
Silver Nanoparticle Production by the Cyanobacterium Cyanothece sp.: De Novo Manipulation of Nano-Biosynthesis by Phytohormones



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of the Cyanobacterium Cyanothece sp.
2.2. Production of Silver Nanoparticles in Cyanothece sp. Cultures
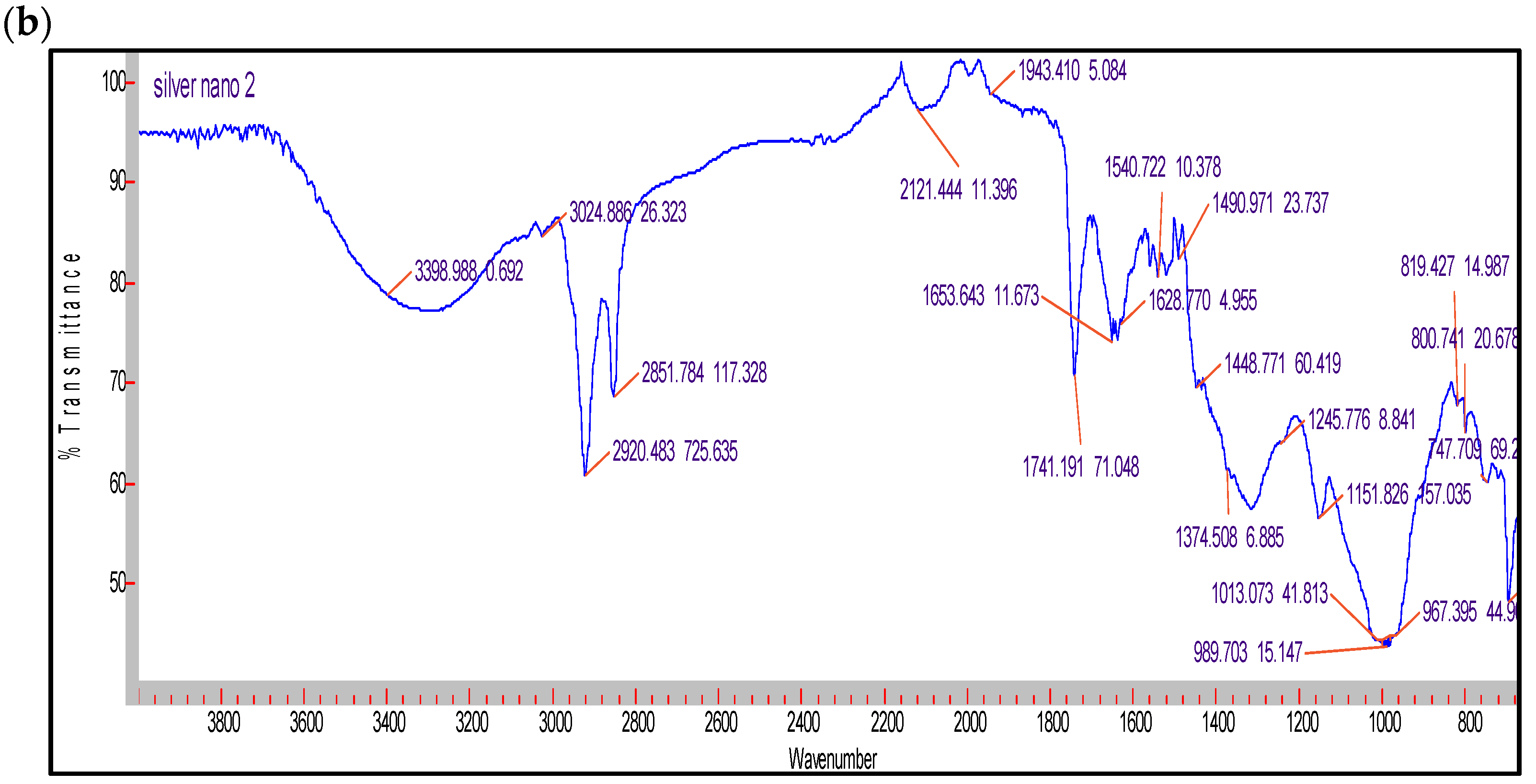
2.3. Characterization of the AgNPs Produced by the Cyanobacterium Cyanothece sp. Cultures
2.4. Determination of Percentage Conversion of Ag+ to Ag0
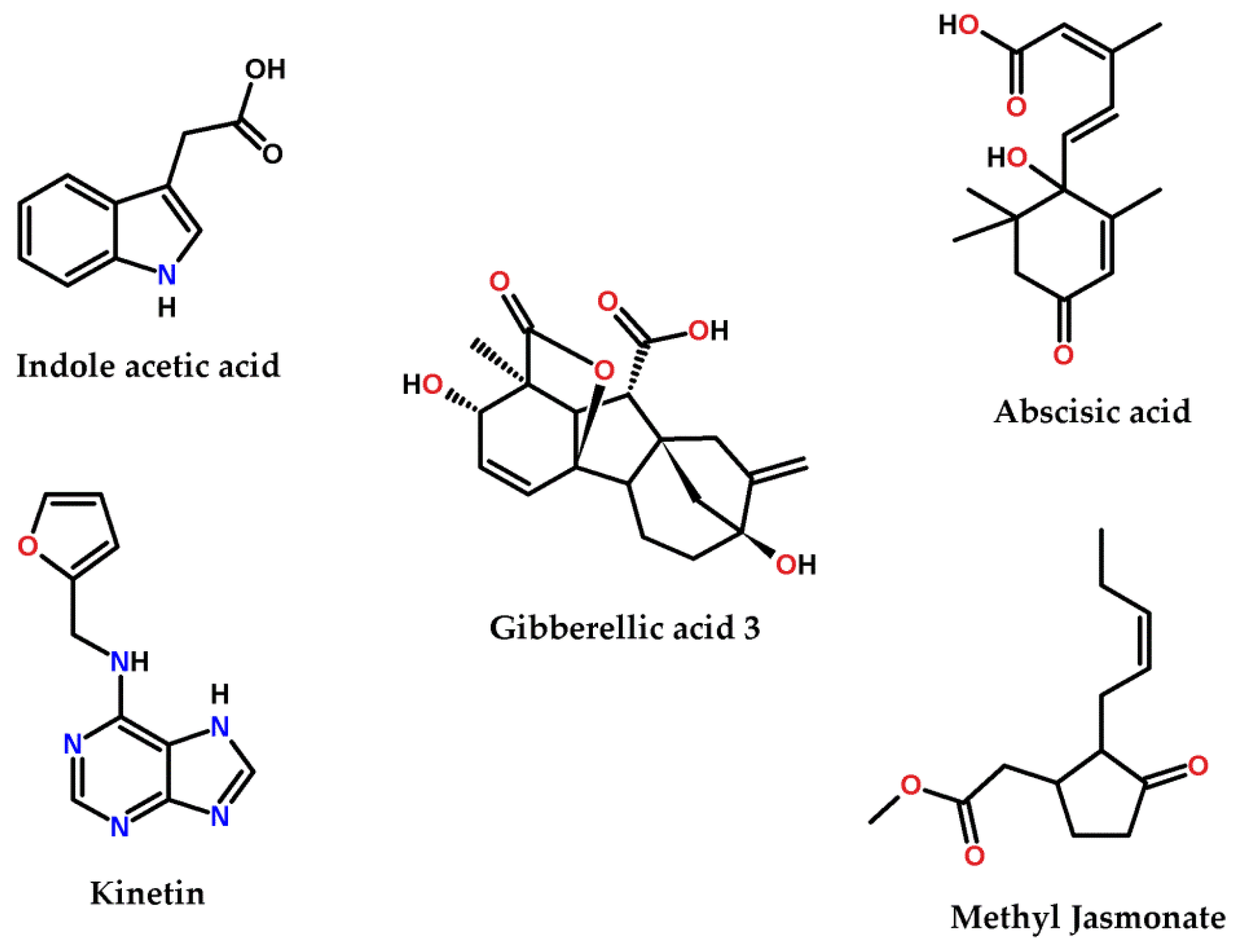
2.5. Examination of Different Phytohormones’ Effect on AgNP Production
2.6. Statistical Analysis
3. Results
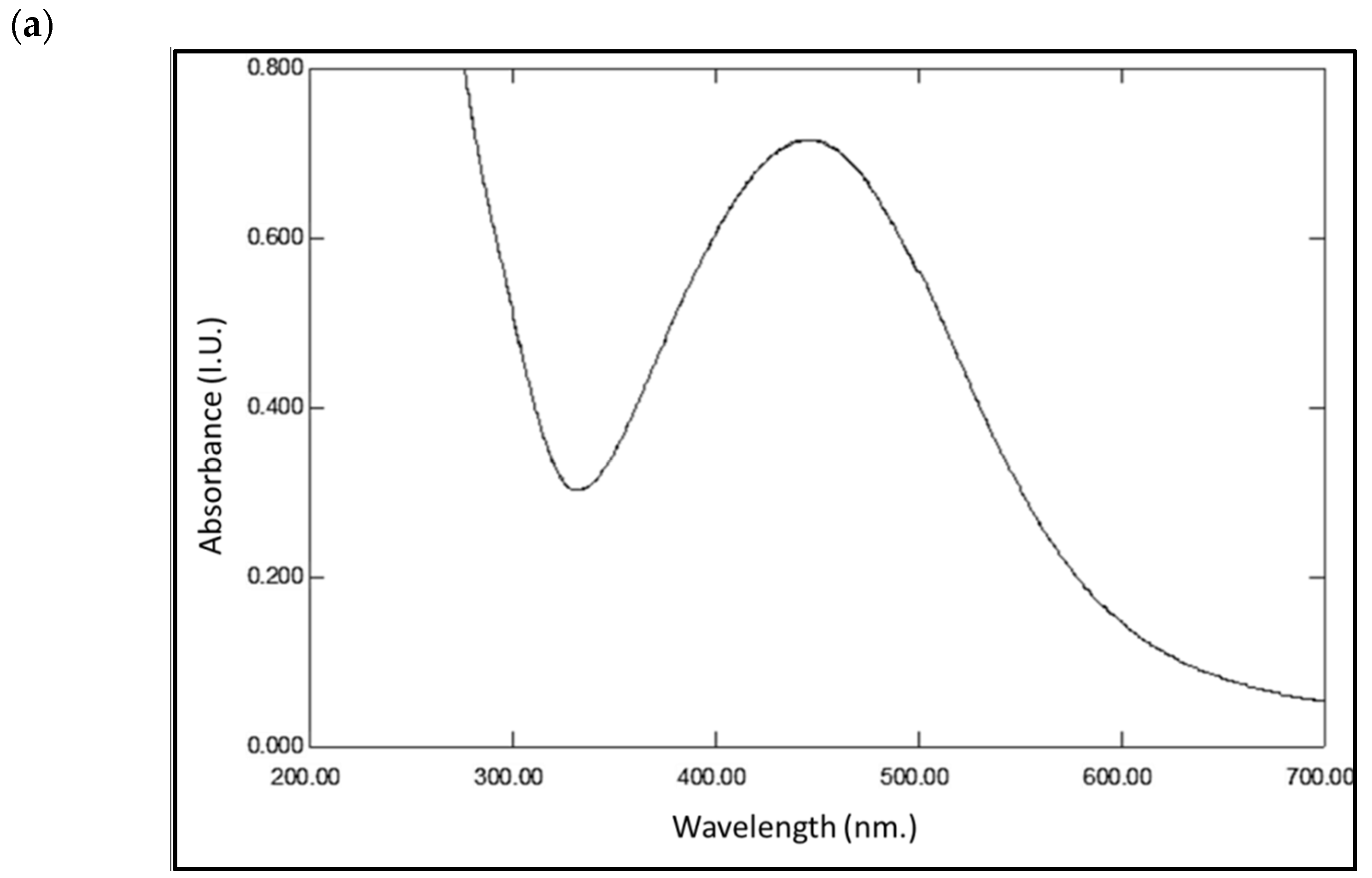
3.1. Production of AgNPs by Cyanothece sp. Cultures
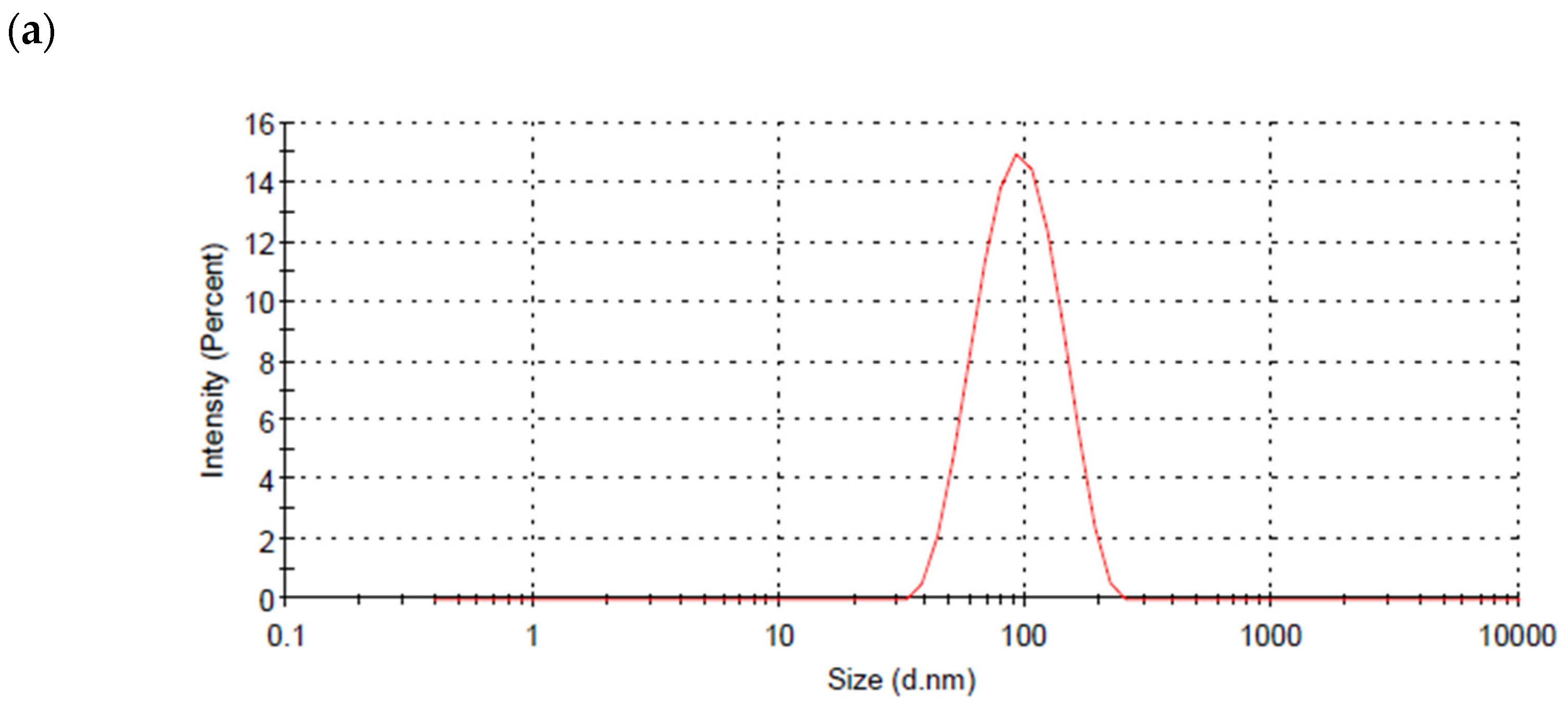
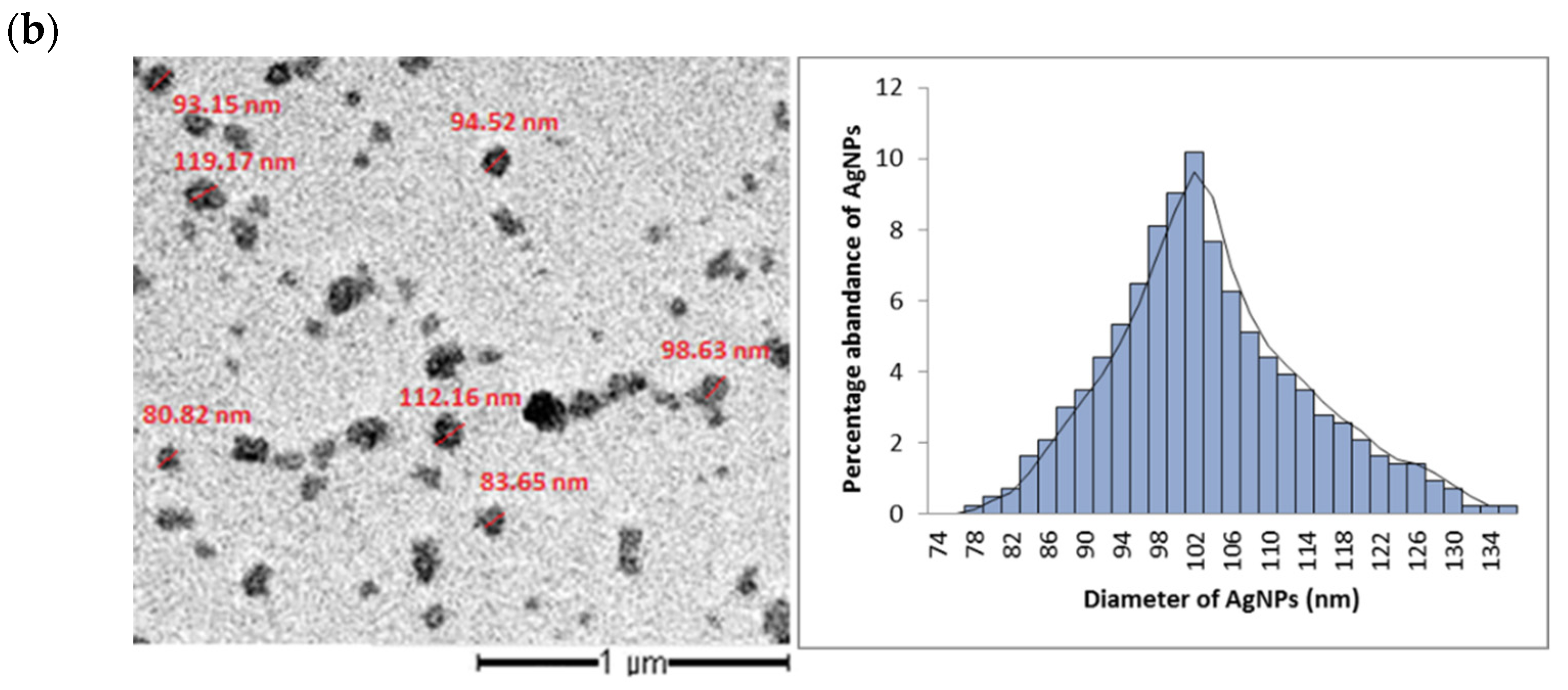
3.2. Particle Size and Shape of AgNPs Produced by the Cyanobacterium Cyanothece sp. Cultures
3.3. Characterization of the AgNPs Produced by the Cyanobacterium Cyanothece sp. Cultures
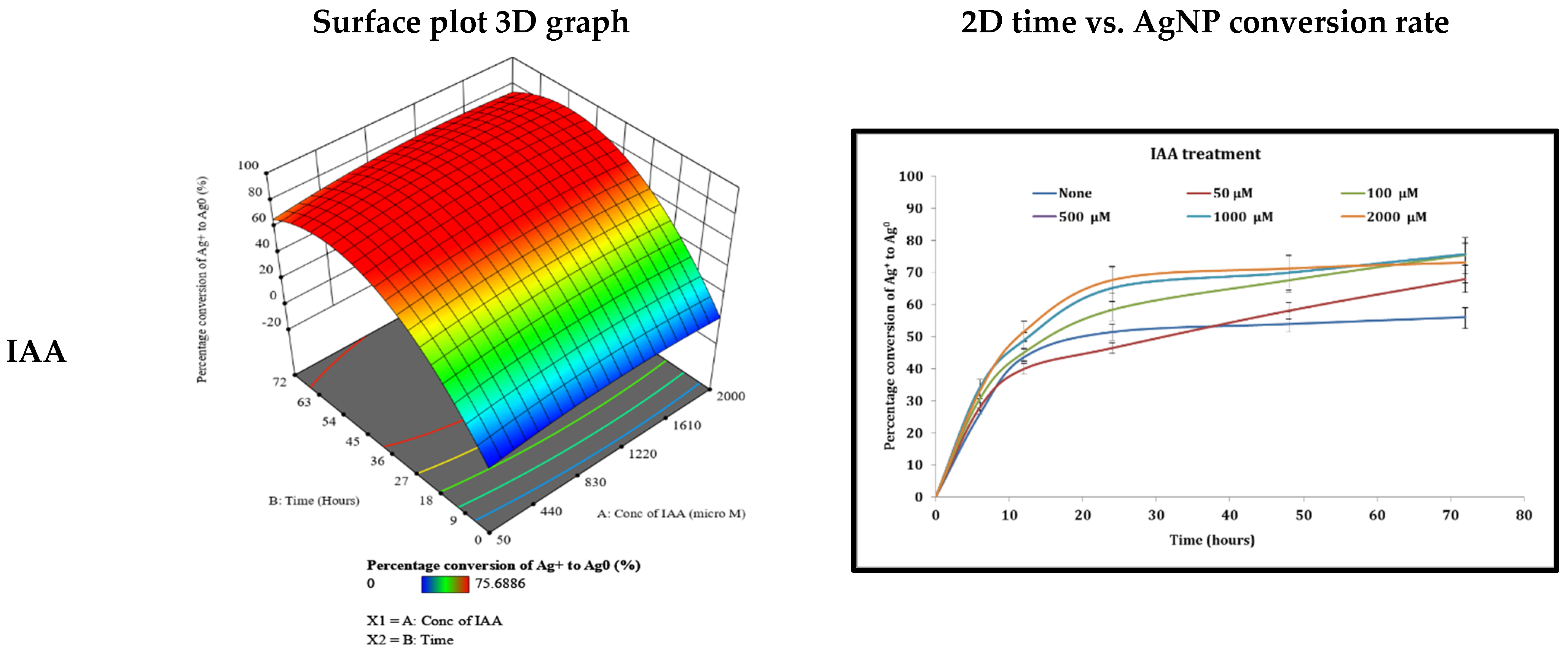
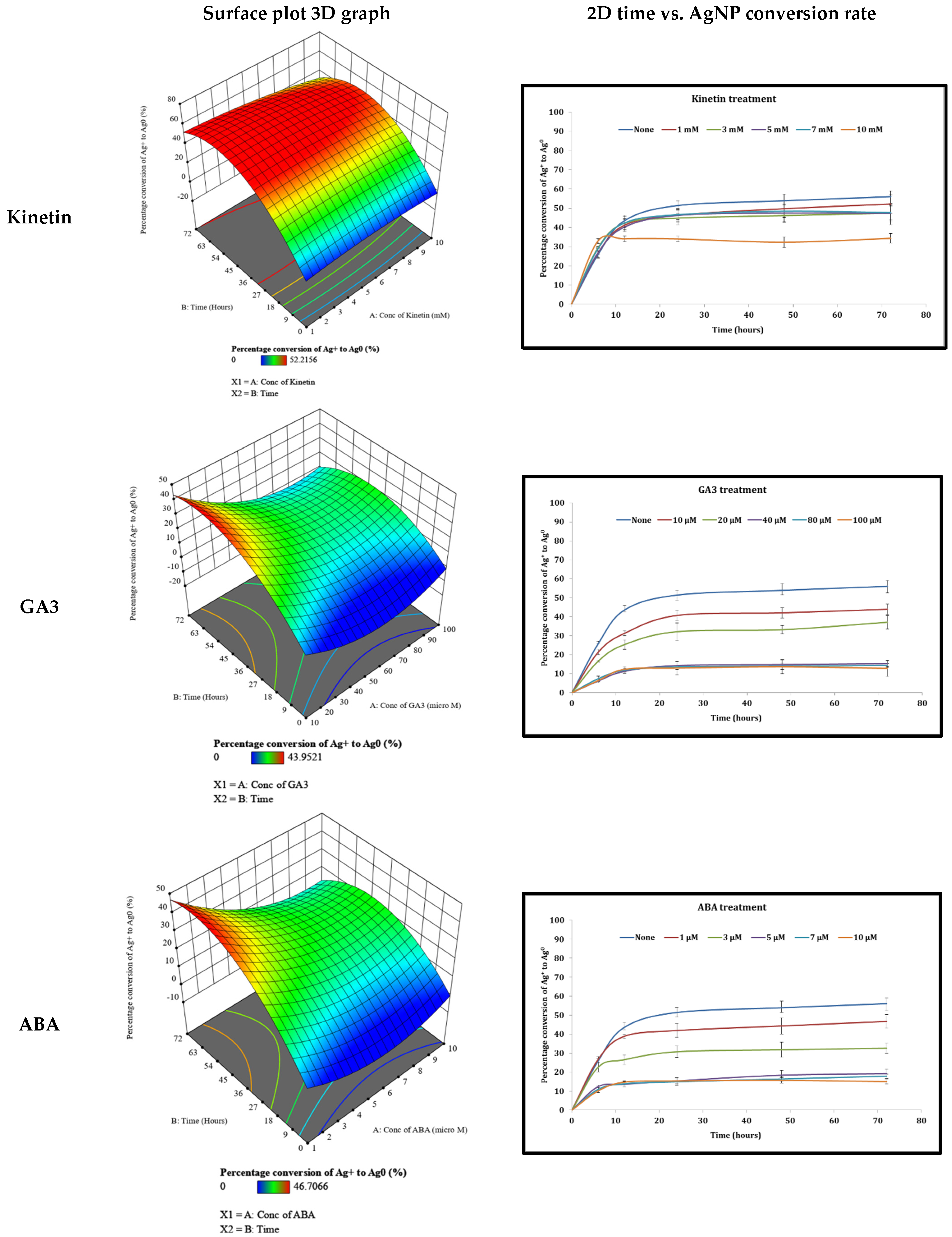
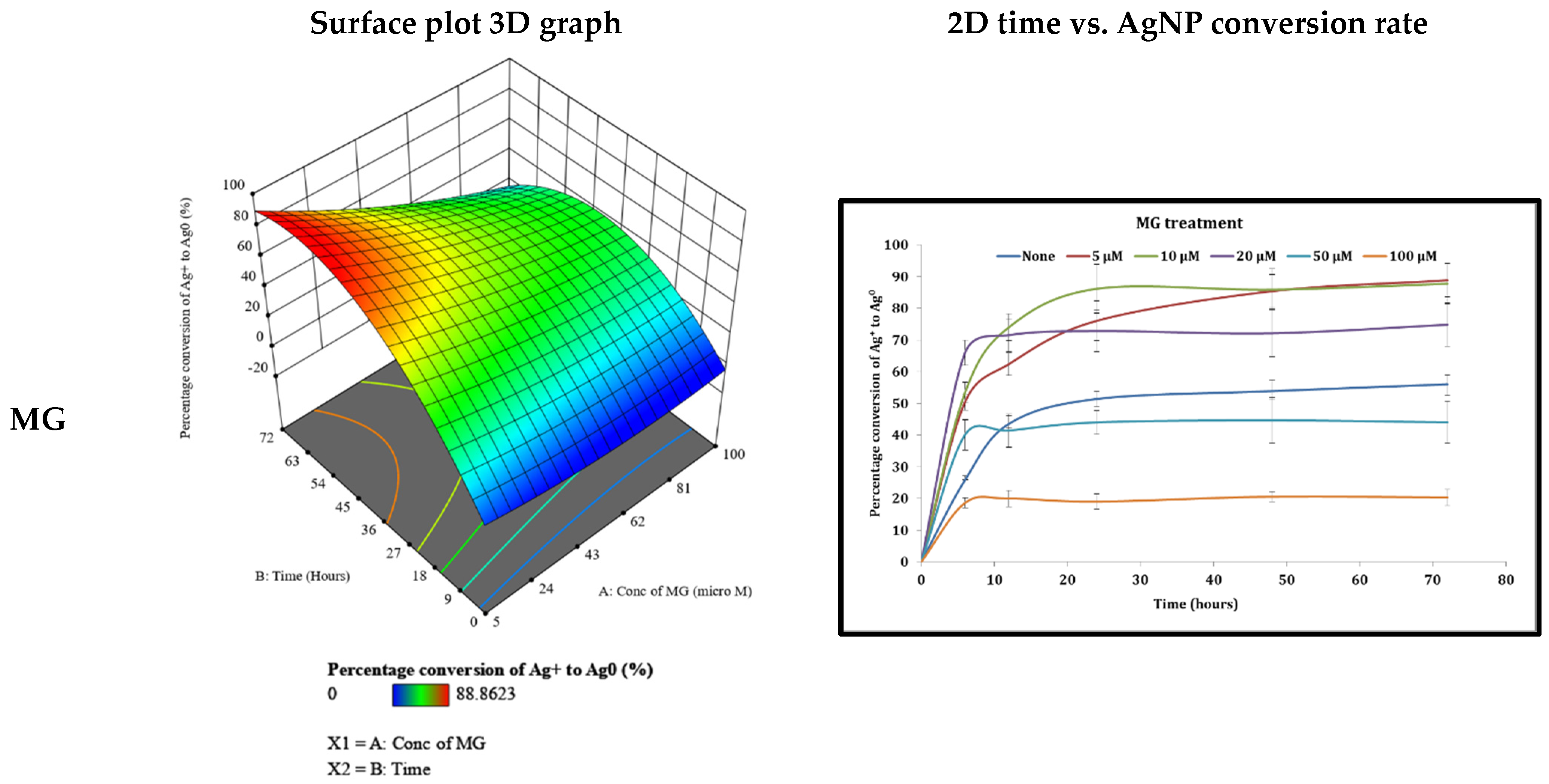
3.4. Examination of Different Phytohormones’ Effects on AgNP Production
4. Discussion
4.1. Prodcution of AgNPs from Cyanothece sp. Cultures
4.2. The Effect of Different Phytohormones on AgNPs’ Production from Cyanothece sp. Cultures
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bandyopadhyay, A.; Elvitigala, T.; Welsh, E.; Stöckel, J.; Liberton, M.; Min, H.; Sherman, L.A.; Pakrasi, H.B. Novel Metabolic Attributes of the Genus Cyanothece, Comprising a Group of Unicellular Nitrogen-Fixing Cyanobacteria. mBio 2011, 2, e00214-11. [Google Scholar] [CrossRef] [Green Version]
- Oftedal, L.; Skjærven, K.H.; Coyne, R.T.; Edvardsen, B.; Rohrlack, T.; Skulberg, O.M.; Døskeland, S.O.; Herfindal, L. The apoptosis-inducing activity towards leukemia and lymphoma cells in a cyanobacterial culture collection is not associated with mouse bioassay toxicity. J. Ind. Microbiol. Biotechnol. 2010, 38, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Ascencio, F.; Gama, N.L.; De Philippis, R.; Ho, B. Effectiveness of Cyanothece spp. andCyanospira capsulata exocellular polysaccharides as antiadhesive agents for blocking attachment of Helicobacter pylori to human gastric cells. Folia Microbiol. 2004, 49, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Bakir, E.M.; Younis, N.S.; Mohamed, M.E.; El Semary, N.A. Cyanobacteria as Nanogold Factories: Chemical and Anti-Myocardial Infarction Properties of Gold Nanoparticles Synthesized by Lyngbya majuscula. Mar. Drugs 2018, 16, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younis, N.S.; Bakir, E.M.; Mohamed, M.E.; El Semary, N.A. Cyanobacteria as Nanogold Factories II: Chemical Reactivity and anti-Myocardial Infraction Properties of Customized Gold Nanoparticles Biosynthesized by Cyanothece sp. Mar. Drugs 2019, 17, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.N.; Verma, I.; Kumar, M. Chapter 23—Cyanobacteria: Potential source of biofertilizer and synthesizer of metallic nanoparticles. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Lengke, M.F.; Fleet, A.M.E.; Southam, G. Morphology of Gold Nanoparticles Synthesized by Filamentous Cyanobacteria from Gold(I)−Thiosulfate and Gold(III)−Chloride Complexes. Langmuir 2006, 22, 2780–2787. [Google Scholar] [CrossRef] [PubMed]
- Kalabegishvili, T.; Kirkesali, E.I.; Rcheulishvili, A.N.; Ginturi, E.; Murusidze, I.; Kuchava, N.; Bagdavadze, N.; Tsertsvadze, G.; Gabunia, V.; Frontasyeva, M.; et al. Synthesis of gold nanoparticles by blue-green algae Spirulina platensis. Adv. Sci. Eng. Med. 2013, 4. [Google Scholar] [CrossRef]
- Kumar, B.; Smita, K.; Sánchez, E.; Guerra, S.; Cumbal, L. Ecofriendly ultrasound-assisted rapid synthesis of gold nanoparticles using Calothrix algae. Adv. Nat. Sci. Nanosci. Nanotechnol. 2016, 7, 025013. [Google Scholar] [CrossRef]
- Burdușel, A.-C.; Gherasim, O.; Grumezescu, A.M.; Mogoantă, L.; Ficai, A.; Andronescu, E. Biomedical Applications of Silver Nanoparticles: An Up-to-Date Overview. Nanomaterials 2018, 8, 681. [Google Scholar] [CrossRef] [Green Version]
- Patel, V.; Berthold, D.; Puranik, P.; Gantar, M. Screening of cyanobacteria and microalgae for their ability to synthesize silver nanoparticles with antibacterial activity. Biotechnol. Rep. 2014, 5, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. Auxin Biosynthesis and Its Role in Plant Development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamayun, M.; Hussain, A.; Khan, S.A.; Irshad, M.; Khan, A.L.; Waqas, M.; Shahzad, R.; Iqbal, A.; Ullah, N.; Rehman, G.; et al. Kinetin modulates physio-hormonal attributes and isoflavone contents of Soybean grown under salinity stress. Front. Plant Sci. 2015, 6, 377. [Google Scholar] [CrossRef] [Green Version]
- Erin, N.; Afacan, B.; Ersoy, Y.; Ercan, F.; Balcı, M.K. Gibberellic acid, a plant growth regulator, increases mast cell recruitment and alters Substance P levels. Toxicology 2008, 254, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Wareing, P.F.; Bradbeer, J.W. Abscisic acid as a natural growth regulator. Philos. Trans. R. Soc. B Biol. Sci. 1978, 284, 483–498. [Google Scholar] [CrossRef]
- Cheong, J.-J.; Do Choi, Y. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Karthikeyan, N.; Prasanna, R.; Sood, A.; Jaiswal, P.; Nayak, S.; Kaushik, B.D. Physiological characterization and electron microscopic investigation of cyanobacteria associated with wheat rhizosphere. Folia Microbiol. 2009, 54, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S.; Pandey, V.D. Alkaline phosphatase activity in freshwater cyanobacteria. Plant Arch. 2011, 11, 827–830. [Google Scholar]
- Lu, Y.; Xu, J. Phytohormones in microalgae: A new opportunity for microalgal biotechnology? Trends Plant Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. Impacts of Silver Nanoparticles on Plants: A Focus on the Phytotoxicity and Underlying Mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Pallavi, N.; Mehta, C.M.; Srivastava, R.; Arora, S.; Sharma, A.K. Impact assessment of silver nanoparticles on plant growth and soil bacterial diversity. 3 Biotech 2016, 6, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmast, M.K.; Salehi, H. Silver Nanoparticles: An Influential Element in Plant Nanobiotechnology. Mol. Biotechnol. 2016, 58, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana hustedt, and detonula Confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, R.A.; Hussein, M.H.; Abo-Elmagd, R.A.; Bawazir, S.S. Synthesis and biological characterization of silver nanoparticles derived from the cyanobacterium Oscillatoria limnetica. Sci. Rep. 2019, 9, 13071. [Google Scholar] [CrossRef]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.A.; Jeffryes, C. Biosynthetic Conversion of Ag+ to highly Stable Ag0 Nanoparticles by Wild Type and Cell Wall Deficient Strains of Chlamydomonas reinhardtii. Molecules 2018, 24, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maarebia, R.Z.; Wahab, A.W.; Taba, P. Synthesis and Characterization Of Silver Nanoparticles Using Water Extract of Sarang Semut (Myrmecodia pendans) For Blood Glucose Sensors. J. Akta Kim. Indones. (Indonesia Chim. Acta) 2019, 12, 29–46. [Google Scholar] [CrossRef]
- Biosynthesis and Characterization of Silver Nanoparticles from Marine Macroscopic Brown Seaweed Colpomenia sinuosa (Mertens ex Roth) Derbes and Solier. J. Adv. Chem. Sci. 2020, 6, 663–666. [CrossRef]
- Borah, D.; Nainamalai, S.; Gopalakrishnan, S.; Rout, J.; Alharbi, N.S.; Alharbi, S.A.; Nooruddin, T. Biolubricant potential of exopolysaccharides from the cyanobacterium Cyanothece epiphytica. Appl. Microbiol. Biotechnol. 2018, 102, 3635–3647. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Su, C.; Lu, W. A new exopolysaccharide produced by marine Cyanothece sp. 113. Bioresour. Technol. 2007, 98, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Welkie, D.G.; Sherman, D.M.; Chrisler, W.B.; Orr, G.; Sherman, L.A. Analysis of carbohydrate storage granules in the diazotrophic cyanobacterium Cyanothece sp. PCC 7822. Photosynth. Res. 2013, 118, 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agami, R.A. Pre-soaking in indole-3-acetic acid or spermidine enhances copper tolerance in wheat seedlings. S. Afr. J. Bot. 2016, 104, 167–174. [Google Scholar] [CrossRef]
- Gangwar, S.; Singh, V.P.; Prasad, S.M.; Maurya, J.N. Differential responses of pea seedlings to indole acetic acid under manganese toxicity. Acta Physiol. Plant. 2010, 33, 451–462. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Ilias, I. Hormone-induced protection of sunflower photosynthetic apparatus against copper toxicity. Biol. Plant. 2005, 49, 223–228. [Google Scholar] [CrossRef]
- Gong, Q.; Li, Z.; Wang, L.; Dai, T.; Kang, Q.; Niu, D. Exogenous of Indole-3-Acetic Acid Application Alleviates Copper Toxicity in Spinach Seedlings by Enhancing Antioxidant Systems and Nitrogen Metabolism. Toxics 2019, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Kotowska, U.; Zambrzycka-Szelewa, E.; Sienkiewicz, A. Auxins and Cytokinins Regulate Phytohormone Homeostasis and Thiol-Mediated Detoxification in the Green Alga Acutodesmus obliquus Exposed to Lead Stress. Sci. Rep. 2020, 10, 10193. [Google Scholar] [CrossRef]
- Khalil, R.; Haroun, S.; Bassyoini, F.; Nagah, A.; Yusuf, M. Salicylic acid in combination with kinetin or calcium ameliorates heavy metal stress in Phaseolus vulgaris plant. J. Agric. Food Res. 2021, 5, 100182. [Google Scholar] [CrossRef]
- Zhou, M.; Ghnaya, T.; Dailly, H.; Cui, G.; Vanpee, B.; Han, R.; Lutts, S. The cytokinin trans-zeatine riboside increased resistance to heavy metals in the halophyte plant species Kosteletzkya pentacarpos in the absence but not in the presence of NaCl. Chemosphere 2019, 233, 954–965. [Google Scholar] [CrossRef] [PubMed]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Shin, J.-H.; Ahmad, B.; Shin, D.-H.; Lee, I.-J. Exogenous Gibberellic Acid Reprograms Soybean to Higher Growth and Salt Stress Tolerance. J. Agric. Food Chem. 2010, 58, 7226–7232. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Interactive effect of calcium and gibberellin on nickel tolerance in relation to antioxidant systems in Triticum aestivum L. Protoplasma 2010, 248, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Gangwar, S.; Singh, V.P.; Srivastava, P.K.; Maurya, J.N. Modification of chromium (VI) phytotoxicity by exogenous gibberellic acid application in Pisum sativum (L.) seedlings. Acta Physiol. Plant. 2010, 33, 1385–1397. [Google Scholar] [CrossRef]
- El-Monem, A.; Sharaf, M.; Farghal, I.; Sofy, M. Role of Gibberellic Acid in Abolishing the Detrimental Effects of Cd and Pb on Broad Bean and Lupin Plants. Res. J. Agric. Biol. Sci. 2009, 668–673. [Google Scholar]
- Khedr, M.A.; Massarotti, A.; Mohamed, M. Rational Discovery of (+) (S) Abscisic Acid as a Potential Antifungal Agent: A Repurposing Approach. Sci. Rep. 2018, 8, 8565. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Y.; Wang, J.W.; Babla, M.; Zhao, C.; García-Mata, C.; Sani, E.; Differ, C.; Mak, M.; Hills, A.; et al. Nitrate reductase mutation alters potassium nutrition as well as nitric oxide-mediated control of guard cell ion channels in Arabidopsis. New Phytol. 2015, 209, 1456–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabala, S.; Chen, G.; Chen, Z.; Pottosin, I. The energy cost of the tonoplast futile sodium leak. New Phytol. 2019, 225, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Amir, R.; Taufiq, S.; Noor, N.; Nauman, I.; Munir, F.; Keyani, R.; Tahir, A.T. Stress Signaling Under Metal and Metalloid Toxicity. In Plants Under Metal and Metalloid Stress; Springer: Singapore, 2018; pp. 149–184. [Google Scholar] [CrossRef]
- Pan, W.; You, Y.; Shentu, J.-L.; Weng, Y.-N.; Wang, S.-T.; Xu, Q.-R.; Liu, H.-J.; Du, S.-T. Abscisic acid (ABA)-importing transporter 1 (AIT1) contributes to the inhibition of Cd accumulation via exogenous ABA application in Arabidopsis. J. Hazard. Mater. 2020, 391, 122189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xing, D. Methyl Jasmonate Induces Production of Reactive Oxygen Species and Alterations in Mitochondrial Dynamics that Precede Photosynthetic Dysfunction and Subsequent Cell Death. Plant Cell Physiol. 2008, 49, 1092–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Zhang, W.; Chen, J.; Li, X. Methyl jasmonate alleviates cadmium toxicity in Solanum nigrum by regulating metal uptake and antioxidative capacity. Biol. Plant. 2015, 59, 373–381. [Google Scholar] [CrossRef]
- Keramat, B.; Kalantari, K.; Arvin, M. Effects of methyl jasmonate in regulating cadmium induced oxidative stress in soybean plant (Glycine max L.). Afr. J. Microbiol. Res. 2009, 3, 240–244. [Google Scholar]
- Alikhani, O.; Abbaspour, H. Effects of methyl jasmonate and cadmium on growth traits, cadmium transport and accumulation, and allene-oxide cyclase gene expression in wheat seedlings. Rev. DE Agric. Neotropical 2019, 6, 20–29. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytohormone * | Ideal Predicted Factors for Maximum Response | Maximum Response | |
|---|---|---|---|
| Concentration | Time (Hours) | Conversion of Ag+ to Ag0 (%) | |
| IAA | 1172.26 (μM) | 45.24 | 87.29 |
| Kinetin | 7.514 (mM) | 40.90 | 55.16 |
| GA3 | 10.56 (μM) | 61.42 | 45.23 |
| ABA | 1.065 (μM) | 52.81 | 47.95 |
| MG | 8.581 (μM) | 54.12 | 90.29 |
| Phytohormone * | Concentration | Behavior of Cyanobacteria over Time | |||||
|---|---|---|---|---|---|---|---|
| 0 (h) | 6 (h) | 12 (h) | 24 (h) | 48 (h) | 72 (h) | ||
| None | None | ||||||
| IAA | 50 (μM) | ||||||
| 100 (μM) | |||||||
| 500 (μM) | |||||||
| 1000 (μM) | |||||||
| 2000 (μM) | |||||||
| Kinetin | 1 (mM) | ||||||
| 3 (mM) | |||||||
| 5 (mM) | |||||||
| 7 (mM) | |||||||
| 10 (mM) | |||||||
| GA3 | 10 (μM) | ||||||
| 20 (μM) | |||||||
| 40 (μM) | |||||||
| 80 (μM) | |||||||
| 100 (μM) | |||||||
| ABA | 1 (μM) | ||||||
| 3 (μM) | |||||||
| 5 (μM) | |||||||
| 7 (μM) | |||||||
| 10 (μM) | |||||||
| MG | 5 (μM) | ||||||
| 10 (μM) | |||||||
| 20 (μM) | |||||||
| 50 (μM) | |||||||
| 100 (μM) | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, M.E.; El Semary, N.A.; Younis, N.S. Silver Nanoparticle Production by the Cyanobacterium Cyanothece sp.: De Novo Manipulation of Nano-Biosynthesis by Phytohormones. Life 2022, 12, 139. https://doi.org/10.3390/life12020139
Mohamed ME, El Semary NA, Younis NS. Silver Nanoparticle Production by the Cyanobacterium Cyanothece sp.: De Novo Manipulation of Nano-Biosynthesis by Phytohormones. Life. 2022; 12(2):139. https://doi.org/10.3390/life12020139
Chicago/Turabian StyleMohamed, Maged E., Nermin A. El Semary, and Nancy S. Younis. 2022. "Silver Nanoparticle Production by the Cyanobacterium Cyanothece sp.: De Novo Manipulation of Nano-Biosynthesis by Phytohormones" Life 12, no. 2: 139. https://doi.org/10.3390/life12020139
APA StyleMohamed, M. E., El Semary, N. A., & Younis, N. S. (2022). Silver Nanoparticle Production by the Cyanobacterium Cyanothece sp.: De Novo Manipulation of Nano-Biosynthesis by Phytohormones. Life, 12(2), 139. https://doi.org/10.3390/life12020139