A Worthy Platform to Produce Vaccine against SARS-CoV-2. Biotechnol. Lett. 2021. [Google Scholar] [CrossRef]
- Maharjan, P.M.; Choe, S. Plant-Based COVID-19 Vaccines: Current Status, Design, and Development Strategies of Candidate Vaccines. Vaccines 2021, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Diego-Martin, B.; González, B.; Vazquez-Vilar, M.; Selma, S.; Mateos-Fernández, R.; Gianoglio, S.; Fernández-del-Carmen, A.; Orzáez, D. Pilot Production of SARS-CoV-2 Related Proteins in Plants: A Proof of Concept for Rapid Repurposing of Indoor Farms Into Biomanufacturing Facilities. Front. Plant Sci. 2020, 11, 2101. [Google Scholar] [CrossRef]
- Kumar, M.; Kumari, N.; Thakur, N.; Bhatia, S.K.; Saratale, G.D.; Ghodake, G.; Mistry, B.M.; Alavilli, H.; Kishor, D.S.; Du, X.; et al. A Comprehensive Overview on the Production of Vaccines in Plant-Based Expression Systems and the Scope of Plant Biotechnology to Combat against SARS-CoV-2 Virus Pandemics. Plants 2021, 10, 1213. [Google Scholar] [CrossRef]
- Feldmann, H.; Jones, S.; Klenk, H.-D.; Schnittler, H.-J. Ebola Virus: From Discovery to Vaccine. Nat. Rev. Immunol. 2003, 3, 677–685. [Google Scholar] [CrossRef]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011; ISBN 978-0-12-384685-3. [Google Scholar]
- Beer, B.; Kurth, R.; Bukreyev, A. Characteristics of Filoviridae: Marburg and Ebola Viruses. Naturwissenschaften 1999, 86, 8–17. [Google Scholar] [CrossRef]
- Feldmann, H.; Sanchez, A.; Geisbert, T. Filoviridae: Marburg and Ebola Viruses. In Fields Virology, 6th ed.; Wolters Kluwer Health Adis (ESP): London, UK, 2013; Volume 1, ISBN 978-1-4511-0563-6. [Google Scholar]
- Mehedi, M.; Falzarano, D.; Seebach, J.; Hu, X.; Carpenter, M.S.; Schnittler, H.-J.; Feldmann, H. A New Ebola Virus Nonstructural Glycoprotein Expressed through RNA Editing. J. Virol. 2011, 85, 5406–5414. [Google Scholar] [CrossRef] [PubMed]
- Baize, S. A Single Shot against Ebola and Marburg Virus. Nat. Med. 2005, 11, 720–721. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Geisbert, J.B.; Leung, A.; Daddario-DiCaprio, K.M.; Hensley, L.E.; Grolla, A.; Feldmann, H. Single-Injection Vaccine Protects Nonhuman Primates against Infection with Marburg Virus and Three Species of Ebola Virus. J. Virol. 2009, 83, 7296–7304. [Google Scholar] [CrossRef]
- Sullivan, N.J.; Sanchez, A.; Rollin, P.E.; Yang, Z.Y.; Nabel, G.J. Development of a Preventive Vaccine for Ebola Virus Infection in Primates. Nature 2000, 408, 605–609. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bausch, D.G.; Feldmann, H. Prospects for Immunisation against Marburg and Ebola Viruses. Rev. Med. Virol. 2010, 20, 344–357. [Google Scholar] [CrossRef]
- Sridhar, S. Clinical Development of Ebola Vaccines. Ther. Adv. Vaccines 2015, 3, 125–138. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Hu, Y.; Liang, Q.; Wei, M.; Zhu, F. Ebola Vaccines in Clinical Trial: The Promising Candidates. Hum. Vaccines Immunother. 2017, 13, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Gómez, R.; Angulo, C.; Monreal-Escalante, E.; Govea-Alonso, D.O.; De Groot, A.S.; Rosales-Mendoza, S. Design of a Multiepitopic Zaire Ebolavirus Protein and Its Expression in Plant Cells. J. Biotechnol. 2019, 295, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Phoolcharoen, W.; Bhoo, S.H.; Lai, H.; Ma, J.; Arntzen, C.J.; Chen, Q.; Mason, H.S. Expression of an Immunogenic Ebola Immune Complex in Nicotiana Benthamiana. Plant Biotechnol. J. 2011, 9, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Phoolcharoen, W.; Dye, J.M.; Kilbourne, J.; Piensook, K.; Pratt, W.D.; Arntzen, C.J.; Chen, Q.; Mason, H.S.; Herbst-Kralovetz, M.M. A Nonreplicating Subunit Vaccine Protects Mice against Lethal Ebola Virus Challenge. Proc. Natl. Acad. Sci. USA 2011, 108, 20695–20700. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Mirazimi, A.; Köksal, I.; Estrada-Pena, A.; Feldmann, H. Recent Advances in Research on Crimean-Congo Hemorrhagic Fever. J. Clin. Virol. 2015, 64, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ergönül, Ö. Crimean-Congo Haemorrhagic Fever. Lancet Infect. Dis. 2006, 6, 203–214. [Google Scholar] [CrossRef]
- Ruiz, S.I.; Zumbrun, E.E.; Nalca, A. Animal Models of Human Viral Diseases. Anim. Models Study Hum. Dis. 2017, 853–901. [Google Scholar] [CrossRef]
- Maes, P.; Alkhovsky, S.V.; Bào, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the Family Arenaviridae and the Order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef] [PubMed]
- Haferkamp, S.; Fernando, L.; Schwarz, T.F.; Feldmann, H.; Flick, R. Intracellular Localization of Crimean-Congo Hemorrhagic Fever (CCHF) Virus Glycoproteins. Virol. J. 2005, 2, 42. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghiasi, S.M.; Salmanian, A.H.; Chinikar, S.; Zakeri, S. Mice Orally Immunized with a Transgenic Plant Expressing the Glycoprotein of Crimean-Congo Hemorrhagic Fever Virus. Clin. Vaccine Immunol. 2011, 18, 2031–2037. [Google Scholar] [CrossRef] [PubMed]
- Dowall, S.D.; Carroll, M.W.; Hewson, R. Development of Vaccines against Crimean-Congo Haemorrhagic Fever Virus. Vaccine 2017, 35, 6015–6023. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, T.; Boulianne, C.; Vincent, M.J.; Pezzanite, L.; Al-Qahtani, M.M.; Al-Mazrou, Y.; Khan, A.S.; Rollin, P.E.; Swanepoel, R.; Ksiazek, T.G.; et al. Genetic Analysis of Viruses Associated with Emergence of Rift Valley Fever in Saudi Arabia and Yemen, 2000-01. Emerg. Infect. Dis. 2002, 8, 1415–1420. [Google Scholar] [CrossRef]
- Kwaśnik, M.; Rożek, W.; Rola, J. Rift Valley Fever—A Growing Threat to Humans and Animals. J. Vet. Res. 2021, 65, 7–14. [Google Scholar] [CrossRef]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley Fever: An Emerging Mosquito-Borne Disease. Annu. Rev. Entomol. 2016, 61, 395–415. [Google Scholar] [CrossRef]
- Balenghien, T.; Cardinale, E.; Chevalier, V.; Elissa, N.; Failloux, A.-B.; Jean Jose Nipomichene, T.N.; Nicolas, G.; Rakotoharinome, V.M.; Roger, M.; Zumbo, B. Towards a Better Understanding of Rift Valley Fever Epidemiology in the South-West of the Indian Ocean. Vet. Res. 2013, 44, 78. [Google Scholar] [CrossRef] [PubMed]
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley Fever Virus (Bunyaviridae: Phlebovirus): An Update on Pathogenesis, Molecular Epidemiology, Vectors, Diagnostics and Prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Indran, S.V.; Balaraman, V.; Wilson, W.C.; Richt, J.A. Molecular Aspects of Rift Valley Fever Virus and the Emergence of Reassortants. Virus Genes 2019, 55, 1–11. [Google Scholar] [CrossRef]
- Faburay, B.; Wilson, W.; McVey, D.S.; Drolet, B.S.; Weingartl, H.; Madden, D.; Young, A.; Ma, W.; Richt, J.A. Rift Valley Fever Virus Structural and Nonstructural Proteins: Recombinant Protein Expression and Immunoreactivity against Antisera from Sheep. Vector-Borne Zoonotic Dis. 2013, 13, 619–629. [Google Scholar] [CrossRef]
- Bouloy, M.; Weber, F. Molecular Biology of Rift Valley Fever Virus. Open Virol. J. 2010, 4. [Google Scholar] [CrossRef]
- Mbewana, S.; Meyers, A.E.; Rybicki, E.P. Chimaeric Rift Valley Fever Virus-Like Particle Vaccine Candidate Production in Nicotiana Benthamiana. Biotechnol. J. 2019, 14, e1800238. [Google Scholar] [CrossRef]
- Kalbina, I.; Lagerqvist, N.; Moiane, B.; Ahlm, C.; Andersson, S.; Strid, Å.; Falk, K.I. Arabidopsis Thaliana Plants Expressing Rift Valley Fever Virus Antigens: Mice Exhibit Systemic Immune Responses as the Result of Oral Administration of the Transgenic Plants. Protein Expr. Purif. 2016, 127, 61–67. [Google Scholar] [CrossRef]
- Chandy, S.; Mathai, D. Globally Emerging Hantaviruses: An Overview. Indian J. Med. Microbiol. 2017, 35, 165–175. [Google Scholar] [CrossRef]
- Hangaragi, P.S. Hantavirus: An Emerging Global Threat. Asian J. Oral Health Allied Sci. 2020, 10, 4. [Google Scholar] [CrossRef]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and Therapeutics against Hantaviruses. Front. Microbiol. 2020, 10, 2989. [Google Scholar] [CrossRef]
- Muyangwa, M.; Martynova, E.V.; Khaiboullina, S.F.; Morzunov, S.P.; Rizvanov, A.A. Hantaviral Proteins: Structure, Functions, and Role in Hantavirus Infection. Front. Microbiol. 2015, 6, 1326. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, K.; Arikawa, J. Antigenic Properties of N Protein of Hantavirus. Viruses 2014, 6, 3097–3109. [Google Scholar] [CrossRef] [PubMed]
- Khattak, S.; Darai, G.; Süle, S.; Rösen-Wolff, A. Characterization of Expression of Puumala Virus Nucleocapsid Protein in Transgenic Plants. Intervirology 2002, 45, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Khattak, S.; Darai, G.; Rösen-Wolff, A. Puumala Virus Nucleocapsid Protein Expressed in Transgenic Plants Is Not Immunogenic after Oral Administration. Virus Genes 2004, 29, 109–116. [Google Scholar] [CrossRef]
- Peyret, H.; Gehin, A.; Thuenemann, E.C.; Blond, D.; Turabi, A.E.; Beales, L.; Clarke, D.; Gilbert, R.J.C.; Fry, E.E.; Stuart, D.I.; et al. Tandem Fusion of Hepatitis B Core Antigen Allows Assembly of Virus-Like Particles in Bacteria and Plants with Enhanced Capacity to Accommodate Foreign Proteins. PLoS ONE 2015, 10, e0120751. [Google Scholar] [CrossRef]
- Ulrich, R.; Koletzki, D.; Lachmann, S.; Lundkvist, A.; Zankl, A.; Kazaks, A.; Kurth, A.; Gelderblom, H.R.; Borisova, G.; Meisel, H.; et al. New Chimaeric Hepatitis B Virus Core Particles Carrying Hantavirus (Serotype Puumala) Epitopes: Immunogenicity and Protection against Virus Challenge. J. Biotechnol. 1999, 73, 141–153. [Google Scholar] [CrossRef]
- Geldmacher, A.; Skrastina, D.; Petrovskis, I.; Borisova, G.; Berriman, J.A.; Roseman, A.M.; Crowther, R.A.; Fischer, J.; Musema, S.; Gelderblom, H.R.; et al. An Amino-Terminal Segment of Hantavirus Nucleocapsid Protein Presented on Hepatitis B Virus Core Particles Induces a Strong and Highly Cross-Reactive Antibody Response in Mice. Virology 2004, 323, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.; Lundkvist, A.; Meisel, H.; Koletzki, D.; Sjölander, K.B.; Gelderblom, H.R.; Borisova, G.; Schnitzler, P.; Darai, G.; Krüger, D.H. Chimaeric HBV Core Particles Carrying a Defined Segment of Puumala Hantavirus Nucleocapsid Protein Evoke Protective Immunity in an Animal Model. Vaccine 1998, 16, 272–280. [Google Scholar] [CrossRef]
- Koletzki, D.; Biel, S.S.; Meisel, H.; Nugel, E.; Gelderblom, H.R.; Krüger, D.H.; Ulrich, R. HBV Core Particles Allow the Insertion and Surface Exposure of the Entire Potentially Protective Region of Puumala Hantavirus Nucleocapsid Protein. Biol. Chem. 1999, 380, 325–333. [Google Scholar] [CrossRef]
- Koletzki, D.; Lundkvist, A.; Sjölander, K.B.; Gelderblom, H.R.; Niedrig, M.; Meisel, H.; Krüger, D.H.; Ulrich, R. Puumala (PUU) Hantavirus Strain Differences and Insertion Positions in the Hepatitis B Virus Core Antigen Influence B-Cell Immunogenicity and Protective Potential of Core-Derived Particles. Virology 2000, 276, 364–375. [Google Scholar] [CrossRef]
- Geldmacher, A.; Skrastina, D.; Borisova, G.; Petrovskis, I.; Krüger, D.H.; Pumpens, P.; Ulrich, R. A Hantavirus Nucleocapsid Protein Segment Exposed on Hepatitis B Virus Core Particles Is Highly Immunogenic in Mice When Applied without Adjuvants or in the Presence of Pre-Existing Anti-Core Antibodies. Vaccine 2005, 23, 3973–3983. [Google Scholar] [CrossRef]
- Simon, F.; Savini, H.; Parola, P. Chikungunya: A Paradigm of Emergence and Globalization of Vector-Borne Diseases. Med. Clin. N. Am. 2008, 92, 1323–1343. [Google Scholar] [CrossRef] [PubMed]
- Simon, F.; Javelle, E.; Oliver, M.; Leparc-Goffart, I.; Marimoutou, C. Chikungunya Virus Infection. Curr. Infect. Dis. Rep. 2011, 13, 218–228. [Google Scholar] [CrossRef]
- Consuegra-Rodríguez, M.P.; Hidalgo-Zambrano, D.M.; Vásquez-Serna, H.; Jimenez-Canizales, C.E.; Parra-Valencia, E.; Rodriguez-Morales, A.J. Post-Chikungunya Chronic Inflammatory Rheumatism: Follow-up of Cases after 1 Year of Infection in Tolima, Colombia. Travel Med. Infect. Dis. 2018, 21, 62–68. [Google Scholar] [CrossRef]
- Cunha, M.S.; Costa, P.A.G.; Correa, I.A.; de Souza, M.R.M.; Calil, P.T.; da Silva, G.P.D.; Costa, S.M.; Fonseca, V.W.P.; da Costa, L.J. Chikungunya Virus: An Emergent Arbovirus to the South American Continent and a Continuous Threat to the World. Front. Microbiol. 2020, 11, 1297. [Google Scholar] [CrossRef]
- Rashad, A.A.; Mahalingam, S.; Keller, P.A. Chikungunya Virus: Emerging Targets and New Opportunities for Medicinal Chemistry. J. Med. Chem. 2014, 57, 1147–1166. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Simmons, G. Antiviral Functions of Monoclonal Antibodies against Chikungunya Virus. Viruses 2019, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Lum, F.-M.; Ng, L.F.P. Cellular and Molecular Mechanisms of Chikungunya Pathogenesis. Antivir. Res. 2015, 120, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Song, S.; Zhang, L. Recent Progress in Vaccine Development Against Chikungunya Virus. Front. Microbiol. 2019, 10, 2881. [Google Scholar] [CrossRef]
- Saraswat, S.; Athmaram, T.N.; Parida, M.; Agarwal, A.; Saha, A.; Dash, P.K. Expression and Characterization of Yeast Derived Chikungunya Virus Like Particles (CHIK-VLPs) and Its Evaluation as a Potential Vaccine Candidate. PLoS Negl. Trop. Dis. 2016, 10, e0004782. [Google Scholar] [CrossRef] [PubMed]
- Metz, S.W.; Gardner, J.; Geertsema, C.; Le, T.T.; Goh, L.; Vlak, J.M.; Suhrbier, A.; Pijlman, G.P. Effective Chikungunya Virus-like Particle Vaccine Produced in Insect Cells. PLoS Negl. Trop. Dis. 2013, 7, e2124. [Google Scholar] [CrossRef]
- Noranate, N.; Takeda, N.; Chetanachan, P.; Sittisaman, P.; A-nuegoonpipat, A.; Anantapreecha, S. Characterization of Chikungunya Virus-Like Particles. PLoS ONE 2014, 9, e108169. [Google Scholar] [CrossRef]
- Arévalo, M.T.; Huang, Y.; Jones, C.A.; Ross, T.M. Vaccination with a Chikungunya Virus-like Particle Vaccine Exacerbates Disease in Aged Mice. PLoS Negl. Trop. Dis. 2019, 13, e0007316. [Google Scholar] [CrossRef]
- Cook, S.; Holmes, E.C. A Multigene Analysis of the Phylogenetic Relationships among the Flaviviruses (Family: Flaviviridae) and the Evolution of Vector Transmission. Arch. Virol. 2006, 151, 309–325. [Google Scholar] [CrossRef]
- Unni, S.K.; Růžek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese Encephalitis Virus: From Genome to Infectome. Microbes Infect. 2011, 13, 312–321. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging Flaviviruses: The Spread and Resurgence of Japanese Encephalitis, West Nile and Dengue Viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef]
- Sambri, V.; Capobianchi, M.; Charrel, R.; Fyodorova, M.; Gaibani, P.; Gould, E.; Niedrig, M.; Papa, A.; Pierro, A.; Rossini, G.; et al. West Nile Virus in Europe: Emergence, Epidemiology, Diagnosis, Treatment, and Prevention. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, 699–704. [Google Scholar] [CrossRef]
- Artsob, H.; Gubler, D.J.; Enria, D.A.; Morales, M.A.; Pupo, M.; Bunning, M.L.; Dudley, J.P. West Nile Virus in the New World: Trends in the Spread and Proliferation of West Nile Virus in the Western Hemisphere. Zoonoses Public Health 2009, 56, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.B.; Gubler, D.J. West Nile Virus: Epidemiology and Clinical Features of an Emerging Epidemic in the United States. Annu. Rev. Med. 2006, 57, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z.; Halouzka, J. West Nile Fever--a Reemerging Mosquito-Borne Viral Disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Hayes, E.B.; Sejvar, J.J.; Zaki, S.R.; Lanciotti, R.S.; Bode, A.V.; Campbell, G.L. Virology, Pathology, and Clinical Manifestations of West Nile Virus Disease. Emerg. Infect. Dis. 2005, 11, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A Structural Perspective of the Flavivirus Life Cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Martín-Acebes, M.A.; Saiz, J.-C. West Nile Virus: A Re-Emerging Pathogen Revisited. World J. Virol. 2012, 1, 51–70. [Google Scholar] [CrossRef]
- Ulbert, S. West Nile Virus: The Complex Biology of an Emerging Pathogen. Intervirology 2011, 54, 171–184. [Google Scholar] [CrossRef]
- De Filette, M.; Ulbert, S.; Diamond, M.S.; Sanders, N.N. Recent Progress in West Nile Virus Diagnosis and Vaccination. Vet. Res. 2012, 43, 16. [Google Scholar] [CrossRef] [PubMed]
- Nybakken, G.E.; Oliphant, T.; Johnson, S.; Burke, S.; Diamond, M.S.; Fremont, D.H. Structural Basis of West Nile Virus Neutralization by a Therapeutic Antibody. Nature 2005, 437, 764–769. [Google Scholar] [CrossRef]
- Chu, J.J.H.; Ng, M.L. Infectious Entry of West Nile Virus Occurs through a Clathrin-Mediated Endocytic Pathway. J. Virol. 2004, 78, 10543–10555. [Google Scholar] [CrossRef] [PubMed]
- Nybakken, G.E.; Nelson, C.A.; Chen, B.R.; Diamond, M.S.; Fremont, D.H. Crystal Structure of the West Nile Virus Envelope Glycoprotein. J. Virol. 2006, 80, 11467–11474. [Google Scholar] [CrossRef]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. Structure of the Dengue Virus Envelope Protein after Membrane Fusion. Nature 2004, 427, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q. Plant-Made Vaccines against West Nile Virus Are Potent, Safe, and Economically Feasible. Biotechnol. J. 2015, 10, 671–680. [Google Scholar] [CrossRef]
- Lai, H.; Paul, A.M.; Sun, H.; He, J.; Yang, M.; Bai, F.; Chen, Q. A Plant-Produced Vaccine Protects Mice against Lethal West Nile Virus Infection without Enhancing Zika or Dengue Virus Infectivity. Vaccine 2018, 36, 1846–1852. [Google Scholar] [CrossRef]
- He, J.; Lai, H.; Brock, C.; Chen, Q. A Novel System for Rapid and Cost-Effective Production of Detection and Diagnostic Reagents of West Nile Virus in Plants. J. Biomed. Biotechnol. 2011, 2012, e106783. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Peng, L.; Lai, H.; Hurtado, J.; Stahnke, J.; Chen, Q. A Plant-Produced Antigen Elicits Potent Immune Responses against West Nile Virus in Mice. BioMed Res. Int. 2014, 2014, e952865. [Google Scholar] [CrossRef]
- He, J.; Lai, H.; Esqueda, A.; Chen, Q. Plant-Produced Antigen Displaying Virus-Like Particles Evokes Potent Antibody Responses against West Nile Virus in Mice. Vaccines 2021, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Thrane, S.; Janitzek, C.M.; Matondo, S.; Resende, M.; Gustavsson, T.; de Jongh, W.A.; Clemmensen, S.; Roeffen, W.; van de Vegte-Bolmer, M.; van Gemert, G.J.; et al. Bacterial Superglue Enables Easy Development of Efficient Virus-like Particle Based Vaccines. J. Nanobiotechnol. 2016, 14, 30. [Google Scholar] [CrossRef]
- Stander, J.; Chabeda, A.; Rybicki, E.P.; Meyers, A.E. A Plant-Produced Virus-Like Particle Displaying Envelope Protein Domain III Elicits an Immune Response against West Nile Virus in Mice. Front. Plant Sci. 2021, 12, 1915. [Google Scholar] [CrossRef]
- Murugesan, A.; Manoharan, M. Chapter 16—Dengue Virus. In Emerging and Reemerging Viral Pathogens; Ennaji, M.M., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 281–359. ISBN 978-0-12-819400-3. [Google Scholar]
- Bäck, A.T.; Lundkvist, A. Dengue Viruses—An Overview. Infect. Ecol. Epidemiol. 2013, 3, 19839. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Jia, R.; Wang, M.; Yin, Z.; Cheng, A. Structure and Function of Capsid Protein in Flavivirus Infection and Its Applications in the Development of Vaccines and Therapeutics. Vet. Res. 2021, 52, 98. [Google Scholar] [CrossRef]
- Yung, C.-F.; Lee, K.-S.; Thein, T.-L.; Tan, L.-K.; Gan, V.C.; Wong, J.G.X.; Lye, D.C.; Ng, L.-C.; Leo, Y.-S. Dengue Serotype-Specific Differences in Clinical Manifestation, Laboratory Parameters and Risk of Severe Disease in Adults, Singapore. Am. J. Trop. Med. Hyg. 2015, 92, 999–1005. [Google Scholar] [CrossRef]
- Leng, C.-H.; Liu, S.-J.; Tsai, J.-P.; Li, Y.-S.; Chen, M.-Y.; Liu, H.-H.; Lien, S.-P.; Yueh, A.; Hsiao, K.-N.; Lai, L.-W.; et al. A Novel Dengue Vaccine Candidate That Induces Cross-Neutralizing Antibodies and Memory Immunity. Microbes Infect. 2009, 11, 288–295. [Google Scholar] [CrossRef]
- Chen, H.-W.; Liu, S.-J.; Li, Y.-S.; Liu, H.-H.; Tsai, J.-P.; Chiang, C.-Y.; Chen, M.-Y.; Hwang, C.-S.; Huang, C.-C.; Hu, H.-M.; et al. A Consensus Envelope Protein Domain III Can Induce Neutralizing Antibody Responses against Serotype 2 of Dengue Virus in Non-Human Primates. Arch. Virol. 2013, 158, 1523–1531. [Google Scholar] [CrossRef]
- Carlos Marques, L.É.; da Silva, B.B.; Lisboa Magalhães, I.C.; Marques Mendes, M.M.; de Almeida, L.M.; Florindo Guedes, M.I. Production of Dengue 2 Envelope Domain III in Plant Using CPMV—Based Vector System. BMC Proc. 2014, 8, P80. [Google Scholar] [CrossRef]
- Kanagaraj, A.P.; Verma, D.; Daniell, H. Expression of Dengue-3 Premembrane and Envelope Polyprotein in Lettuce Chloroplasts. Plant Mol. Biol. 2011, 76, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.A.; Topal, E.; Giulietti, A.M.; Talou, J.R.; Mason, H. Exploring Different Strategies to Express Dengue Virus Envelope Protein in a Plant System. Biotechnol. Lett. 2010, 32, 867–875. [Google Scholar] [CrossRef]
- Gottschamel, J.; Lössl, A.; Ruf, S.; Wang, Y.; Skaugen, M.; Bock, R.; Clarke, J.L. Production of Dengue Virus Envelope Protein Domain III-Based Antigens in Tobacco Chloroplasts Using Inducible and Constitutive Expression Systems. Plant Mol. Biol. 2016, 91, 497–512. [Google Scholar] [CrossRef] [PubMed]
- van Eerde, A.; Gottschamel, J.; Bock, R.; Hansen, K.E.A.; Munang’andu, H.M.; Daniell, H.; Liu Clarke, J. Production of Tetravalent Dengue Virus Envelope Protein Domain III Based Antigens in Lettuce Chloroplasts and Immunologic Analysis for Future Oral Vaccine Development. Plant Biotechnol. J. 2019, 17, 1408–1417. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Reljic, R.; Kilbourne, J.; Ceballos-Olvera, I.; Yang, M.-S.; Reyes-del Valle, J.; Mason, H.S. Novel Vaccination Approach for Dengue Infection Based on Recombinant Immune Complex Universal Platform. Vaccine 2015, 33, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-Y.; Yang, M.-S.; Kim, T.-G. Expression of Dengue Virus E Glycoprotein Domain III in Non-Nicotine Transgenic Tobacco Plants. Biotechnol. Bioprocess Eng. 2009, 14, 725–730. [Google Scholar] [CrossRef]
- Saejung, W.; Fujiyama, K.; Takasaki, T.; Ito, M.; Hori, K.; Malasit, P.; Watanabe, Y.; Kurane, I.; Seki, T. Production of Dengue 2 Envelope Domain III in Plant Using TMV-Based Vector System. Vaccine 2007, 25, 6646–6654. [Google Scholar] [CrossRef]
- Ponndorf, D.; Meshcheriakova, Y.; Thuenemann, E.C.; Dobon Alonso, A.; Overman, R.; Holton, N.; Dowall, S.; Kennedy, E.; Stocks, M.; Lomonossoff, G.P.; et al. Plant-Made Dengue Virus-like Particles Produced by Co-Expression of Structural and Non-Structural Proteins Induce a Humoral Immune Response in Mice. Plant Biotechnol. J. 2021, 19, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Gorshkov, K.; Shiryaev, S.A.; Fertel, S.; Lin, Y.-W.; Huang, C.-T.; Pinto, A.; Farhy, C.; Strongin, A.Y.; Zheng, W.; Terskikh, A.V. Zika Virus: Origins, Pathological Action, and Treatment Strategies. Front. Microbiol. 2019, 9, 3252. [Google Scholar] [CrossRef]
- Gregory, C.J.; Oduyebo, T.; Brault, A.C.; Brooks, J.T.; Chung, K.-W.; Hills, S.; Kuehnert, M.J.; Mead, P.; Meaney-Delman, D.; Rabe, I.; et al. Modes of Transmission of Zika Virus. J. Infect. Dis. 2017, 216, S875–S883. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, E.; Orovou, E.; Sarella, A.; Iliadou, M.; Rigas, N.; Palaska, E.; Iatrakis, G.; Dagla, M. Zika Virus and the Risk of Developing Microcephaly in Infants: A Systematic Review. Int. J. Environ. Res. Public Health 2020, 17, 3806. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, T.V.B.; Rodrigues, L.C.; de Alencar Ximenes, R.A.; de Barros Miranda-Filho, D.; Montarroyos, U.R.; de Melo, A.P.L.; Valongueiro, S.; de Albuquerque, M.d.F.P.M.; Souza, W.V.; Braga, C.; et al. Association between Zika Virus Infection and Microcephaly in Brazil, January to May 2016: Preliminary Report of a Case-Control Study. Lancet Infect. Dis. 2016, 16, 1356–1363. [Google Scholar] [CrossRef]
- Paniz-Mondolfi, A.E.; Blohm, G.M.; Hernandez-Perez, M.; Larrazabal, A.; Moya, D.; Marquez, M.; Talamo, A.; Carrillo, A.; Rothe de Arocha, J.; Lednicky, J.; et al. Cutaneous Features of Zika Virus Infection: A Clinicopathological Overview. Clin. Exp. Dermatol. 2019, 44, 13–19. [Google Scholar] [CrossRef]
- Faye, O.; Freire, C.C.M.; Iamarino, A.; Faye, O.; de Oliveira, J.V.C.; Diallo, M.; Zanotto, P.M.A.; Sall, A.A. Molecular Evolution of Zika Virus during Its Emergence in the 20th Century. PLoS Negl. Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. The Continued Threat of Emerging Flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Valiant, W.G.; Huang, Y.-J.S.; Vanlandingham, D.L.; Higgs, S.; Lewis, M.G.; Mattapallil, J.J. Zika Convalescent Macaques Display Delayed Induction of Anamnestic Cross-Neutralizing Antibody Responses after Dengue Infection. Emerg. Microbes Infect. 2018, 7, 1–11. [Google Scholar] [CrossRef]
- Chambers, T.; Monath, T. The Flaviviruses: Structure, Replication and Evolution, 1st ed.; Academic Press: Cambridge, MA, USA, 2003; ISBN 9780080493817. [Google Scholar]
- Wang, L.; Wang, R.; Wang, L.; Ben, H.; Yu, L.; Gao, F.; Shi, X.; Yin, C.; Zhang, F.; Xiang, Y.; et al. Structural Basis for Neutralization and Protection by a Zika Virus-Specific Human Antibody. Cell Rep. 2019, 26, 3360–3368. [Google Scholar] [CrossRef]
- Yang, M.; Sun, H.; Lai, H.; Hurtado, J.; Chen, Q. Plant-Produced Zika Virus Envelope Protein Elicits Neutralizing Immune Responses That Correlate with Protective Immunity against Zika Virus in Mice. Plant Biotechnol. J. 2018, 16, 572–580. [Google Scholar] [CrossRef]
- Yang, M.; Lai, H.; Sun, H.; Chen, Q. Virus-like Particles That Display Zika Virus Envelope Protein Domain III Induce Potent Neutralizing Immune Responses in Mice. Sci. Rep. 2017, 7, 7679. [Google Scholar] [CrossRef]
- Diamos, A.G.; Pardhe, M.D.; Sun, H.; Hunter, J.G.L.; Kilbourne, J.; Chen, Q.; Mason, H.S. A Highly Expressing, Soluble, and Stable Plant-Made IgG Fusion Vaccine Strategy Enhances Antigen Immunogenicity in Mice Without Adjuvant. Front. Immunol. 2020, 11, 3140. [Google Scholar] [CrossRef]
- Diamos, A.G.; Pardhe, M.D.; Sun, H.; Hunter, J.G.L.; Mor, T.; Meador, L.; Kilbourne, J.; Chen, Q.; Mason, H.S. Codelivery of Improved Immune Complex and Virus-like Particle Vaccines Containing Zika Virus Envelope Domain III Synergistically Enhances Immunogenicity. Vaccine 2020, 38, 3455–3463. [Google Scholar] [CrossRef]
- Jentes, E.S.; Poumerol, G.; Gershman, M.D.; Hill, D.R.; Lemarchand, J.; Lewis, R.F.; Staples, J.E.; Tomori, O.; Wilder-Smith, A.; Monath, T.P. The Revised Global Yellow Fever Risk Map and Recommendations for Vaccination, 2010: Consensus of the Informal WHO Working Group on Geographic Risk for Yellow Fever. Lancet Infect. Dis. 2011, 11, 622–632. [Google Scholar] [CrossRef]
- WHO Yellow Fever. Available online: https://www.who.int/news-room/fact-sheets/detail/yellow-fever (accessed on 20 November 2021).
- Jones, C.T.; Patkar, C.G.; Kuhn, R.J. Construction and Applications of Yellow Fever Virus Replicons. Virology 2005, 331, 247–259. [Google Scholar] [CrossRef]
- Tottey, S.; Shoji, Y.; Jones, R.M.; Chichester, J.A.; Green, B.J.; Musiychuk, K.; Si, H.; Manceva, S.D.; Rhee, A.; Shamloul, M.; et al. Plant-Produced Subunit Vaccine Candidates against Yellow Fever Induce Virus Neutralizing Antibodies and Confer Protection against Viral Challenge in Animal Models. Am. J. Trop. Med. Hyg. 2017, 98, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.A.; Barrett, A.D.T. The Present and Future of Yellow Fever Vaccines. Pharmaceuticals 2021, 14, 891. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and Geographical Expansion of Japanese Encephalitis Virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, H.; Mori, C.; Fuke, I.; Morita, K.; Kuhara, S.; Kondou, J.; Kikuchi, Y.; Nagamatu, H.; Igarashi, A. Complete Nucleotide Sequence of the Japanese Encephalitis Virus Genome RNA. Virology 1987, 161, 497–510. [Google Scholar] [CrossRef]
- Xu, X.-G.; Wang, Z.-S.; Zhang, Q.; Li, Z.-C.; Zhao, H.-N.; Li, W.; Tong, D.-W.; Liu, H.-J. Baculovirus Surface Display of E Envelope Glycoprotein of Japanese Encephalitis Virus and Its Immunogenicity of the Displayed Proteins in Mouse and Swine Models. Vaccine 2011, 29, 636–643. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, H.; Zhang, X.; Xiao, H.; Jiang, Y.; Song, Y.; Fang, L.; Xiao, S.; Zhen, Y.; Chen, H. Generation and Immunogenicity of Japanese Encephalitis Virus Envelope Protein Expressed in Transgenic Rice. Biochem. Biophys. Res. Commun. 2009, 380, 292–297. [Google Scholar] [CrossRef]
- Chen, T.-H.; Hu, C.-C.; Liao, J.-T.; Lee, Y.-L.; Huang, Y.-W.; Lin, N.-S.; Lin, Y.-L.; Hsu, Y.-H. Production of Japanese Encephalitis Virus Antigens in Plants Using Bamboo Mosaic Virus-Based Vector. Front. Microbiol. 2017, 8, 788. [Google Scholar] [CrossRef] [PubMed]
- Valarcher, J.F.; Hägglund, S.; Juremalm, M.; Blomqvist, G.; Renström, L.; Zohari, S.; Leijon, M.; Chirico, J. Tick-Borne Encephalitis. Rev. Sci. Tech. Int. Off. Epizoot. 2015, 34, 453–466. [Google Scholar] [CrossRef]
- Lindquist, L.; Vapalahti, O. Tick-Borne Encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T.Y. 2009 Tick-Borne Encephalitis Virus—A Review of an Emerging Zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef] [PubMed]
- Jääskeläinen, A.; Han, X.; Niedrig, M.; Vaheri, A.; Vapalahti, O. Diagnosis of Tick-Borne Encephalitis by a μ-Capture Immunoglobulin M-Enzyme Immunoassay Based on Secreted Recombinant Antigen Produced in Insect Cells. J. Clin. Microbiol. 2003, 41, 4336–4342. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.L.; Stadler, K.; Mandl, C.W.; Kunz, C.; Heinz, F.X. Synthesis and Secretion of Recombinant Tick-Borne Encephalitis Virus Protein E in Soluble and Particulate Form. J. Virol. 1995, 69, 5816–5820. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Allison, S.L.; Stiasny, K.; Schalich, J.; Holzmann, H.; Mandl, C.W.; Kunz, C. Recombinant and Virion-Derived Soluble and Particulate Immunogens for Vaccination against Tick-Borne Encephalitis. Vaccine 1995, 13, 1636–1642. [Google Scholar] [CrossRef]
- Andersson, S.; Asghar, N.; Davidsson, Å.; Gunaltay, S.; Johansson, M.; Melik, W.; Persson, A.; Särndahl, E.; Tran, H. Molecular Flavivirus Research—Örebro University. Available online: https://www.oru.se/english/research/research-teams/rt/?rdb=g225 (accessed on 20 November 2021).
- Barré-Sinoussi, F.; Chermann, J.C.; Rey, F.; Nugeyre, M.T.; Chamaret, S.; Gruest, J.; Dauguet, C.; Axler-Blin, C.; Vézinet-Brun, F.; Rouzioux, C.; et al. Isolation of a T-Lymphotropic Retrovirus from a Patient at Risk for Acquired Immune Deficiency Syndrome (AIDS). Science 1983, 220, 868–871. [Google Scholar] [CrossRef]
- Gallo, R.C.; Salahuddin, S.Z.; Popovic, M.; Shearer, G.M.; Kaplan, M.; Haynes, B.F.; Palker, T.J.; Redfield, R.; Oleske, J.; Safai, B. Frequent Detection and Isolation of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and at Risk for AIDS. Science 1984, 224, 500–503. [Google Scholar] [CrossRef]
- Huet, T.; Cheynier, R.; Meyerhans, A.; Roelants, G.; Wain-Hobson, S. Genetic Organization of a Chimpanzee Lentivirus Related to HIV-1. Nature 1990, 345, 356–359. [Google Scholar] [CrossRef]
- Hirsch, V.M.; Olmsted, R.A.; Murphey-Corb, M.; Purcell, R.H.; Johnson, P.R. An African Primate Lentivirus SIVsmclosely Related to HIV-2. Nature 1989, 339, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS Pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [Google Scholar] [CrossRef] [PubMed]
- De Cock, K.M.; Adjorlolo, G.; Ekpini, E.; Sibailly, T.; Kouadio, J.; Maran, M.; Brattegaard, K.; Vetter, K.M.; Doorly, R.; Gayle, H.D. Epidemiology and Transmission of HIV-2: Why There Is No HIV-2 Pandemic. JAMA 1993, 270, 2083–2086. [Google Scholar] [CrossRef] [PubMed]
- van Schooten, J.; van Gils, M.J. HIV-1 Immunogens and Strategies to Drive Antibody Responses towards Neutralization Breadth. Retrovirology 2018, 15, 74. [Google Scholar] [CrossRef] [PubMed]
- Tremouillaux-Guiller, J.; Moustafa, K.; Hefferon, K.; Gaobotse, G.; Makhzoum, A. Plant-Made HIV Vaccines and Potential Candidates. Curr. Opin. Biotechnol. 2020, 61, 209–216. [Google Scholar] [CrossRef]
- Scotti, N.; Buonaguro, L.; Tornesello, M.L.; Cardi, T.; Buonaguro, F.M. Plant-Based Anti-HIV-1 Strategies: Vaccine Molecules and Antiviral Approaches. Expert Rev. Vaccines 2010, 9, 925–936. [Google Scholar] [CrossRef]
- Zolla-Pazner, S. Identifying Epitopes of HIV-1 That Induce Protective Antibodies. Nat. Rev. Immunol. 2004, 4, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Govea-Alonso, D.O.; Gómez-Cardona, E.E.; Rubio-Infante, N.; García-Hernández, A.L.; Varona-Santos, J.T.; Salgado-Bustamante, M.; Korban, S.S.; Moreno-Fierros, L.; Rosales-Mendoza, S. Production of an Antigenic C4(V3)6 Multiepitopic HIV Protein in Bacterial and Plant Systems. Plant Cell Tissue Organ Cult. 2013, 113, 73–79. [Google Scholar] [CrossRef]
- Lindh, I.; Kalbina, I.; Thulin, S.; Scherbak, N.; Sävenstrand, H.; Bråve, A.; Hinkula, J.; Strid, Å.; Andersson, S. Feeding of Mice with Arabidopsis Thaliana Expressing the HIV-1 Subtype C P24 Antigen Gives Rise to Systemic Immune Responses. APMIS 2008, 116, 985–994. [Google Scholar] [CrossRef]
- Gonzalez-Rabade, N.; McGowan, E.G.; Zhou, F.; McCabe, M.S.; Bock, R.; Dix, P.J.; Gray, J.C.; Ma, J.K.-C. Immunogenicity of Chloroplast-Derived HIV-1 P24 and a P24-Nef Fusion Protein Following Subcutaneous and Oral Administration in Mice. Plant Biotechnol. J. 2011, 9, 629–638. [Google Scholar] [CrossRef]
- Pérez-Filgueira, D.M.; Brayfield, B.P.; Phiri, S.; Borca, M.V.; Wood, C.; Morris, T.J. Preserved Antigenicity of HIV-1 P24 Produced and Purified in High Yields from Plants Inoculated with a Tobacco Mosaic Virus (TMV)-Derived Vector. J. Virol. Methods 2004, 121, 201–208. [Google Scholar] [CrossRef]
- McLain, L.; Durrani, Z.; Wisniewski, L.A.; Porta, C.; Lomonossoff, G.P.; Dimmock, N.J. Stimulation of Neutralizing Antibodies to Human Immunodeficiency Virus Type 1 in Three Strains of Mice Immunized with a 22 Amino Acid Peptide of Gp41 Expressed on the Surface of a Plant Virus. Vaccine 1996, 14, 799–810. [Google Scholar] [CrossRef]
- Guetard, D.; Greco, R.; Cervantes Gonzalez, M.; Celli, S.; Kostrzak, A.; Langlade-Demoyen, P.; Sala, F.; Wain-Hobson, S.; Sala, M. Immunogenicity and Tolerance Following HIV-1/HBV Plant-Based Oral Vaccine Administration. Vaccine 2008, 26, 4477–4485. [Google Scholar] [CrossRef]
- Greco, R.; Michel, M.; Guetard, D.; Cervantes-Gonzalez, M.; Pelucchi, N.; Wain-Hobson, S.; Sala, F.; Sala, M. Production of Recombinant HIV-1/HBV Virus-like Particles in Nicotiana Tabacum and Arabidopsis Thaliana Plants for a Bivalent Plant-Based Vaccine. Vaccine 2007, 25, 8228–8240. [Google Scholar] [CrossRef]
- Margolin, E.; Oh, Y.J.; Verbeek, M.; Naude, J.; Ponndorf, D.; Meshcheriakova, Y.A.; Peyret, H.; van Diepen, M.T.; Chapman, R.; Meyers, A.E.; et al. Co-expression of Human Calreticulin Significantly Improves the Production of HIV Gp140 and Other Viral Glycoproteins in Plants. Plant Biotechnol. J. 2020, 18, 2109–2117. [Google Scholar] [CrossRef] [PubMed]
- Margolin, E.; Chapman, R.; Meyers, A.E.; van Diepen, M.T.; Ximba, P.; Hermanus, T.; Crowther, C.; Weber, B.; Morris, L.; Williamson, A.-L.; et al. Production and Immunogenicity of Soluble Plant-Produced HIV-1 Subtype C Envelope Gp140 Immunogens. Front. Plant Sci. 2019, 10, 1378. [Google Scholar] [CrossRef]
- Porta, C.; Spall, V.E.; Loveland, J.; Johnson, J.E.; Barker, P.J.; Lomonossoff, G.P. Development of Cowpea Mosaic Virus as a High-Yielding System for the Presentation of Foreign Peptides. Virology 1994, 202, 949–955. [Google Scholar] [CrossRef] [PubMed]
- McLAIN, L.; Porta, C.; Lomonossoff, G.P.; Durrani, Z.; Dimmock, N.J. Human Immunodeficiency Virus Type 1-Neutralizing Antibodies Raised to a Glycoprotein 41 Peptide Expressed on the Surface of a Plant Virus. AIDS Res. Hum. Retrovir. 1995, 11, 327–334. [Google Scholar] [CrossRef]
- Matoba, N.; Magérus, A.; Geyer, B.C.; Zhang, Y.; Muralidharan, M.; Alfsen, A.; Arntzen, C.J.; Bomsel, M.; Mor, T.S. A Mucosally Targeted Subunit Vaccine Candidate Eliciting HIV-1 Transcytosis-Blocking Abs. Proc. Natl. Acad. Sci. USA 2004, 101, 13584–13589. [Google Scholar] [CrossRef]
- Shchelkunov, S.N.; Salyaev, R.K.; Pozdnyakov, S.G.; Rekoslavskaya, N.I.; Nesterov, A.E.; Ryzhova, T.S.; Sumtsova, V.M.; Pakova, N.V.; Mishutina, U.O.; Kopytina, T.V.; et al. Immunogenicity of a Novel, Bivalent, Plant-Based Oral Vaccine against Hepatitis B and Human Immunodeficiency Viruses. Biotechnol. Lett. 2006, 28, 959–967. [Google Scholar] [CrossRef]
- Ramírez, Y.J.P.; Tasciotti, E.; Gutierrez-Ortega, A.; Torres, A.J.D.; Flores, M.T.O.; Giacca, M.; Lim, M.Á.G. Fruit-Specific Expression of the Human Immunodeficiency Virus Type 1 Tat Gene in Tomato Plants and Its Immunogenic Potential in Mice. Clin. Vaccine Immunol. 2007, 14, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Meyers, A.; Chakauya, E.; Shephard, E.; Tanzer, F.L.; Maclean, J.; Lynch, A.; Williamson, A.-L.; Rybicki, E.P. Expression of HIV-1 Antigens in Plants as Potential Subunit Vaccines. BMC Biotechnol. 2008, 8, 53. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Kajiura, H.; Cherni, I.; Doran, J.D.; Bomsel, M.; Fujiyama, K.; Mor, T.S. Biochemical and Immunological Characterization of the Plant-Derived Candidate Human Immunodeficiency Virus Type 1 Mucosal Vaccine CTB–MPR649–684. Plant Biotechnol. J. 2009, 7, 129–145. [Google Scholar] [CrossRef]
- Lindh, I.; Bråve, A.; Hallengärd, D.; Hadad, R.; Kalbina, I.; Strid, Å.; Andersson, S. Oral Delivery of Plant-Derived HIV-1 P24 Antigen in Low Doses Shows a Superior Priming Effect in Mice Compared to High Doses. Vaccine 2014, 32, 2288–2293. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Infante, N.; Govea-Alonso, D.O.; Romero-Maldonado, A.; García-Hernández, A.L.; Ilhuicatzi-Alvarado, D.; Salazar-González, J.A.; Korban, S.S.; Rosales-Mendoza, S.; Moreno-Fierros, L. A Plant-Derived Multi-HIV Antigen Induces Broad Immune Responses in Orally Immunized Mice. Mol. Biotechnol. 2015, 57, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; Rubio-Infante, N.; Monreal-Escalante, E.; Govea-Alonso, D.O.; García-Hernández, A.L.; Salazar-González, J.A.; González-Ortega, O.; Paz-Maldonado, L.M.T.; Moreno-Fierros, L. Chloroplast Expression of an HIV Envelop-Derived Multiepitope Protein: Towards a Multivalent Plant-Based Vaccine. Plant Cell Tissue Organ Cult. 2014, 116, 111–123. [Google Scholar] [CrossRef]
- Orellana-Escobedo, L.; Rosales-Mendoza, S.; Romero-Maldonado, A.; Parsons, J.; Decker, E.L.; Monreal-Escalante, E.; Moreno-Fierros, L.; Reski, R. An Env-Derived Multi-Epitope HIV Chimeric Protein Produced in the Moss Physcomitrella Patens Is Immunogenic in Mice. Plant Cell Rep. 2015, 34, 425–433. [Google Scholar] [CrossRef]
- Kessans, S.A.; Linhart, M.D.; Matoba, N.; Mor, T. Biological and Biochemical Characterization of HIV-1 Gag/Dgp41 Virus-like Particles Expressed in Nicotiana Benthamiana. Plant Biotechnol. J. 2013, 11, 681–690. [Google Scholar] [CrossRef]
- Kessans, S.A.; Linhart, M.D.; Meador, L.R.; Kilbourne, J.; Hogue, B.G.; Fromme, P.; Matoba, N.; Mor, T.S. Immunological Characterization of Plant-Based HIV-1 Gag/Dgp41 Virus-Like Particles. PLoS ONE 2016, 11, e0151842. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.E.; Dolin, R.; Blaser, M.J. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-323-55027-7. [Google Scholar]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Primer 2017, 3, 1–19. [Google Scholar] [CrossRef]
- Julia Paoli Is Rabies Really 100% Fatal? Viruses101. Learn Science at Scitable. Available online: https://www.nature.com/scitable/blog/viruses101/is_rabies_really_100_fatal/ (accessed on 20 November 2021).
- Albertini, A.A.V.; Ruigrok, R.W.H.; Blondel, D. Rabies Virus Transcription and Replication. Adv. Virus Res. 2011, 79, 1–22. [Google Scholar] [CrossRef]
- Takeyama, N.; Kiyono, H.; Yuki, Y. Plant-Based Vaccines for Animals and Humans: Recent Advances in Technology and Clinical Trials. Ther. Adv. Vaccines 2015, 3, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Loza-Rubio, E.; Rojas-Anaya, E. Edible Rabies Vaccines. In Commercial Plant-Produced Recombinant Protein Products: Case Studies; Howard, J.A., Hood, E.E., Eds.; Biotechnology in Agriculture and Forestry; Springer: Berlin/Heidelberg, Germany, 2014; pp. 153–177. ISBN 978-3-662-43836-7. [Google Scholar]
- Ashraf, S.; Singh, P.K.; Yadav, D.K.; Shahnawaz, M.; Mishra, S.; Sawant, S.V.; Tuli, R. High Level Expression of Surface Glycoprotein of Rabies Virus in Tobacco Leaves and Its Immunoprotective Activity in Mice. J. Biotechnol. 2005, 119, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Srivastava, S.; Chouksey, A.; Panwar, B.S.; Verma, P.C.; Roy, S.; Singh, P.K.; Saxena, G.; Tuli, R. Expression of Rabies Glycoprotein and Ricin Toxin B Chain (RGP–RTB) Fusion Protein in Tomato Hairy Roots: A Step Towards Oral Vaccination for Rabies. Mol. Biotechnol. 2015, 57, 359–370. [Google Scholar] [CrossRef]
- Perea Arango, I.; Loza Rubio, E.; Rojas Anaya, E.; Olivera Flores, T.; de la Vara, L.G.; Gómez Lim, M.A. Expression of the Rabies Virus Nucleoprotein in Plants at High-Levels and Evaluation of Immune Responses in Mice. Plant Cell Rep. 2008, 27, 677–685. [Google Scholar] [CrossRef] [PubMed]
- McGarvey, P.B.; Hammond, J.; Dienelt, M.M.; Hooper, D.C.; Fu, Z.F.; Dietzschold, B.; Koprowski, H.; Michaels, F.H. Expression of the Rabies Virus Glycoprotein in Transgenic Tomatoes. Biotechnology 1995, 13, 1484–1487. [Google Scholar] [CrossRef]
- Rojas-Anaya, E.; Loza-Rubio, E.; Olivera-Flores, M.T.; Gomez-Lim, M. Expression of Rabies Virus G Protein in Carrots (Daucus Carota). Transgenic Res. 2009, 18, 911. [Google Scholar] [CrossRef] [PubMed]
- Modelska, A.; Dietzschold, B.; Sleysh, N.; Fu, Z.F.; Steplewski, K.; Hooper, D.C.; Koprowski, H.; Yusibov, V. Immunization against Rabies with Plant-Derived Antigen. Proc. Natl. Acad. Sci. USA 1998, 95, 2481–2485. [Google Scholar] [CrossRef]
- Yusibov, V.; Hooper, D.C.; Spitsin, S.V.; Fleysh, N.; Kean, R.B.; Mikheeva, T.; Deka, D.; Karasev, A.; Cox, S.; Randall, J.; et al. Expression in Plants and Immunogenicity of Plant Virus-Based Experimental Rabies Vaccine. Vaccine 2002, 20, 3155–3164. [Google Scholar] [CrossRef]
- Loza-Rubio, E.; Rojas, E.; Gómez, L.; Olivera, M.T.J.; Gómez-Lim, M.A. Development of an Edible Rabies Vaccine in Maize Using the Vnukovo Strain. Dev. Biol. 2008, 131, 477–482. [Google Scholar]
- Loza-Rubio, E.; Rojas-Anaya, E.; López, J.; Olivera-Flores, M.T.; Gómez-Lim, M.; Tapia-Pérez, G. Induction of a Protective Immune Response to Rabies Virus in Sheep after Oral Immunization with Transgenic Maize, Expressing the Rabies Virus Glycoprotein. Vaccine 2012, 30, 5551–5556. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.-J. Structural and Molecular Biology of Hepatitis E Virus. Comput. Struct. Biotechnol. J. 2021, 19, 1907–1916. [Google Scholar] [CrossRef]
- Yugo, D.M.; Meng, X.-J. Hepatitis E Virus: Foodborne, Waterborne and Zoonotic Transmission. Int. J. Environ. Res. Public Health 2013, 10, 4507–4533. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Origin and Dispersal of Hepatitis E Virus. Emerg. Microbes Infect. 2018, 7, 11. [Google Scholar] [CrossRef]
- Takova, K.; Koynarski, T.; Minkov, I.; Ivanova, Z.; Toneva, V.; Zahmanova, G. Increasing Hepatitis E Virus Seroprevalence in Domestic Pigs and Wild Boar in Bulgaria. Animals 2020, 10, 1521. [Google Scholar] [CrossRef]
- Tsachev, I.; Baymakova, M.; Pepovich, R.; Palova, N.; Marutsov, P.; Gospodinova, K.; Kundurzhiev, T.; Ciccozzi, M. High Seroprevalence of Hepatitis E Virus Infection Among East Balkan Swine (Sus Scrofa) in Bulgaria: Preliminary Results. Pathogens 2020, 9, 911. [Google Scholar] [CrossRef]
- Tsachev, I.; Baymakova, M.; Marutsov, P.; Gospodinova, K.; Kundurzhiev, T.; Petrov, V.; Pepovich, R. Seroprevalence of Hepatitis E Virus Infection Among Wild Boars in Western Bulgaria. Vector Borne Zoonotic Dis. 2021, 21, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Holla, R.P.; Jameel, S. Molecular Virology of Hepatitis E Virus. Virus Res. 2011, 161, 47–58. [Google Scholar] [CrossRef]
- Ma, Y.; Lin, S.-Q.; Gao, Y.; Li, M.; Luo, W.-X.; Zhang, J.; Xia, N.-S. Expression of ORF2 Partial Gene of Hepatitis E Virus in Tomatoes and Immunoactivity of Expression Products. World J. Gastroenterol. 2003, 9, 2211–2215. [Google Scholar] [CrossRef]
- Maloney, B.J.; Takeda, N.; Suzaki, Y.; Ami, Y.; Li, T.C.; Miyamura, T.; Arntzen, C.J.; Mason, H.S. Challenges in Creating a Vaccine to Prevent Hepatitis E. Vaccine 2005, 23, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-X.; Lee, M.Y.-T.; Ng, J.M.-H.; Chye, M.-L.; Yip, W.-K.; Zee, S.-Y.; Lam, E. A Truncated Hepatitis E Virus ORF2 Protein Expressed in Tobacco Plastids Is Immunogenic in Mice. World J. Gastroenterol. 2006, 12, 306–312. [Google Scholar] [CrossRef][Green Version]
- Mazalovska, M.; Varadinov, N.; Koynarski, T.; Minkov, I.; Teoharov, P.; Lomonossoff, G.P.; Zahmanova, G. Detection of Serum Antibodies to Hepatitis E Virus Based on HEV Genotype 3 ORF2 Capsid Protein Expressed in Nicotiana Benthamiana. Ann. Lab. Med. 2017, 37, 313–319. [Google Scholar] [CrossRef][Green Version]
- Mardanova, E.; Takova, K.; Toneva, V.; Zahmanova, G.; Tsybalova, L.; Ravin, N. A Plant-Based Transient Expression System for the Rapid Production of Highly Immunogenic Hepatitis E Virus-like Particles. Biotechnol. Lett. 2020, 42, 2441–2446. [Google Scholar] [CrossRef] [PubMed]
- Takova, K.; Koynarski, T.; Minkov, G.; Toneva, V.; Mardanova, E.; Ravin, N.; Lukov, G.L.; Zahmanova, G. Development and Optimization of an Enzyme Immunoassay to Detect Serum Antibodies against the Hepatitis E Virus in Pigs, Using Plant-Derived ORF2 Recombinant Protein. Vaccines 2021, 9, 991. [Google Scholar] [CrossRef]
- Gogoi, P.; Ganar, K.; Kumar, S. Avian Paramyxovirus: A Brief Review. Transbound. Emerg. Dis. 2017, 64, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nayak, B.; Collins, P.L.; Samal, S.K. Evaluation of the Newcastle Disease Virus F and HN Proteins in Protective Immunity by Using a Recombinant Avian Paramyxovirus Type 3 Vector in Chickens. J. Virol. 2011, 85, 6521–6534. [Google Scholar] [CrossRef]
- Cox, R.M.; Plemper, R.K. Structure and Organization of Paramyxovirus Particles. Curr. Opin. Virol. 2017, 24, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.R.; Bevins, S.N. A Review of Virulent Newcastle Disease Viruses in the United States and the Role of Wild Birds in Viral Persistence and Spread. Vet. Res. 2017, 48, 68. [Google Scholar] [CrossRef]
- Fox, J.G.; Otto, G.; Colby, L.A. Chapter 28—Selected Zoonoses. In Laboratory Animal Medicine, 3rd ed.; Fox, J.G., Anderson, L.C., Otto, G.M., Pritchett-Corning, K.R., Whary, M.T., Eds.; American College of Laboratory Animal Medicine; Academic Press: Boston, MA, USA, 2015; pp. 1313–1370. ISBN 978-0-12-409527-4. [Google Scholar]
- Alexander, D.J. Gordon Memorial Lecture. Newcastle Disease. Br. Poult. Sci. 2001, 42, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Berinstein, A.; Vazquez-Rovere, C.; Asurmendi, S.; Gómez, E.; Zanetti, F.; Zabal, O.; Tozzini, A.; Conte Grand, D.; Taboga, O.; Calamante, G.; et al. Mucosal and Systemic Immunization Elicited by Newcastle Disease Virus (NDV) Transgenic Plants as Antigens. Vaccine 2005, 23, 5583–5589. [Google Scholar] [CrossRef]
- Yang, Z.-Q.; Liu, Q.-Q.; Pan, Z.-M.; Yu, H.-X.; Jiao, X.-A. Expression of the Fusion Glycoprotein of Newcastle Disease Virus in Transgenic Rice and Its Immunogenicity in Mice. Vaccine 2007, 25, 591–598. [Google Scholar] [CrossRef]
- Guerrero-Andrade, O.; Loza-Rubio, E.; Olivera-Flores, T.; Fehérvári-Bone, T.; Gómez-Lim, M.A. Expression of the Newcastle Disease Virus Fusion Protein in Transgenic Maize and Immunological Studies. Transgenic Res. 2006, 15, 455–463. [Google Scholar] [CrossRef]
- Shahid, N.; Samiullah, T.R.; Shakoor, S.; Latif, A.; Yasmeen, A.; Azam, S.; Shahid, A.A.; Husnain, T.; Rao, A.Q. Early Stage Development of a Newcastle Disease Vaccine Candidate in Corn. Front. Vet. Sci. 2020, 7, 499. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, M.J.; Ebrahimi, M.M.; Shahsavandi, S.; Amani, J.; Kazemi, R.; Jafari, M.; Salmanian, A.-H. The Immunogenicity of a Novel Chimeric Hemagglutinin-Neuraminidase-Fusion Antigen from Newcastle Disease Virus by Oral Delivery of Transgenic Canola Seeds to Chickens. Mol. Biotechnol. 2020, 62, 344–354. [Google Scholar] [CrossRef]
- Natilla, A.; Hammond, R.W.; Nemchinov, L.G. Epitope Presentation System Based on Cucumber Mosaic Virus Coat Protein Expressed from a Potato Virus X-Based Vector. Arch. Virol. 2006, 151, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Natilla, A.; Nemchinov, L.G. Improvement of PVX/CMV CP Expression Tool for Display of Short Foreign Antigens. Protein Expr. Purif. 2008, 59, 117–121. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gómez, E.; Zoth, S.C.; Asurmendi, S.; Vázquez Rovere, C.; Berinstein, A. Expression of Hemagglutinin-Neuraminidase Glycoprotein of Newcastle Disease Virus in Agroinfiltrated Nicotiana Benthamiana Plants. J. Biotechnol. 2009, 144, 337–340. [Google Scholar] [CrossRef]
- Hahn, B.-S.; Jeon, I.-S.; Jung, Y.-J.; Kim, J.-B.; Park, J.-S.; Ha, S.-H.; Kim, K.-H.; Kim, H.-M.; Yang, J.-S.; Kim, Y.-H. Expression of Hemagglutinin-Neuraminidase Protein of Newcastle Disease Virus in Transgenic Tobacco. Plant Biotechnol. Rep. 2007, 1, 85–92. [Google Scholar] [CrossRef]
- Vermij, P. News In Brief. Nat. Biotechnol. 2006, 24, 233–234. [Google Scholar] [CrossRef]
- Eaton, B.T.; Broder, C.C.; Middleton, D.; Wang, L.-F. Hendra and Nipah Viruses: Different and Dangerous. Nat. Rev. Microbiol. 2006, 4, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Clayton, B.A.; Wang, L.F.; Marsh, G.A. Henipaviruses: An Updated Review Focusing on the Pteropid Reservoir and Features of Transmission. Zoonoses Public Health 2013, 60, 69–83. [Google Scholar] [CrossRef]
- Islam, M.S.; Sazzad, H.M.S.; Satter, S.M.; Sultana, S.; Hossain, M.J.; Hasan, M.; Rahman, M.; Campbell, S.; Cannon, D.L.; Ströher, U.; et al. Nipah Virus Transmission from Bats to Humans Associated with Drinking Traditional Liquor Made from Date Palm Sap, Bangladesh, 2011–2014. Emerg. Infect. Dis. J. 2016, 22, 664. [Google Scholar] [CrossRef] [PubMed]
- Enchéry, F.; Horvat, B. Understanding the Interaction between Henipaviruses and Their Natural Host, Fruit Bats: Paving the Way toward Control of Highly Lethal Infection in Humans. Int. Rev. Immunol. 2017, 36, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Amaya, M.; Broder, C.C. Vaccines to Emerging Viruses: Nipah and Hendra. Annu. Rev. Virol. 2020, 7, 447–473. [Google Scholar] [CrossRef] [PubMed]



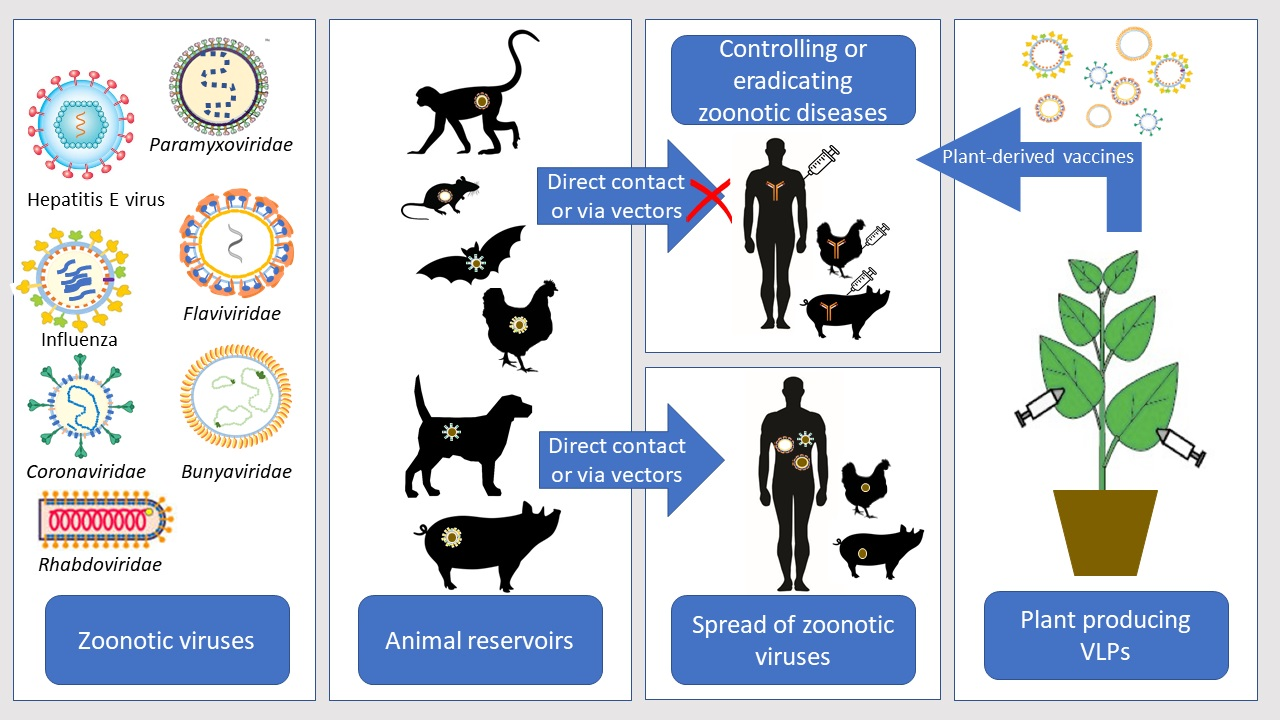{kind=link}
{kind=link}
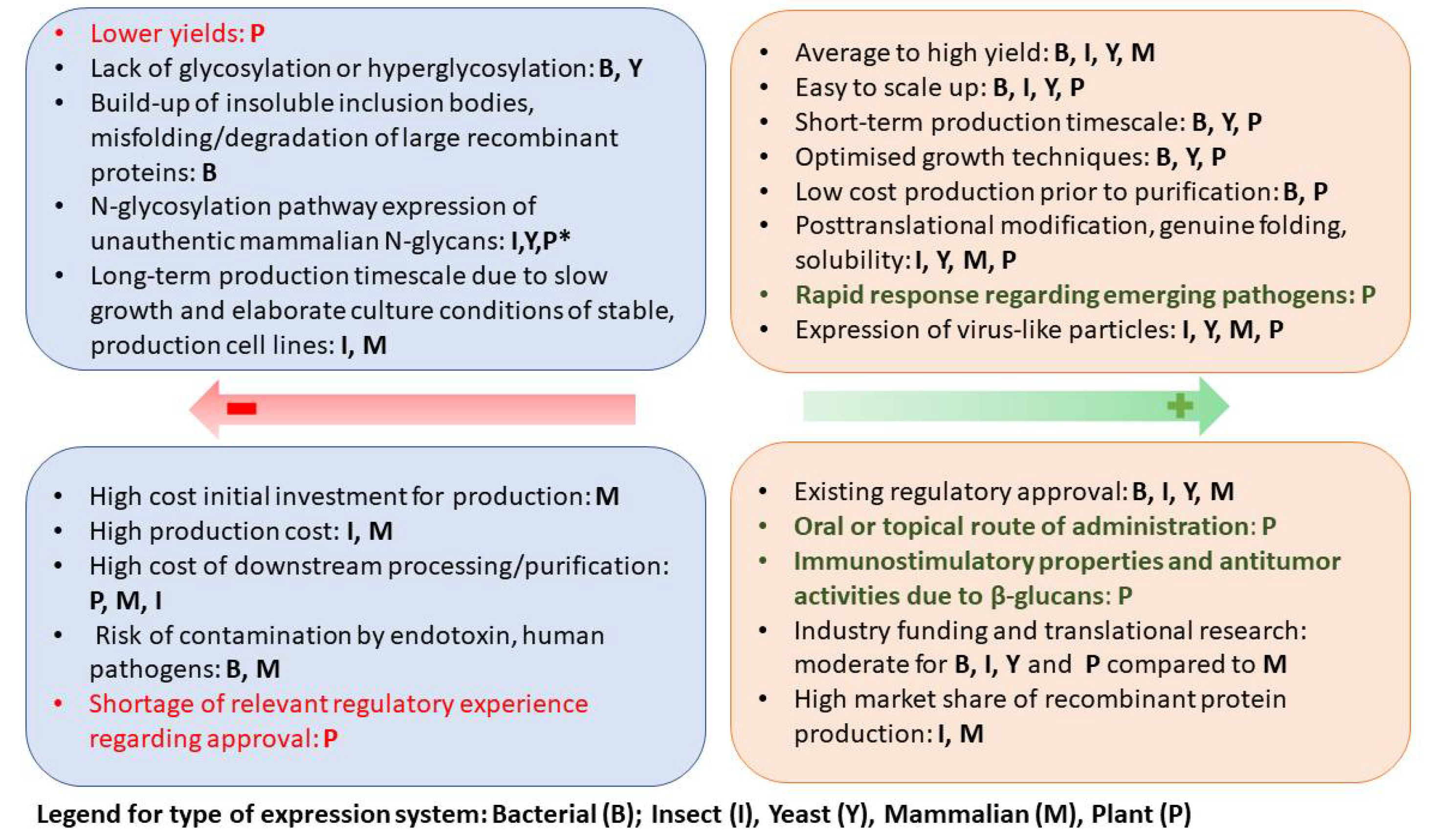{kind=link}
| Virus Family/Name | Main Animal Reservoir | Distribution | Cases/Mortality Rate |
|---|---|---|---|
| Family Orthomyxoviridae Influenza A virus | Wide variety of birds and mammals (dogs, cats, pigs, whales, horses) | Worldwide | 3–5 million severe cases annually, ~2% fatal rate |
| Family Coronaviridae Coronavirus SARS-CoV MERS CoV SARS-CoV-2 | Bats, pangolins, camels, monkeys, ferrets, minks, pigs, horses, dogs, cats, snakes | Worldwide | SARS-CoV~9.6%; MERS-CoV~34.3%; SARS-CoV-2 ranging from 2% to 7.6% depending on the country |
| Family Flaviviridae West Nile virus | Birds and horses | Africa, Europe, the Middle East, North America, and West Asia | ~1% of neuroinvasive disease and 5% of them are fatal |
| Zika virus | Human and non-human primates | Spread from Africa to the Pacific and the Americas | In Brazil, the estimated case fatality rate is 8.3% |
| Japanese encephalitis virus | Ardeid wading birds and pigs | Isolated in Japan. Spread to Southeast Asia, Australia | 35,000–50,000 cases of JE, with a mortality rate of 10,000–15,000 people per year |
| Dengue virus | Monkeys and dogs | Caribbean, Central Africa, the Eastern Mediterranean, Southeast Asia, and the Western Pacific | 390 million dengue virus infections per year, 1.3% fatality rate |
| Yellow fever virus | Humans and non-human primates | West, central and east Africa and in South America | 200,000 cases of YFVD and 30,000 deaths annually |
| Tick-borne encephalitis virus | Small rodents and large woodland animals | Europe, Siberia, northern China, Japan, and South Korea | 10,000–12,000 clinical cases of TBE and mortality rate depending on subtype (1% to 20%) |
| Family Filoviridae Ebola virus | Bats, Humans and non-human primates, wild antelopes | West and Central Africa | 2013–2016—28,616 cases. The fatality is ranging from 27% to 79% |
| Marburg virus | Fruit bats and monkeys | Central and South Africa | Fatality ratio of up to 88% |
| Family Nairoviridae Crime-Congo Hemorrhagic Fever Virus | Wild and domestic animals such as cattle, sheep, and goats | Africa, Asia, and Europe | Fatality ratio (10–40%) |
| Family Peribunyaviridae Rift Valley fever virus | Buffaloes, camels, cattle, goats, and sheep | Sub-Saharan Africa and Arabian Peninsula | Most human cases are mild. 50% mortality within hemorrhagic form |
| Family Togaviridae Chikungunya virus | Monkeys, birds, and rodents | Southeast Asia, Europe, Caribbean, North, Central, and South America | 1 million cases per year, the hospital mortality rate raging (10–26%) |
| Family Rhabdoviridae Rabies virus | Dogs and cats, foxes, wolves, mongooses, skunks, raccoons, bats | 95% of cases occurring in Africa and Asia | 59,000 human deaths annually |
| Family Retroviridae HIV | Originated in non-human primates | Worldwide | 37.7 million people are with HIV, the fatality rate is 1.8% |
| Family Paramyxoviridae Henipaviruses (Nipah virus and Hendra henipavirus) | Large fruit bats, pigs, horses | Southeast Asia, Australia, Papua New Guinea | The fatality rate is estimated at 40% to 75% |
| Newcastle disease virus | Poultry and wild birds | Mild systemic ND can be observed in humans | |
| Family Hepeviridae Hepatitis E virus | Pigs, wild boar, rats, rabbits, dears, dromedary camel, birds | Worldwide | 3.3 million symptomatic cases with a fatality rate up to 4% in the general population |
| Family Hantaviridae Hantavirus | Rodents, bats, and insectivores | Asia, Europe, and the Americas | Mortality rates of 12% |
| NCT Number | Study Title | Phase | Responsible Party |
|---|---|---|---|
| NCT04450004 | Safety, Tolerability and Immunogenicity of a Coronavirus-Like Particle COVID-19 Vaccine in Adults Aged 18–55 Years | 1 | Medicago |
| NCT04636697 | Study of a Recombinant Coronavirus-Like Particle COVID-19 Vaccine in Adults | 2/3 | Medicago |
| NCT05040789 | Phase-3 Study to Evaluate the Lot Consistency of a Recombinant Coronavirus-Like Particle COVID-19 Vaccine | 3 | Medicago |
| NCT05065619 | Safety Immunogenicity Study of MT-2766 in Japanese Adults (COVID-19) | 1/2 | Medicago |
| NCT04473690 | KBP-201 COVID-19 Vaccine Trial in Healthy Volunteers | 1/2 | KBP |
| NCT04953078 | A Study to Evaluate Safety, Tolerability, and Reactogenicity of an RBD-Fc-based Vaccine to Prevent COVID-19 | 1 | Baiya Phytopharm |
| Antigen | Production System and Expression Level | Immunogenicity | References |
|---|---|---|---|
| 22 aa epitope from gp41 | Cowpea; CPMV-HIV chimera; n/a. | Mice; Stimulated a strong serum neutralizing antibody response in three strains of mice. | [267,272,273] |
| CTB-P1 | N. benthamiana (transient); >80% of the CTB and CTB-P1 were assembled into functional oligomers. | Mice; Serum IgG response; mucosal IGA response; induction of immunological memory. | [274] |
| p24 | N. benthamiana (transient); 100 μg/g FW. | Rabbit; Induced specific humoral immune response. | [266] |
| NV and GAG | Tomato (transgenic); VLPs composed of the major antigenic protein for the hepatitis B virus (HBV); 0.3 ng/mg powder. | Mice; Induced a humoral immune response. | [275] |
| polHIV-1.op | Nicotiana tabacum (transgenic); VLPs composed of the major antigenic protein for the hepatitis B virus (HBV); 2–26 ng/g FW. | Mice; Elicited anti-HIV-1 specific CD8+ T cell activation detectable in mesenteric lymph nodes. | [268,269] |
| Tat | Tomato (transgenic); 1 μg/mg fruit (dry weight). | Mice; A strong anti-Tat immunological response after either intraperitoneal, intramuscular, or oral application. | [276] |
| p17/p24 | Nicotiana tabacum (transient); >1 mg p24/kg of FW. | Mice; Induced humoral and T cell immune response. | [277] |
| CTB-MPR | N. benthamiana (transgenic); 1–2 mg/kg FW. | Mice; Induction of serum and mucosal antibodies. | [278] |
| p24 | Nicotiana tabacum L. (transplastomic); n/a. | Mice; Serum IgG response. | [265] |
| C4(V3)6 | Lettuce (transgenic); 240 μg/g freeze-dried leaves. | Mice; Induction of humoral and cell-mediated immune response. | [263] |
| p24 | Arabidopsis thaliana (transgenic); 0.2 μg–0.5 μg/g FW. | Mice; Serum IgG response. | [264,279] |
| C4V3 (Multi-HIV) | Tobacco (transplastomic); 16 μg/g FW. | Mice; Induction of systemic mucosal, humoral, and T cell immune response. | [263,280,281] |
| Poly HIV | Physcomitrella patens (transgenic); 3.7 µg/g−1 FW. | Mice; Induction of specific antibody response. | [282] |
| Dgp41 and Gag | N. benthamiana (transient and transgenic respectively); ~9 mg/kg FW and ~22 mg/kg FW, respectively. | Mice; Serum antibodies against both the Gag and gp41 antigens were produced. CD4 and CD8 T cell response. | [283,284] |
| gp140 | N. benthamiana (transient); 21.5 mg/kg FW. | Rabbits; High titers of binding antibodies, including against the V1V2 loop region, and neutralizing antibodies against Tier 1 viruses. | [270,271] |
| Antigen | Production System and Expression Level | Immunogenicity | References |
|---|---|---|---|
| Fusion (F) and haemagglutinin-neuraminidase (HN) protein | Potato/stable transformation/0.3–0.6 µg/mg of total leaf protein | Oral and intraperitoneal delivery of the antigens elicited mucosal and systemic immune response | [319] |
| Fusion (F) | Rice/stable transformation/0.25–0.55 μg of purified NDV in 100 μg of total soluble leaf or seed proteins | Intraperitoneally immunized mice with crude protein extracts from transgenic rice plants elicited specific antibodies | [320] |
| Fusion (F) | Maize/stable transformation/ 0.9–3% TSP | Orally immunized chicken developed protective immune response | [321] |
| Fusion (F) and hemagglutinin-neuraminidase (HN) proteins | 0.5–0.8% of total seed protein Mize/stable transformation | Induced specific immune response after oral administration to chicken | [322] |
| Fusion (F) and hemagglutinin-neuraminidase (HN) proteins | Transgenic Canola Seeds | Chickens immunized orally with recombinant HN-F showed a significant rise in specific and hemagglutination inhibition (HI) antibodies | [323] |
| F and HN epitopes fused to Cucumber mosaic virus (CMV) | N. benthamiana/CMV formed VLPs, which served as carriers for display of neutralizing epitopes of F and HN/20–200 g of infected leaves | No data | [324,325] |
| Hemagglutinin-neuraminidase (HN) | N. benthamiana/transient expression | No data | [326] |
| Hemagglutinin-neuraminidase protein | Transgenic N. tobaccum 0.069% of TSP | Orally immunized chickens developed low titers of anti-HN serum IgG | [327] |
| Ectodomain of hemagglutinin-neuraminidase protein | Tobacco cell culture 0.2–0.4% of TSP | Mice receiving purified eHN protein from transgenic tobacco BY-2 cells produced specific anti-NDV antibodies | [19] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahmanova, G.; Takova, K.; Valkova, R.; Toneva, V.; Minkov, I.; Andonov, A.; Lukov, G.L. Plant-Derived Recombinant Vaccines against Zoonotic Viruses. Life 2022, 12, 156. https://doi.org/10.3390/life12020156
Zahmanova G, Takova K, Valkova R, Toneva V, Minkov I, Andonov A, Lukov GL. Plant-Derived Recombinant Vaccines against Zoonotic Viruses. Life. 2022; 12(2):156. https://doi.org/10.3390/life12020156
Chicago/Turabian StyleZahmanova, Gergana, Katerina Takova, Rumyana Valkova, Valentina Toneva, Ivan Minkov, Anton Andonov, and Georgi L. Lukov. 2022. "Plant-Derived Recombinant Vaccines against Zoonotic Viruses" Life 12, no. 2: 156. https://doi.org/10.3390/life12020156
APA StyleZahmanova, G., Takova, K., Valkova, R., Toneva, V., Minkov, I., Andonov, A., & Lukov, G. L. (2022). Plant-Derived Recombinant Vaccines against Zoonotic Viruses. Life, 12(2), 156. https://doi.org/10.3390/life12020156