
Blue Laser Light Counteracts HSV-1 in the SH-SY5Y Neuronal Cell Model of Infection



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. HSV-1 Culture Infection
2.1.1. Viability Assay
2.1.2. Viral DNA Quantification
2.2. Laser Irradiation
2.3. Antiviral Effect of PBMT
- Irradiation of HSV-1 (5 × 104 viral copies)—the virus was treated and after 30 min transferred to the cells and maintained in incubator for 24 h.
- Irradiation of HSV-1 infected culture (5 × 104 viral copies)—the cells were infected for 1 h and then irradiated.
2.4. Statistical Analysis
3. Results
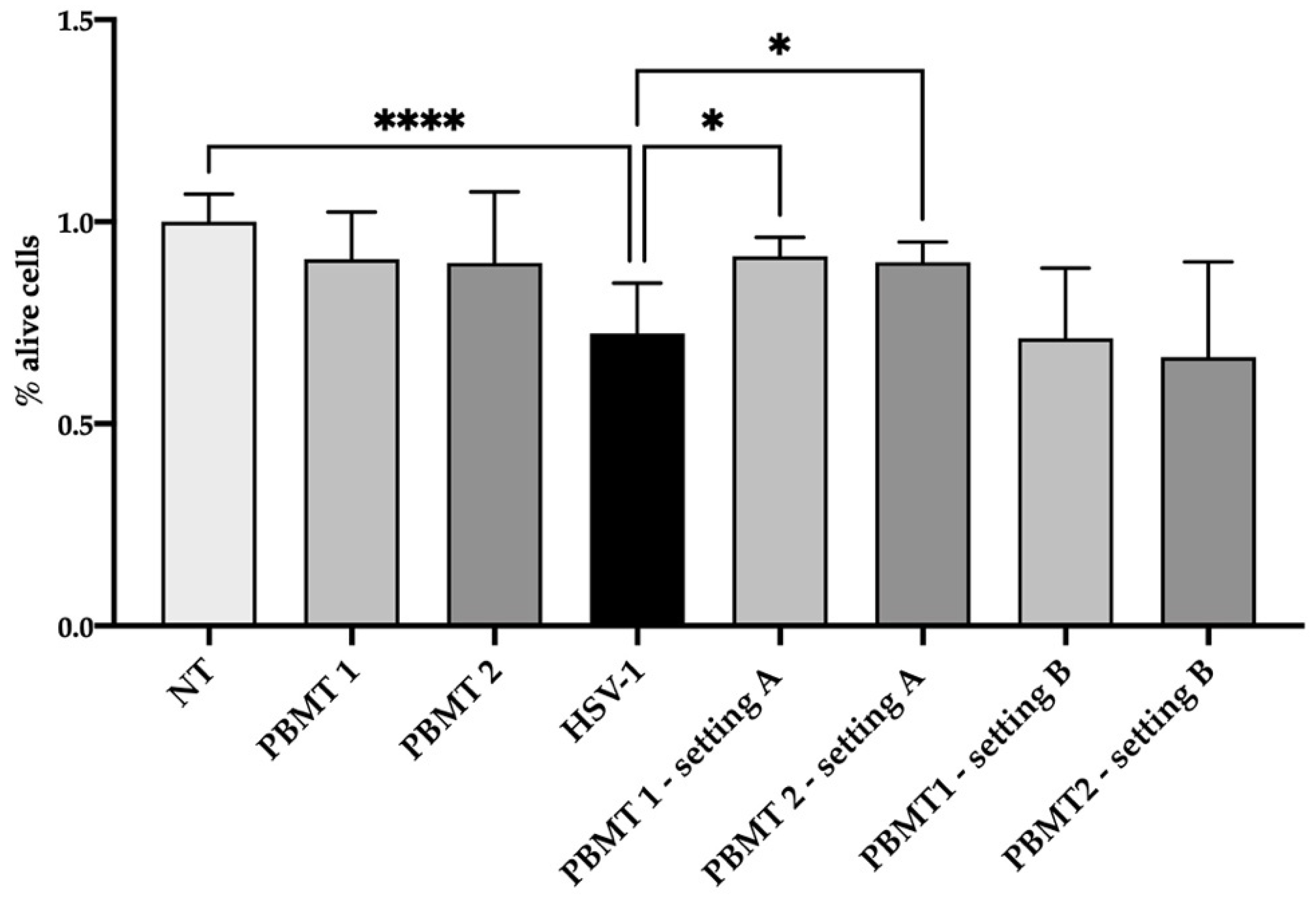
3.1. HSV-1 Cell Culture Infection
3.2. PBMT Protocols
3.3. Irradiation of HSV-1
3.4. HSV-1 Cell Culture Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Corey, L. Herpes Simplex Virus Infections. In Harrison’s Principles of Internal Medicine; Longo, D.L., Fauci, A.S., Kasper, D.L., Hauser, S.L., Jameson, J.L., Loscalzo, J., Eds.; The McGraw-Hill Companies: New York, NY, USA, 2012; Chapter 179. [Google Scholar]
- World Health Organization. Herpes Simplex Virus. Available online: http://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 15 November 2021).
- Yousuf, W.; Ibrahim, H.; Harfouche, M.; Abu Hijleh, F.; Abu-Raddad, L. Herpes Simplex Virus Type 1 in Europe: Systematic Review, Meta-Analyses and Meta-Regressions. BMJ Glob. Health 2020, 5, e002388. [Google Scholar] [CrossRef] [PubMed]
- Opstelten, W.; Neven, A.K.; Eekhof, J. Treatment and Prevention of Herpes Labialis. Can. Fam. Physician 2008, 54, 1683–1687. [Google Scholar]
- De Freitas, L.F.; Hamblin, M.R. Proposed Mechanisms of Photobiomodulation or Low-Level Light Therapy. IEEE J. Sel. Top. Quantum Electron. 2016, 22, 348–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paula Eduardo, C.; Aranha, A.C.C.; Simões, A.; Bello-Silva, M.S.; Ramalho, K.M.; Esteves-Oliveira, M.; de Freitas, P.M.; Marotti, J.; Tunér, J. Laser Treatment of Recurrent Herpes Labialis: A Literature Review. Lasers Med. Sci. 2013, 29, 1517–1529. [Google Scholar] [CrossRef] [PubMed]
- De Paula Eduardo, C.; Bezinelli, L.M.; de Paula Eduardo, F.; da Graça Lopes, R.M.; Ramalho, K.M.; Bello-Silva, M.S.; Esteves-Oliveira, M. Prevention of Recurrent Herpes Labialis Outbreaks through Low-Intensity Laser Therapy: A Clinical Protocol with 3-Year Follow-Up. Lasers Med. Sci. 2012, 27, 1077–1083. [Google Scholar] [CrossRef]
- Wainwright, M. Local Treatment of Viral Disease Using Photodynamic Therapy. Int. J. Antimicrob. Agents 2003, 21, 510–520. [Google Scholar] [CrossRef]
- Donnarumma, G.; De Gregorio, V.; Fusco, A.; Farina, E.; Baroni, A.; Esposito, V.; Contaldo, M.; Petruzzi, M.; Pannone, G.; Serpico, R. Inhibition of HSV-1 Replication by Laser Diode-Irradiation: Possible Mechanism of Action. Int. J. Immunopathol. Pharmacol. 2010, 23, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Dai, T. The Antimicrobial Effect of Blue Light: What Are Behind? Virulence 2017, 8, 649–652. [Google Scholar] [CrossRef] [Green Version]
- Rupel, K.; Zupin, L.; Ottaviani, G.; Bertani, I.; Martinelli, V.; Porrelli, D.; Vodret, S.; Vuerich, R.; Passos da Silva, D.; Bussani, R.; et al. Blue Laser Light Inhibits Biofilm Formation in Vitro and in Vivo by Inducing Oxidative Stress. NPJ Biofilms Microbiomes 2019, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zupin, L.; Gratton, R.; Fontana, F.; Clemente, L.; Pascolo, L.; Ruscio, M.; Crovella, S. Blue Photobiomodulation LED Therapy Impacts SARS-CoV-2 by Limiting Its Replication in Vero Cells. J. Biophotonics 2021, 14, e202000496. [Google Scholar] [CrossRef]
- Zupin, L.; Caracciolo, I.; Tricarico, P.M.; Ottaviani, G.; D’Agaro, P.; Crovella, S. Antiviral Properties of Blue Laser in an in Vitro Model of HSV-1 Infection. Microbiol. Immunol. 2018, 62, 477–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zupin, L.; Caracciolo, I.; Tricarico, P.M.; Ottaviani, G.; D’Agaro, P.; Crovella, S. Photobiomodulation Therapy Reduces Viral Load and Cell Death in ZIKV-Infected Glioblastoma Cell Line. Lasers Med. Sci. 2018, 33, 2011–2013. [Google Scholar] [CrossRef]
- Bunz, O.; Mese, K.; Funk, C.; Wulf, M.; Bailer, S.M.; Piwowarczyk, A.; Ehrhardt, A. Cold Atmospheric Plasma as Antiviral Therapy—Effect on Human Herpes Simplex Virus Type 1. J. Gen. Virol. 2020, 101, 208–215. [Google Scholar] [CrossRef]
- Li, W.; Luo, Z.; Yan, C.-Y.; Wang, X.-H.; He, Z.-J.; Ouyang, S.-H.; Yan, C.; Liu, L.-F.; Zhou, Q.-Q.; Mu, H.-L.; et al. Autophagic Degradation of PML Promotes Susceptibility to HSV-1 by Stress-Induced Corticosterone. Theranostics 2020, 10, 9032–9049. [Google Scholar] [CrossRef] [PubMed]
- Frobert, E.; Cortay, J.-C.; Ooka, T.; Najioullah, F.; Thouvenot, D.; Lina, B.; Morfin, F. Genotypic Detection of Acyclovir-Resistant HSV-1: Characterization of 67 ACV-Sensitive and 14 ACV-Resistant Viruses. Antiviral. Res. 2008, 79, 28–36. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Zupin, L.; Rupel, K.; Ottaviani, G.; Poropat, A.; Di Lenarda, R.; Biasotto, M.; Crovella, S. Effect of Laser Therapy on Defensins’ Gene Expression in TR146 Epithelial Cell Line. Protein Pept. Lett. 2018, 25, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Zupin, L.; Ottaviani, G.; Rupel, K.; Biasotto, M.; Zacchigna, S.; Crovella, S.; Celsi, F. Analgesic Effect of Photobiomodulation Therapy: An in Vitro and in Vivo Study. J. Biophotonics 2019, 12, e201900043. [Google Scholar] [CrossRef] [PubMed]
- Zupin, L.; Barbi, E.; Sagredini, R.; Ottaviani, G.; Crovella, S.; Celsi, F. In Vitro Effects of Photobiomodulation Therapy on 50B11 Sensory Neurons: Evaluation of Cell Metabolism, Oxidative Stress, Mitochondrial Membrane Potential (MMP), and Capsaicin-Induced Calcium Flow. J. Biophotonics 2021, 14, e202000347. [Google Scholar] [CrossRef] [PubMed]
- Rupel, K.; Zupin, L.; Colliva, A.; Kamada, A.; Poropat, A.; Ottaviani, G.; Gobbo, M.; Fanfoni, L.; Gratton, R.; Santoro, M.; et al. Photobiomodulation at Multiple Wavelengths Differentially Modulates Oxidative Stress In Vitro and In Vivo. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Enwemeka, C.S.; Bumah, V.V.; Mokili, J.L. Pulsed Blue Light Inactivates Two Strains of Human Coronavirus. J. Photochem. Photobiol. B Biol. 2021, 222, 112282. [Google Scholar] [CrossRef]
- Stasko, N.; Kocher, J.F.; Annas, A.; Henson, I.; Seitz, T.S.; Miller, J.M.; Arwood, L.; Roberts, R.C.; Womble, T.M.; Keller, E.G.; et al. Visible Blue Light Inhibits Infection and Replication of SARS-CoV-2 at Doses That Are Well-Tolerated by Human Respiratory Tissue. Sci. Rep. 2021, 11, 20595. [Google Scholar] [CrossRef]
- Oh, P.; Kang, K.W.; Ryu, S.R.; Lim, S.; Sohn, M.; Lee, S.; Jeong, H. Evaluation of Photobiogoverning Role of Blue Light Irradiation on Viral Replication. J. Photochem. Photobiol. 2021, php.13514. [Google Scholar] [CrossRef] [PubMed]
- De Santis, R.; Luca, V.; Faggioni, G.; Fillo, S.; Stefanelli, P.; Rezza, G.; Lista, F. Rapid Inactivation of SARS-CoV-2 with LED Irradiation of Visible Spectrum Wavelenghts. J. Photochem. Photobiol. 2020, 8, 100082. [Google Scholar] [CrossRef] [PubMed]
- Rathnasinghe, R.; Jangra, S.; Miorin, L.; Schotsaert, M.; Yahnke, C.; Garcίa-Sastre, A. The Virucidal Effects of 405 Nm Visible Light on SARS-CoV-2 and Influenza A Virus. Sci. Rep. 2021, 11, 19470. [Google Scholar] [CrossRef] [PubMed]
- Vatter, P.; Hoenes, K.; Hessling, M. Photoinactivation of the Coronavirus Surrogate Phi6 by Visible Light. Photochem. Photobiol. 2021, 97, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Baillie, G.J.; Vetter, I. Neuronal Cell Lines as Model Dorsal Root Ganglion Neurons: A Transcriptomic Comparison. Mol. Pain 2016, 12, 174480691664611. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zupin, L.; Crovella, S. Blue Laser Light Counteracts HSV-1 in the SH-SY5Y Neuronal Cell Model of Infection. Life 2022, 12, 55. https://doi.org/10.3390/life12010055
Zupin L, Crovella S. Blue Laser Light Counteracts HSV-1 in the SH-SY5Y Neuronal Cell Model of Infection. Life. 2022; 12(1):55. https://doi.org/10.3390/life12010055
Chicago/Turabian StyleZupin, Luisa, and Sergio Crovella. 2022. "Blue Laser Light Counteracts HSV-1 in the SH-SY5Y Neuronal Cell Model of Infection" Life 12, no. 1: 55. https://doi.org/10.3390/life12010055
APA StyleZupin, L., & Crovella, S. (2022). Blue Laser Light Counteracts HSV-1 in the SH-SY5Y Neuronal Cell Model of Infection. Life, 12(1), 55. https://doi.org/10.3390/life12010055