
Site-Specific and Common Prostate Cancer Metastasis Genes as Suggested by Meta-Analysis of Gene Expression Data


Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Expression Datasets
2.2. Meta-Analysis of Gene Expression Data and Enrichment Analysis
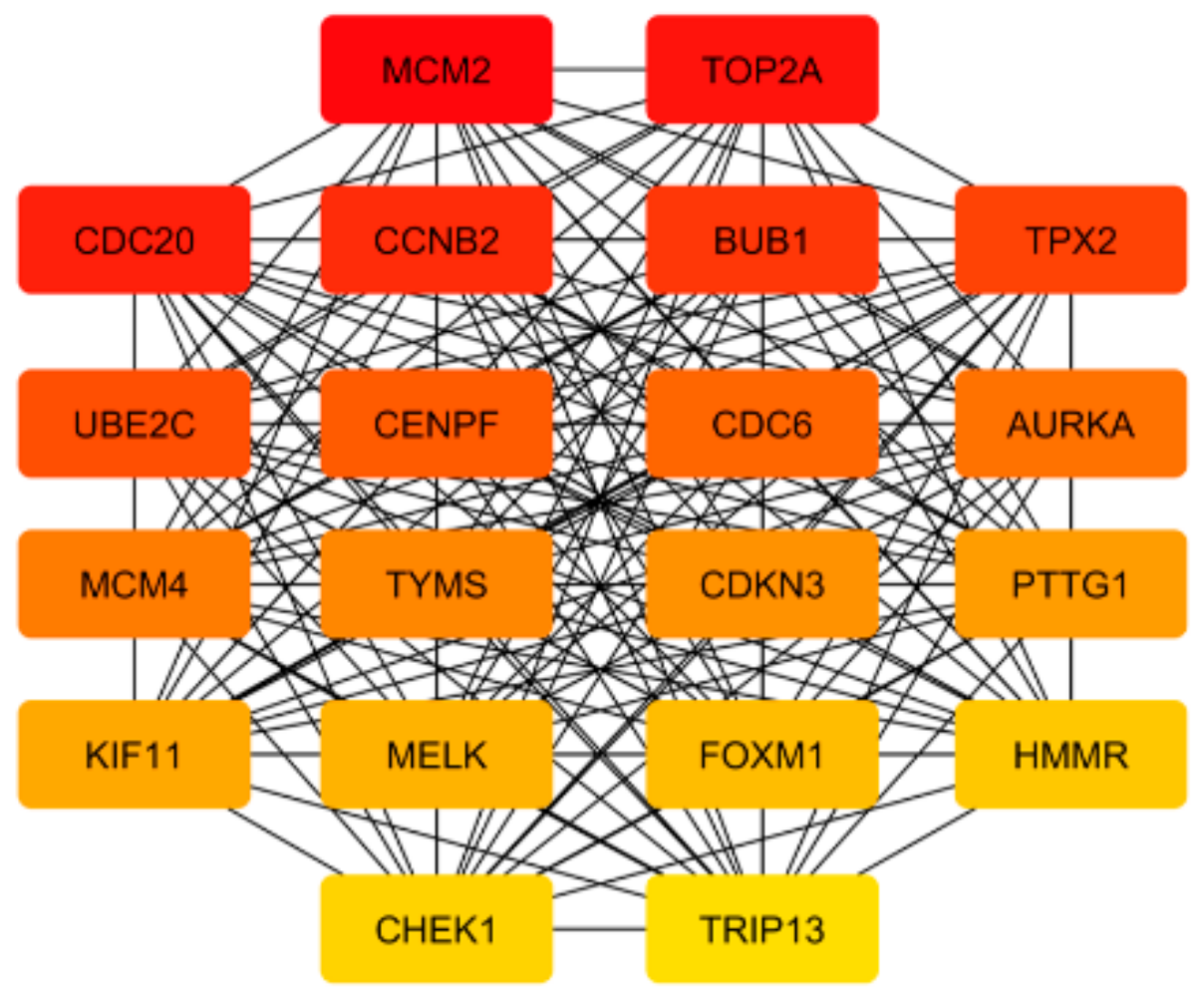
2.3. Visualization of Metastasis Genes Networks and Identification of the Hub Genes
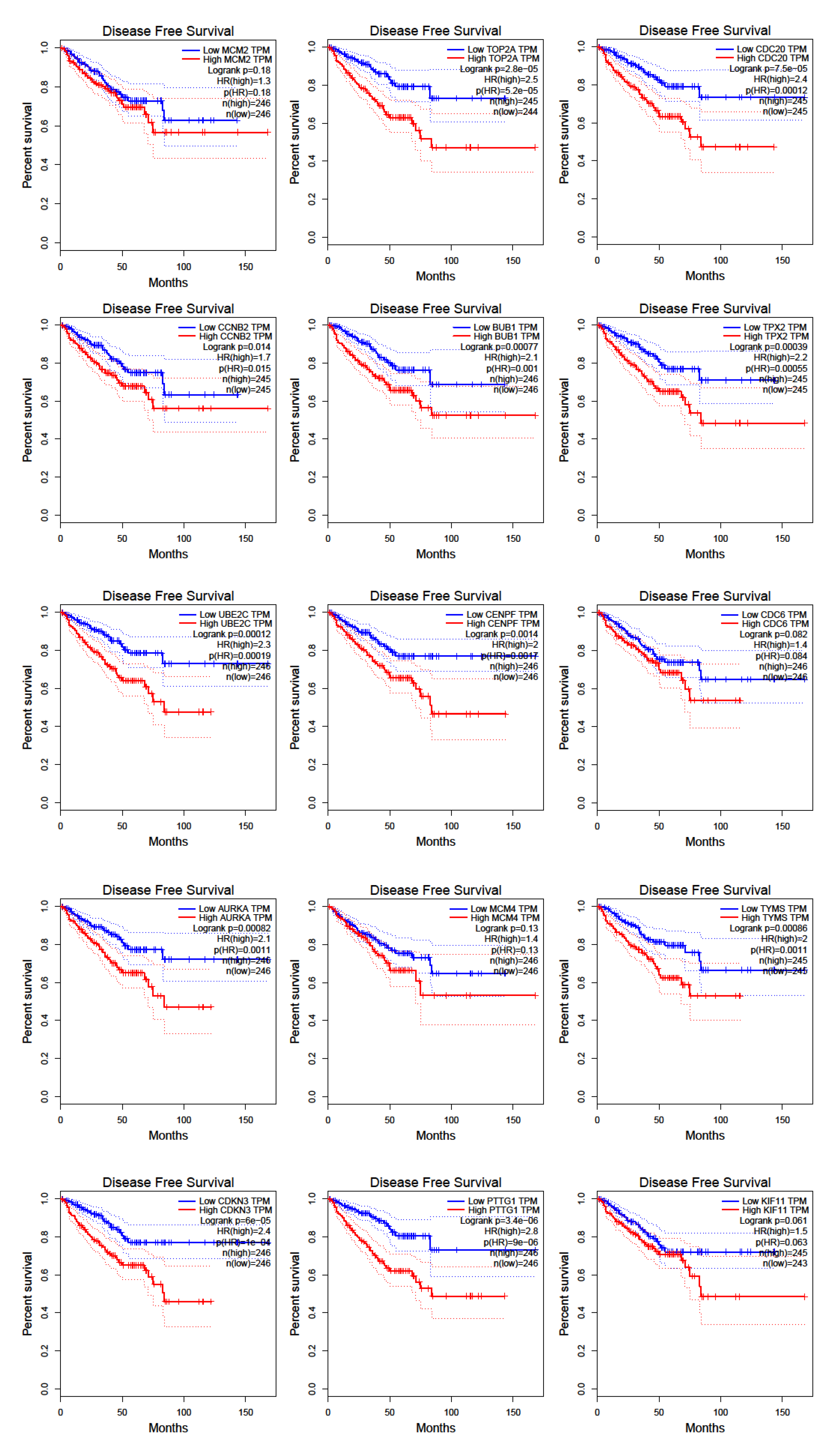
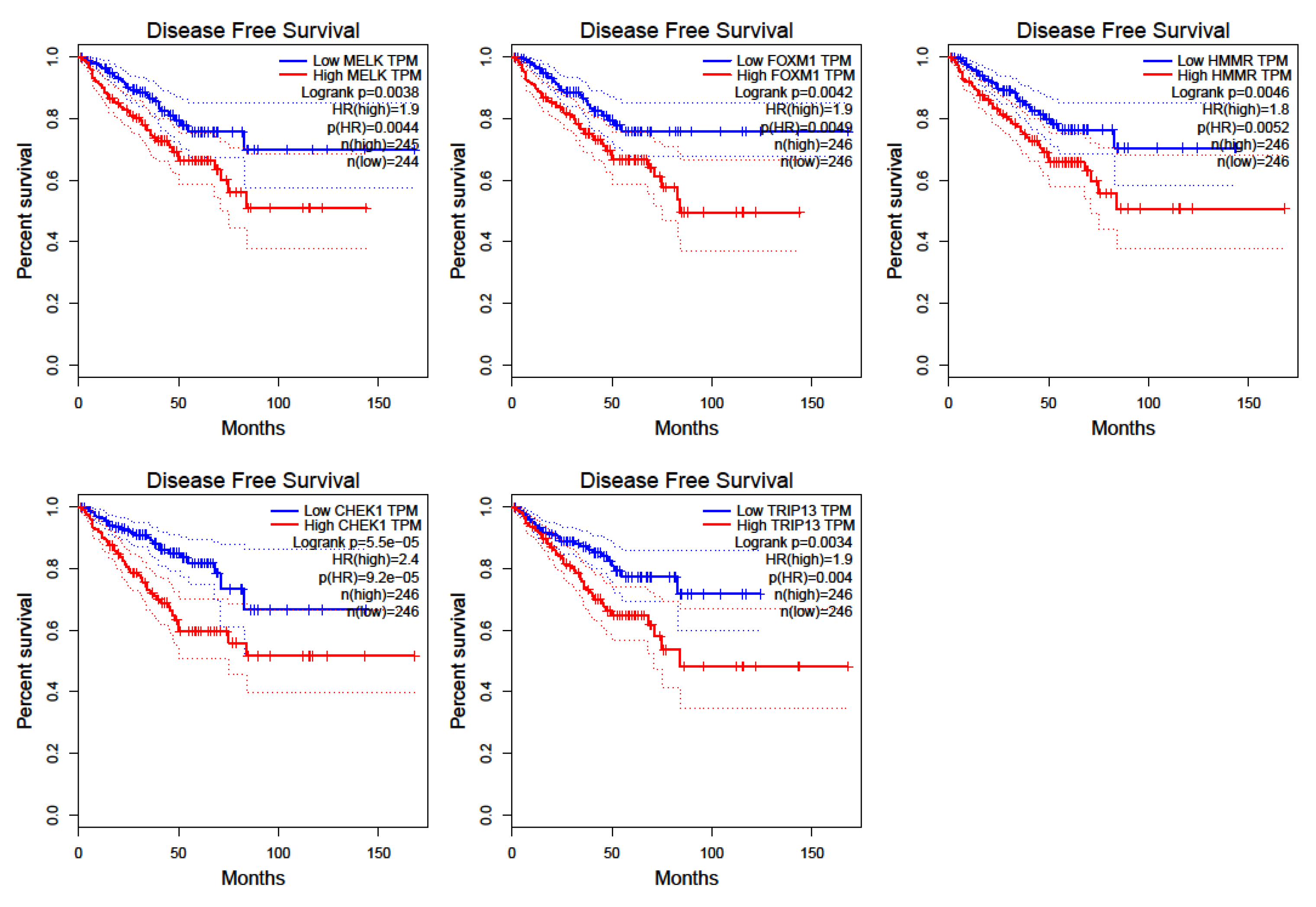
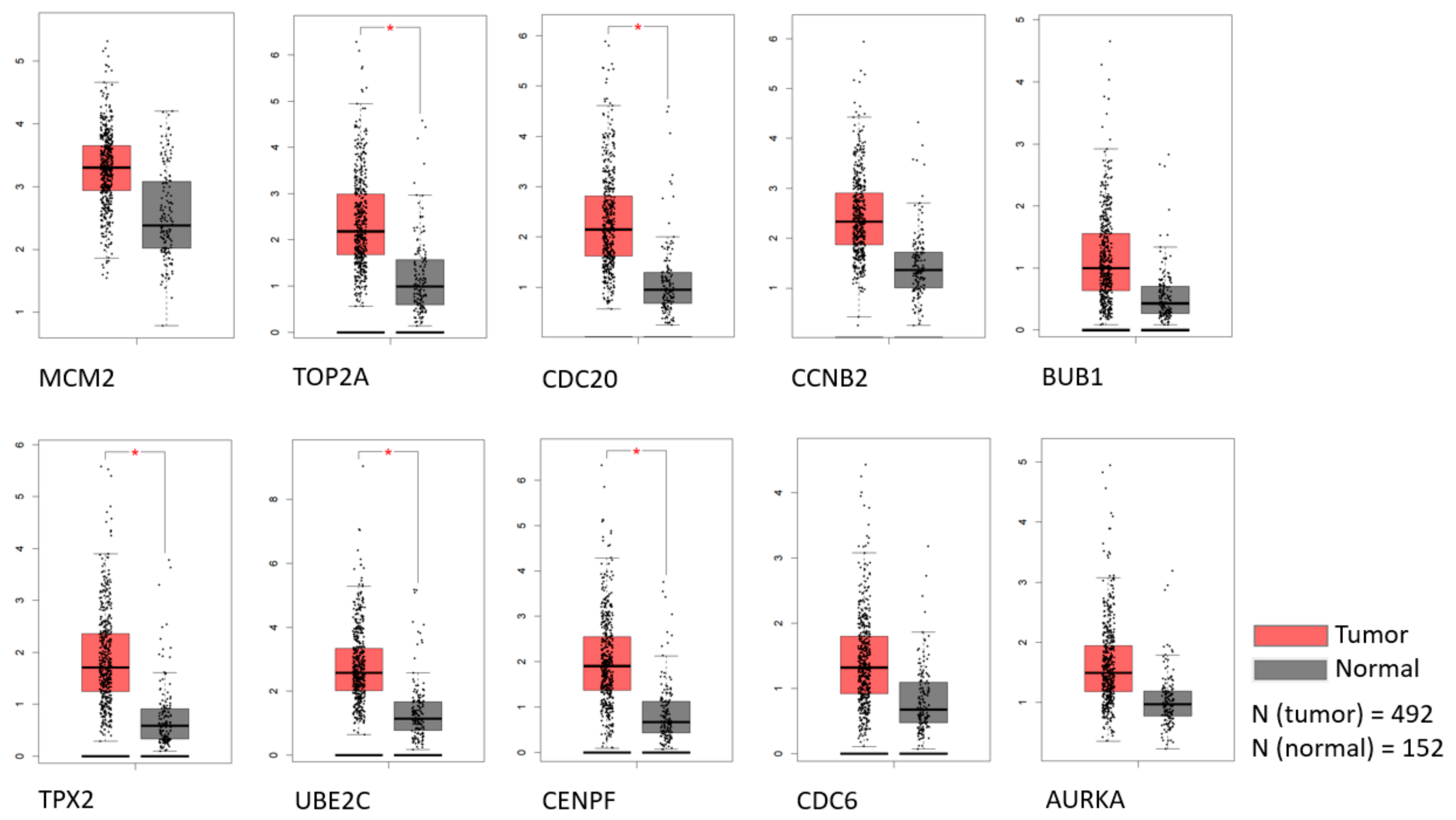
2.4. Survival and Expression Analysis for the Hub Genes
3. Results
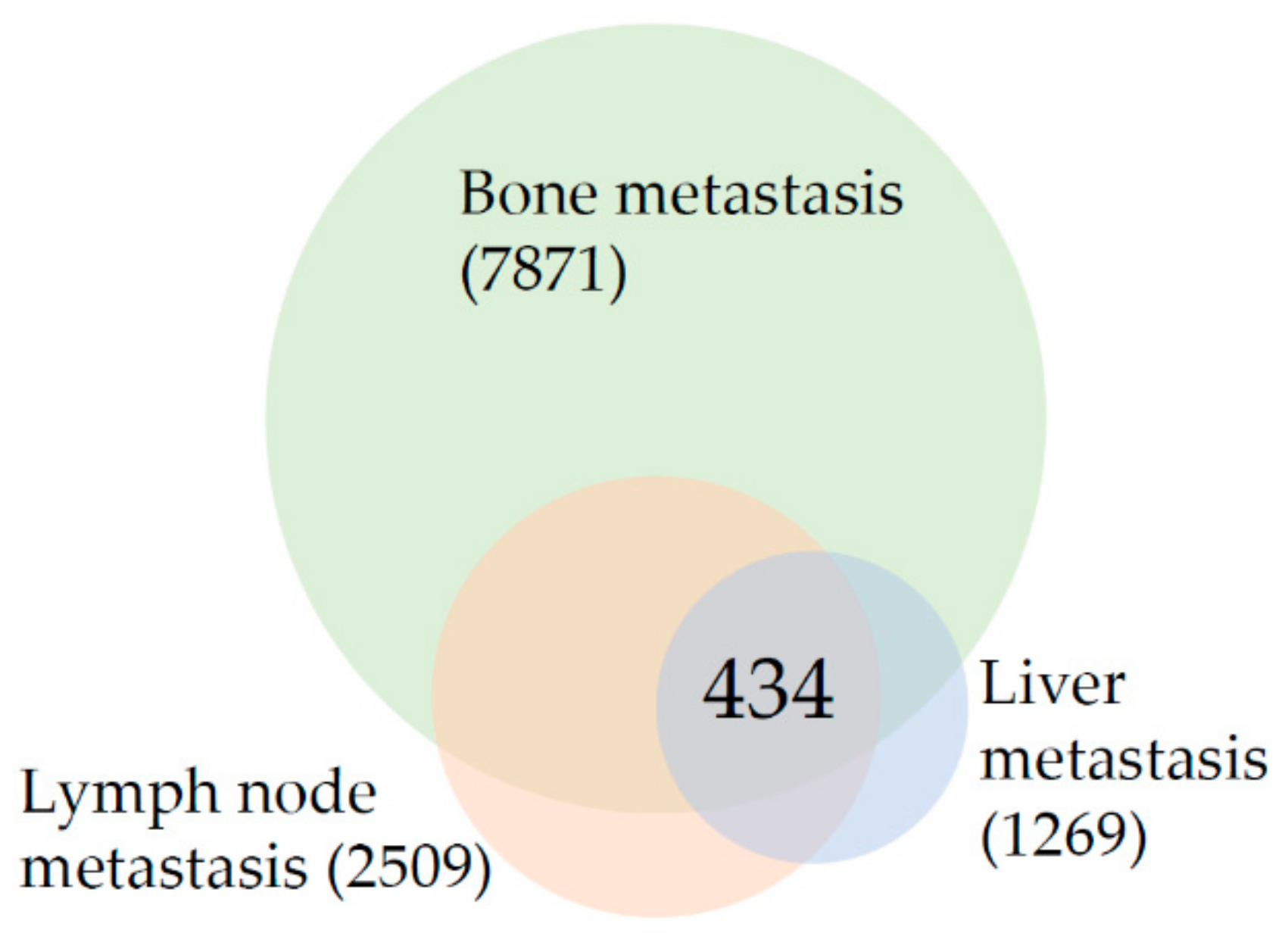
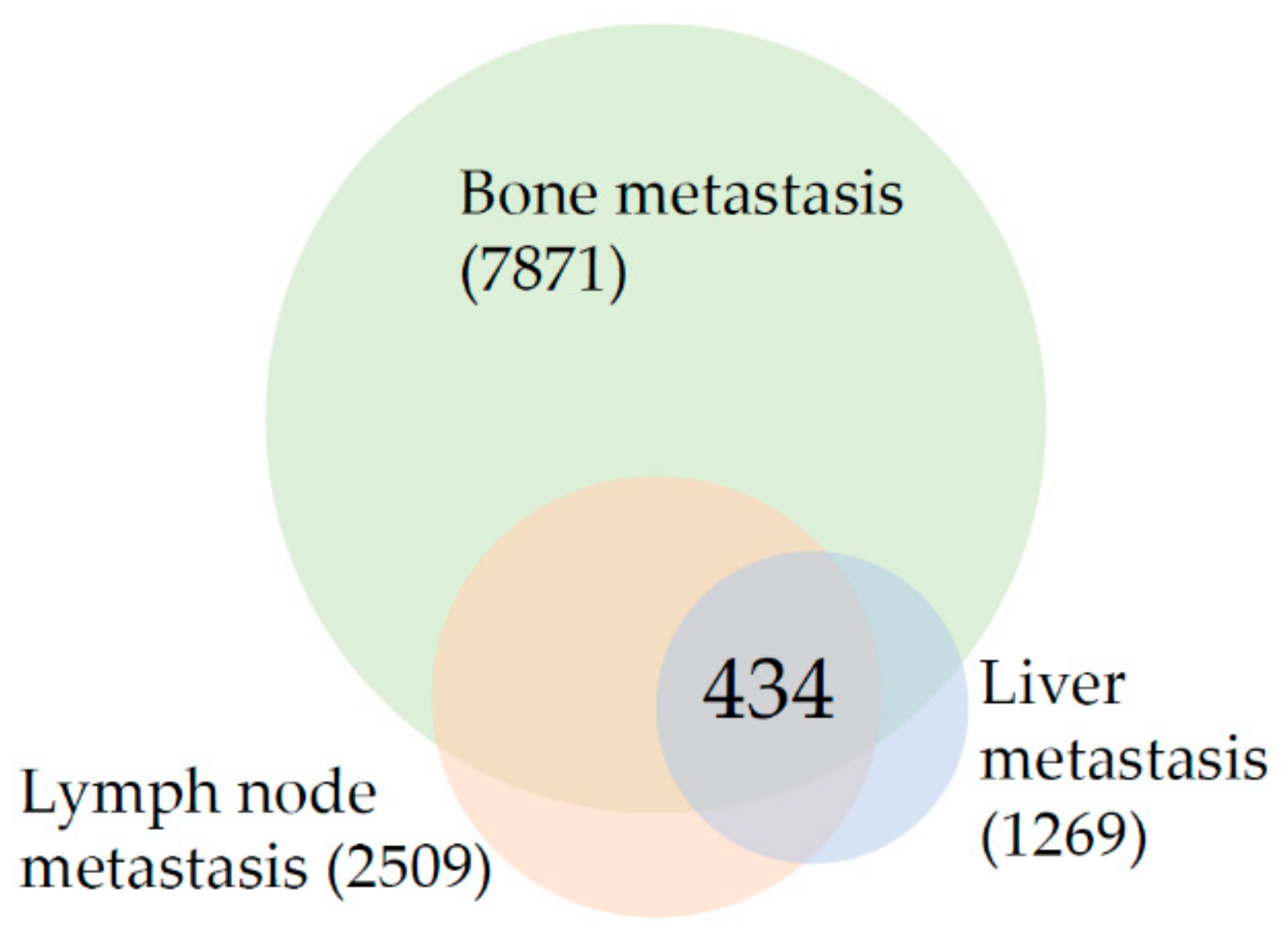
3.1. The Top Ten Most Changed Genes Shared by Metastasis from All Analyzed Sites Belong to the Class of Filaments and Proteins Involved in Bone and Prostate Biology
3.2. Reorganization of Focal Adhesions and Changes in Androgen Receptor Signaling Are Common Characteristics of Prostate Cancer Metastasis Regardless of the Target Organ
3.3. Results Suggest That Transcriptional Landscape of Bone Metastasis Is Profoundly Rewired in Comparison to Lymph Node and Liver Metastasis
3.4. Lymph Node Metastasis Are Characterized by Changes in Signaling Networks While the Liver Metastasis Transcriptional Program Is Reminiscent of Processes That Take Place in the Target Organ
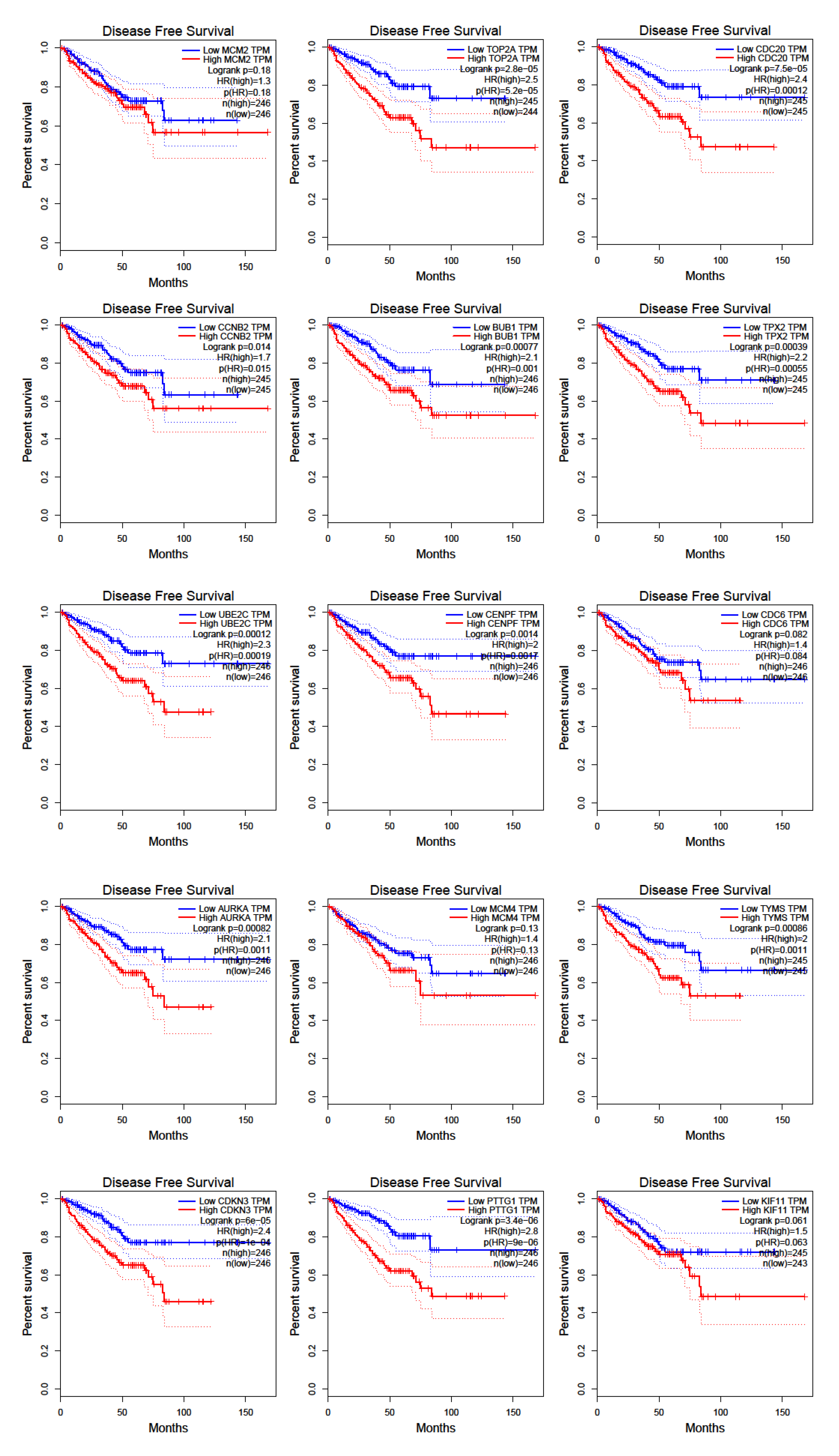
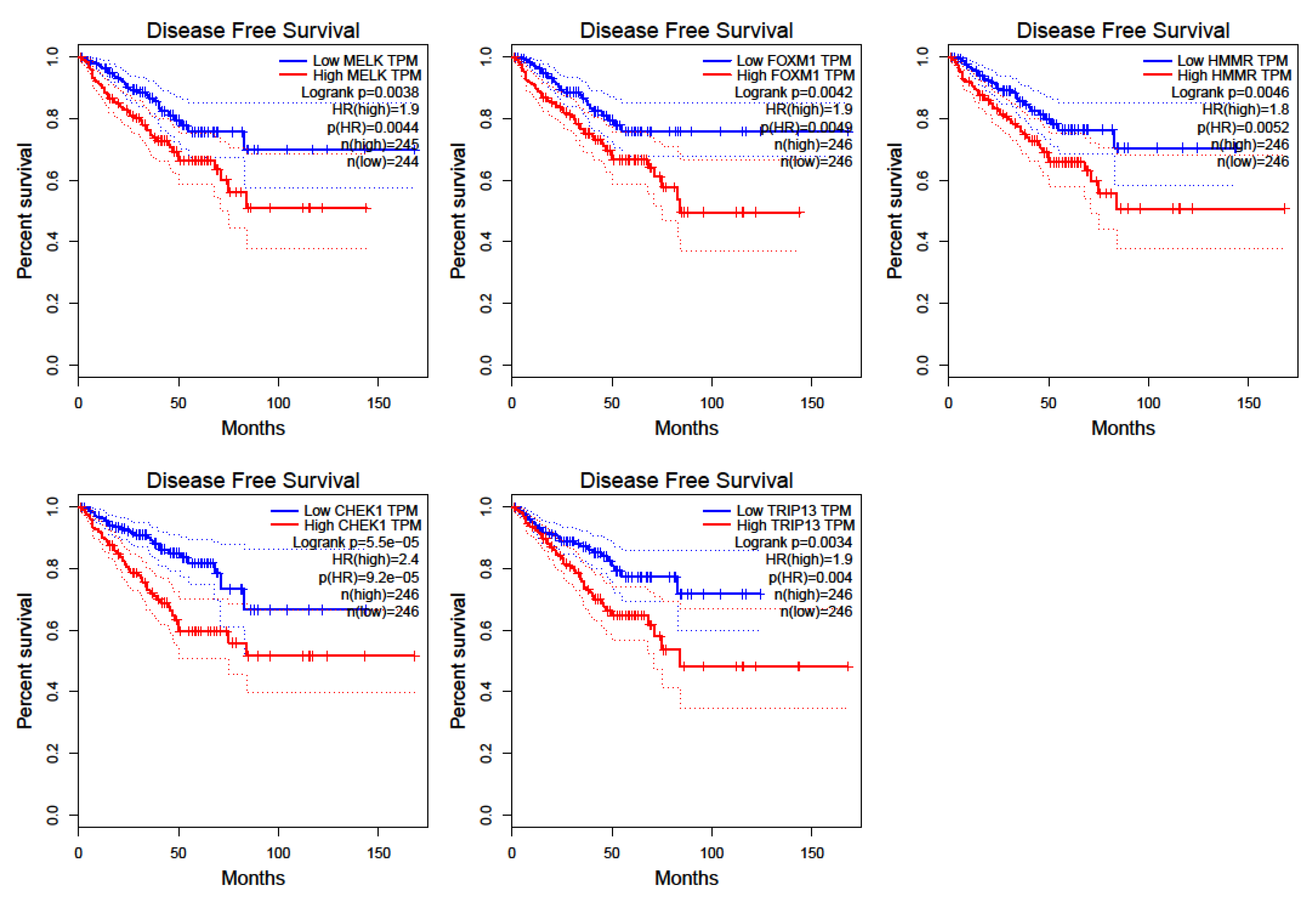
3.5. Prostate Cancer Metastasis Hub Genes and Their Involvement in Patient Disease-Free Survival
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559. [Google Scholar] [CrossRef]
- Weber, G.F. Why does cancer therapy lack effective anti-metastasis drugs? Cancer Lett. 2013, 328, 207. [Google Scholar] [CrossRef]
- Zheng, G.; Ma, Y.; Zou, Y.; Yin, A.; Li, W.; Dong, D. HCMDB: The human cancer metastasis database. Nucleic Acids Res. 2018, 46(D1), D950. [Google Scholar] [CrossRef] [Green Version]
- Hartung, F.; Wang, Y.; Aronow, B.; Weber, G.F. A core program of gene expression characterizes cancer metastases. Oncotarget 2017, 8, 10216. [Google Scholar] [CrossRef]
- Hartung, F.; Patil, A.; Meshram, R.J.; Weber, G.F. Gene expression signatures of site-specificity in cancer metastases. Clin. Exp. Metastasis 2019, 37, 159. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Zieren, R.C.; Xue, W.; de Reijke, T.M.; Pienta, K.J. Metastatic prostate cancer remains incurable, why? Asian J. Urol. 2019, 6, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Bi, R.; Zhu, Z.; Li, S.; Xu, B.; Rather, W.A.; Wang, C. A surveillance, epidemiology and end results database analysis of the prognostic value of organ-specific metastases in patients with advanced prostatic adenocarcinoma. Oncol. Lett. 2019, 18, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Castelli, G.; Pelosi, E. Cellular and Molecular Mechanisms Underlying Prostate Cancer Development: Therapeutic Implications. Medicines 2019, 6, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaržija, I. Post-Translational Modifications that Drive Prostate Cancer Progression. Biomolecules 2021, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets-Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists. Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Chandran, U.R.; Ma, C.; Dhir, R.; Bisceglia, M.; Lyons-Weiler, M.; Liang, W.; Michalopoulos, G.; Becich, M.; Monzon, F.A. Gene expression profiles of prostate cancer reveal involvement of multiple molecular pathways in the metastatic process. BMC Cancer 2007, 12, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative Genomic Profiling of Human Prostate Cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Wang, H.; He, H.H.; Chen, S.; He, L.; Ma, F.; Mucci, L.; Wang, Q.; Fiore, C.; Sowalsky, A.G.; et al. ERG induces androgen receptor-mediated regulation of SOX9 in prostate cancer. J. Clin. Investig. 2013, 123, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, R.; Marije Hoogland, A.; Dits, N.; Verhoef, E.I.; Kweldam, C.; Waranecki, P.; Bangma, C.H.; van Leenders, G.J.L.H.; Jenster, G. Novel long non-coding RNAs are specific diagnostic and prognostic markers for prostate cancer. Oncotarget 2015, 6, 4036–4050. [Google Scholar] [CrossRef]
- Kumar, A.; Coleman, I.; Morrissey, C.; Zhang, X.; True, L.D.; Gulati, R.; Etzioni, R.; Bolouri, H.; Montgomery, B.; White, T.; et al. Substantial interindividual and limited intraindividual genomic diversity among tumors from men with metastatic prostate cancer. Nat. Med. 2016, 22, 369–378. [Google Scholar] [CrossRef]
- Toro-Domínguez, D.; Martorell-Marugán, J.; López-Domínguez, R.; García-Moreno, A.; González-Rumayor, V.; Alarcón-Riquelme, M.E.; Carmona-Sáez, P. ImaGEO: Integrative gene expression meta-analysis from GEO database. Bioinformatics 2019, 35, 880–882. [Google Scholar] [CrossRef]
- Pirooznia, M.; Nagarajan, V.; Deng, Y. GeneVenn—A web application for comparing gene lists using Venn diagrams. Bioinformation 2007, 1, 420–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. S4), S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, F.; Hartwig, J.H.; Stossel, T.P.; Szymanski, P.T. Ca2+ and calmodulin regulate the binding of filamin A to actin filaments. J. Biol. Chem. 2005, 280, 32426–32433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, B.; Zheng, X.F.; Jiang, S.D.; Zhou, Z.Z.; Xu, W.N.; Zheng, H.L.; Wang, C.D.; Zhang, X.L.; Jiang, L.S. Chordin-like 1 improves osteogenesis of bone marrow mesenchymal stem cells through enhancing BMP4-SMAD pathway. Front. Endocrinol. 2019, 10, 360. [Google Scholar] [CrossRef]
- Su, N.; Chen, M.; Chen, S.; Li, C.; Xie, Y.; Zhu, Y.; Zhang, Y.; Zhao, L.; He, Q.; Du, X.; et al. Overexpression of H1 calponin in osteoblast lineage cells leads to a decrease in bone mass by disrupting osteoblast function and promoting osteoclast formation. J. Bone Miner. Res. 2013, 28, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Pathak, B.R.; Breed, A.A.; Nakhawa, V.H.; Jagtap, D.D.; Mahale, S.D. Growth inhibition mediated by PSP94 or CRISP-3 is prostate cancer cell line specific. Asian J. Androl. 2010, 12, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Zou, H.; Wang, H.; Li, Q.; Yu, D. Identification of Key Gene Signatures Associated With Bone Metastasis in Castration-Resistant Prostate Cancer Using Co-Expression Analysis. Front. Oncol. 2021, 10, 571524. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wen, Y.; Xuan, C.; Chen, Q.; Xiang, Q.; Wang, J.; Liu, Y.; Luo, L.; Zhao, S.; Deng, Y.; et al. Identifying the key genes and microRNAs in prostate cancer bone metastasis by bioinformatics analysis. FEBS Open Bio. 2020, 10, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ding, Y.; Lu, W.; Zhang, K.; Wang, F.; Ding, G.; Wang, J. Comparison of metastatic castration-resistant prostate cancer in bone with other sites: Clinical characteristics, molecular features and immune status. PeerJ. 2021, 9, e11133. [Google Scholar] [CrossRef]
- Shamsara, E.; Shamsara, J. Bioinformatics analysis of the genes involved in the extension of prostate cancer to adjacent lymph nodes by supervised and unsupervised machine learning methods: The role of SPAG1 and PLEKHF2. Genomics 2020, 112, 3871–3882. [Google Scholar] [CrossRef]
- Xu, N.; Chen, S.-H.; Lin, T.-T.; Cai, H.; Ke, Z.-B.; Dong, R.-N.; Huang, P.; Li, X.-D.; Chen, Y.-H.; Zheng, Q.-S. Development and validation of hub genes for lymph node metastasis in patients with prostate cancer. J. Cell Mol. Med. 2020, 24, 4402–4414. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, D.; Spring, D.J.; Depinho, R.A. Genetics and biology of prostate cancer. Genes Dev. 2018, 32, 1105–1140. [Google Scholar] [CrossRef] [Green Version]
- Li, F.X.Z.; Liu, J.J.; Xu, F.; Lin, X.; Zhong, J.Y.; Wu, F.; Yuan, L.Q. Role of tumor-derived exosomes in bone metastasis (Review). Oncol. Lett. 2019, 18, 3935–3945. [Google Scholar] [CrossRef]
- Gao, Y.; Bado, I.; Wang, H.; Zhang, W.; Rosen, J.M.; Zhang, X.H.F. Metastasis Organotropism: Redefining the Congenial Soil. Dev. Cell 2019, 49, 375–391. [Google Scholar] [CrossRef]
- Kruger, T.E.; Miller, A.H.; Godwin, A.K.; Wang, J. Bone sialoprotein and osteopontin in bone metastasis of osteotropic cancers. Crit. Rev. Oncol. Hematol. 2014, 89, 330–341. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Gilkes, D.M. The contribution of the immune system in bone metastasis pathogenesis. Int. J. Mol. Sci. 2019, 20, 999. [Google Scholar] [CrossRef] [Green Version]
- Zlotnik, A.; Burkhardt, A.M.; Homey, B. Homeostatic chemokine receptors and organ-specific metastasis. Nat. Rev. Immunol. 2011, 11, 597–606. [Google Scholar] [CrossRef]
- Borsig, L.; Wolf, M.J.; Roblek, M.; Lorentzen, A.; Heikenwalder, M. Inflammatory chemokines and metastasis-Tracing the accessory. Oncogene 2014, 33, 3217–3224. [Google Scholar] [CrossRef] [Green Version]
- Roca, H.; Jones, J.D.; Purica, M.C.; Weidner, S.; Koh, A.J.; Kuo, R.; Wilkinson, J.E.; Wang, Y.; Daignault-Newton, S.; Pienta, K.J.; et al. Apoptosis-induced CXCL5 accelerates inflammation and growth of prostate tumor metastases in bone. J. Clin. Investig. 2018, 128, 248–266. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, H.; Ivaska, J. Every step of the way: Integrins in cancer progression and metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [Green Version]
- Samaržija, I.; Dekanić, A.; Humphries, J.D.; Paradžik, M.; Stojanović, N.; Humphries, M.J.; Ambriović-Ristov, A. Integrin crosstalk contributes to the complexity of signalling and unpredictable cancer cell fates. Cancers 2020, 12, 1910. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.; Møller, M.; Haldrup, C.; Strand, S.H.; Vang, S.; Hedegaard, J.; Høyer, S.; Borre, M.; Ørntoft, T.; Sørensen, K.D. Exploring the transcriptome of hormone-naive multifocal prostate cancer and matched lymph node metastases. Br. J. Cancer 2018, 119, 1527–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calmon, M.F.; Jeschke, J.; Zhang, W.; Dhir, M.; Siebenkäs, C.; Herrera, A.; Tsai, H.C.; O’Hagan, H.M.; Pappou, E.P.; Hooker, C.M.; et al. Epigenetic silencing of neurofilament genes promotes an aggressive phenotype in breast cancer. Epigenetics 2015, 10, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Chang, X.; LeBron, C.; Nagpal, J.K.; Lee, J.; Huang, Y.; Yamashita, K.; Trink, B.; Ratovitski, E.A.; Sidransky, D. Neurofilament heavy polypeptide regulates the Akt-β-catenin pathway in human esophageal squamous cell carcinoma. PLoS ONE 2010, 5, e9003. [Google Scholar] [CrossRef]
- Yoshimura, T.; Hamada, T.; Hijioka, H.; Souda, M.; Hatanaka, K.; Yoshioka, T.; Yamada, S.; Tsutsui, M.; Umekita, Y.; Nakamura, N.; et al. PCP4/PEP19 promotes migration, invasion and adhesion in human breast cancer MCF-7 and T47D cells. Oncotarget 2016, 7, 49065–49074. [Google Scholar] [CrossRef] [Green Version]
- Arentz, G.; Chataway, T.; Price, T.J.; Izwan, Z.; Hardi, G.; Cummins, A.G.; Hardingham, J.E. Desmin expression in colorectal cancer stroma correlates with advanced stage disease and marks angiogenic microvessels. Clin. Proteom. 2011, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Edfeldt, K.; Hellman, P.; Westin, G.; Stalberg, P. A plausible role for actin gamma smooth muscle 2 (ACTG2) in small intestinal neuroendocrine tumorigenesis. BMC Endocr. Disord. 2016, 16, 19. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Liu, Z.G.; Shi, M.Q.; Yu, H.Z.; Jiang, X.Y.; Yang, A.H.; Fu, X.S.; Xu, Y.; Yang, S.; Ni, H.; et al. Identification of ACTG2 functions as a promoter gene in hepatocellular carcinoma cells migration and tumor metastasis. Biochem. Biophys. Res. Commun. 2017, 491, 537–544. [Google Scholar] [CrossRef]
- Cioni, B.; Zwart, W.; Bergman, A.M. Androgen receptor moonlighting in the prostate cancer microenvironment. Endocr. Relat. Cancer 2018, 25, R331–R349. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Qiu, Y. Role of androgen receptor splice variants in prostate cancer metastasis. Asian J. Urol. 2016, 3, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.Y.; Jan, Y.J.; Kuo, L.K.; Wang, B.J.; Huo, C.; Jiang, S.S.; Chen, S.C.; Kuo, Y.Y.; Chang, C.R.; Chuu, C.P. Elevation of androgen receptor promotes prostate cancer metastasis by induction of epithelial-mesenchymal transition and reduction of KAT5. Cancer Sci. 2018, 109, 3564–3574. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Lin, M.; Cheng, Z.; Chen, Y.; Huang, Y.; Xu, K. Identification of key genes and multiple molecular pathways of metastatic process in prostate cancer. PeerJ 2019, 7, e7899. [Google Scholar] [CrossRef]
- Xu, Y.H.; Deng, J.L.; Wang, G.; Zhu, Y.S. Long non-coding RNAs in prostate cancer: Functional roles and clinical implications. Cancer Lett. 2019, 464, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Su, Z.; Liang, X.; Huang, Y.; Lan, Z.; Jiang, X. Long non-coding rnas in prostate tumorigenesis and therapy (Review). Mol. Clin. Oncol. 2020, 13, 76. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEO Set | Reference | Platform | Samples | |||
|---|---|---|---|---|---|---|
| Primary Tumors | LN Met. | Liver Met. | Bone Met. | |||
| GSE6605, GSE6606 | Chandran et al. 2007 [13] | GPL8300 | 61 | 15 | 5 | - |
| GSE21034 | Taylor et al. 2010 [14] | GPL10264 | 131 | 7 | - | 2 |
| GSE32269 | Cai et al. 2013 [15] | GPL96 | 22 | - | - | 29 |
| GSE59745 | Böttcher et al. 2015 [16] | GPL5175 | 18 | 12 | - | - |
| GSE77930 | Kumar et al. 2016 [17] | GPL15659 | 22 | 69 | 21 | 20 |
| Bone Metastasis | Lymph Node Metastases | Liver Metastasis |
|---|---|---|
| COL11A1, SPP1, HBB, IBSP, HBD, FABP4, MMP13, TNFAIP6, COL5A2, LRRC15, S100A8, MMP9, PTX3, TOP2A, FMO3, AKR1C3, COL1A1, SLPI, CA1, COL3A1, ENPEP, GMFG, COL1A2, CD36, LPL, OMD, APOE, SERPINE2, CFH, IGF2BP2, AHSP, TPX2, OLFML2B, CYP26A1, KIF20A, MEPE, COL10A1, MKI67, CTSZ, BUB1, DMP1, DPT, COL5A1, PPBP, IFI27, SULT1B1, COL4A1, STAB1, KIF4A, ALAS2 | CDKN3, THOC5, CCNE2, PPP3CA, TPH1, RFC5, UBE2C, HIST1H4L, FABP4, PAK2, PMAIP1, ESM1, MPHOSPH9, RAD23B, MVD, CENPF, SLC26A3, CD36, SPP1, RIPK2, RFPL3S, HMGCS1, TIMM8A, VDR, KCTD20, ATP8A2, NTNG1, BUB1, MAGEA12, CDC6, TOP2A, UBFD1, PTTG1, OSGIN2, COL9A3, KRR1, COL11A1, EZH2, ANGPT2, THBS2, FGF12, FOXM1, MYL4, KIF3A, CDKN2A, TPX2, NCBP1, MLF1, SEL1L, LMNB1 | FGB, FGG, PCK1, GC, FABP1, CRP, HP, CFHR4, APOC3, TM4SF4, CDKN3, APOB, APOA1, FGL1, RBP4, PEG10, HPGD, UGT2B7, ARG1, PLG, F2, FGA, SERPINC1, HBD, ALDOB, ALB, AMBP, SLC1A2, HIST1H4L, SPP1, AKR1C4, KNG1, KIF11, NMU, SERPINA1, COL9A3, APCS, IGFBP1, CPS1, F9, AADAC, MKNK2, UBE2C, ASGR2, AHSG, C4BPA, KIF23, C8A, SAA1, HSPA6 |
| ACADL, PGM3, SORD, MT1E, ANXA3, SMARCA1, KIAA1324, CHRDL1, MYBPC1, RLN1, PAK1IP1, ANPEP, GULP1, ACTA2, JMJD1C, NR4A2, LIFR, HSD17B6, DHRS7, KLF6, EPHX2, KIF5C, SFRP1, IQGAP2, MYLK, DMXL1, NCAPD3, CPE, SLC4A4, GREB1, FOS, ALDH1A3, ZFP36, HGD, DES, AZGP1, NEFH, DPP4, ABCC4, TRPM8, KRT15, FOSB, MEIS2, SORBS1, PCP4, CNN1, MAOB, ACTG2, MSMB, MYH11 | LTF, FBLN1, EGR2, AKAP5, ABCA8, SLC20A2, EGR3, FHL1, KRT5, PAMR1, MYLK, BDNF, EYA4, CHRDL1, EPHB6, NR4A2, HDAC9, GPM6B, EYA1, ATP1A2, EDNRA, CXCL11, PTGS2, FGFR2, PTPRD, KRT15, IGFBP6, PLN, SPOCK3, RLN1, DKK1, MFAP4, PTN, MOXD1, MSMB, PTGDS, MMP7, MEIS1, FOXF1, SYNM, CNTN1, DES, TCF21, CNN1, PCP4, OLFM4, NEFH, PAGE4, ACTG2, MYH11 | NT5E, PHF14, SPOCK3, VWF, NOV, CPM, NR4A3, COL16A1, EGR2, FGFR2, DYNC1I1, VPS13D, DDR2, PLA2G4A, TAC1, EGR3, CDH11, KRT15, FRZB, ATP1A2, SFRP1, SULF1, FOXF1, EDNRA, MYL9, PCP4, SPARCL1, UBE2K, CHRDL1, IGFBP6, DES, MFAP4, PTGDS, ACTC1, KLK11, SLC20A2, PAGE4, KLRD1, FBLN1, NEFH, MSMB, CNN1, KLK2, HDAC9, RLN2, CNTN1, PKP2, ACTG2, F3, MYH11 |
| Category | Term | Count | Adj. p-Value |
|---|---|---|---|
| BP | Platelet aggregation (GO:0070527) | 8/33 | 0.0010 |
| Negative regulation of cellular macromolecule biosynthetic process (GO:2000113) | 30/512 | 0.0011 | |
| Homotypic cell-cell adhesion (GO:0034109) | 8/38 | 0.0011 | |
| Regulation of retinoic acid receptor signaling pathway (GO:0048385) | 5/13 | 0.0027 | |
| Neutrophil degranulation (GO:0043312) | 27/479 | 0.0027 | |
| CC | Cytoskeleton (GO:0005856) | 32/520 | 3.37 × 10−5 |
| Focal adhesion (GO:0005925) | 24/356 | 1.31 × 10−4 | |
| Ficolin-1-rich granule (GO:0101002) | 15/184 | 9.15 × 10−4 | |
| Microtubule cytoskeleton (GO:0015630) | 23/388 | 9.15 × 10−4 | |
| Chromatin (GO:0000785) | 19/296 | 0.0014 | |
| MF | Kinase binding (GO:0019900) | 30/418 | 5.99 × 10−6 |
| Protein kinase binding (GO:0019901) | 30/495 | 1.22 × 10−4 | |
| Cadherin binding (GO:0045296) | 19/313 | 0.0103 | |
| Metal ion binding (GO:0046872) | 23/442 | 0.0143 | |
| Kinesin binding (GO:0019894) | 5/28 | 0.0268 | |
| KEGG | Focal adhesion | 20/199 | 3.7 × 10−6 |
| Cell cycle | 14/124 | 6.72 × 10−5 | |
| Prostate cancer | 10/97 | 0.0042 | |
| Glycolysis/Gluconeogenesis | 8/68 | 0.0071 | |
| p53 signaling pathway | 8/72 | 0.0085 | |
| WikiPathways | Focal Adhesion WP306 | 19/198 | 2.17 × 10−5 |
| Cell Cycle WP179 | 14/120 | 4.16 × 10−5 | |
| miRNA regulation of prostate cancer signaling pathways WP3981 | 8/33 | 4.16 × 10−5 | |
| Androgen receptor signaling pathway WP138 | 12/90 | 4.37 × 10−5 | |
| DNA Damage Response WP707 | 10/68 | 1.24 × 10−4 |
| Category | Term | Count | Adj. p-Value |
|---|---|---|---|
| BP | Cellular protein modification process (GO:0006464) | 582/1001 | 3.21 × 10−31 |
| Neutrophil activation involved in immune response (GO:0002283) | 320/483 | 2.26 × 10−30 | |
| Neutrophil mediated immunity (GO:0002446) | 322/487 | 2.26 × 10−30 | |
| Neutrophil degranulation (GO:0043312) | 317/479 | 4.20 × 10−30 | |
| Positive regulation of gene expression (GO:0010628) | 459/771 | 7.62 × 10−28 | |
| CC | Focal adhesion (GO:0005925) | 245/356 | 3.51 × 10−27 |
| Secretory granule lumen (GO:0034774) | 217/317 | 1.00 × 10−23 | |
| Nuclear body (GO:0016604) | 350/618 | 1.37 × 10−16 | |
| Mitochondrion (GO:0005739) | 537/1026 | 4.30 × 10−16 | |
| Nucleolus (GO:0005730) | 375/676 | 4.63 × 10−16 | |
| MF | RNA binding (GO:0003723) | 794/1387 | 2.90 × 10−41 |
| Cadherin binding (GO:0045296) | 221/313 | 9.62 × 10−27 | |
| Protein kinase activity (GO:0004672) | 308/513 | 3.11 × 10−19 | |
| Protein homodimerization activity (GO:0042803) | 380/664 | 6.99 × 10−19 | |
| Protein kinase binding (GO:0019901) | 295/495 | 6.33 × 10−18 | |
| KEGG | Pathways in cancer | 335/530 | 7.73 × 10−27 |
| Human T-cell leukemia virus 1 infection | 155/219 | 4.57 × 10−19 | |
| Human papillomavirus infection | 214/330 | 3.89 × 10−19 | |
| PI3K-Akt signaling pathway | 222/354 | 2.39 × 10−17 | |
| Protein processing in the endoplasmic reticulum | 121/165 | 3.98 × 10−17 | |
| WikiPathways | VEGFA-VEGFR2 Signaling Pathway WP3888 | 166/236 | 1.57 × 10−19 |
| Focal Adhesion-PI3K-Akt-mTOR-signaling pathway WP3932 | 198/303 | 7.32 × 10−18 | |
| Focal Adhesion WP306 | 141/198 | 1.17 × 10−17 | |
| EGF/EGFR Signaling Pathway WP437 | 120/162 | 2.70 × 10−17 | |
| TGF-beta Signaling Pathway WP366 | 102/132 | 5.84 × 10−17 |
| Category | Term | Count | Adj. p-Value |
|---|---|---|---|
| BP | Regulation of transcription from RNA polymerase II promoter (GO:0006357) | 303/1478 | 9.34 × 10−16 |
| Positive regulation of transcription, DNA-templated (GO:0045893) | 241/1120 | 8.53 × 10−15 | |
| Positive regulation of transcription from RNA polymerase II promoter (GO:0045944) | 195/848 | 8.73 × 10−15 | |
| Negative regulation of apoptotic process (GO:0043066) | 130/485 | 9.64 × 10−15 | |
| Regulation of cell migration (GO:0030334) | 96/316 | 2.69 × 10−14 | |
| CC | Focal adhesion (GO:0005925) | 110/356 | 1.26 × 10−17 |
| Cytoskeleton (GO:0005856) | 127/520 | 8.73 × 10−12 | |
| Membrane raft (GO:0045121) | 46/119 | 5.95 × 10−11 | |
| Perinuclear region of cytoplasm (GO:0048471) | 96/378 | 5.33 × 10−10 | |
| Platelet alpha granule (GO:0031091) | 36/90 | 3.67 × 10−9 | |
| MF | Protein kinase binding (GO:0019901) | 148/49 | 3.44 × 10−22 |
| Kinase binding (GO:0019900) | 124/418 | 2.94 × 10−18 | |
| Cadherin binding (GO:0045296) | 101/313 | 1.19 × 10−17 | |
| RNA binding (GO:0003723) | 274/1387 | 4.87 × 10−13 | |
| RNA polymerase II regulatory region sequence-specific DNA binding (GO:0000977) | 116/460 | 1.11 × 10−11 | |
| KEGG | Pathways in cancer | 136/530 | 2.06 × 10−14 |
| Focal adhesion | 70/199 | 2.06 × 10−14 | |
| MAPK signaling pathway | 86/295 | 1.89 × 10−12 | |
| PI3K-Akt signaling pathway | 94/354 | 4.25 × 10−11 | |
| Human T-cell leukemia virus 1 infection | 67/219 | 7.58 × 10−11 | |
| WikiPathways | VEGFA-VEGFR2 Signaling Pathway WP3888 | 80/236 | 5.16 × 10−15 |
| Focal Adhesion WP306 | 69/198 | 8.89 × 10−14 | |
| Integrated Breast Cancer Pathway WP1984 | 52/151 | 4.22 × 10−10 | |
| Cell Cycle WP179 | 44/120 | 9.84 × 10−10 | |
| Ebola Virus Pathway on Host WP4217 | 46/129 | 9.84 × 10−10 |
| Category | Term | Count | Adj. p-Value |
|---|---|---|---|
| BP | Cellular protein metabolic process (GO:0044267) | 76/484 | 7.17 × 10−10 |
| Platelet degranulation (GO:0002576) | 33/124 | 1.90 × 10−9 | |
| Regulated exocytosis (GO:0045055) | 35/148 | 1.07 × 10−8 | |
| Negative regulation of cellular process (GO:0048523) | 73/534 | 4.23 × 10−7 | |
| DNA metabolic process (GO:0006259) | 51/314 | 4.31 × 10−7 | |
| CC | Endoplasmic reticulum lumen (GO:0005788) | 49/270 | 5.10 × 10−9 |
| Focal adhesion (GO:0005925) | 58/356 | 5.10 × 10−9 | |
| Secretory granule lumen (GO:0034774) | 53/317 | 9.09 × 10−9 | |
| Perinuclear region of cytoplasm (GO:0048471) | 53/378 | 3.82 × 10−6 | |
| Cytoplasmic vesicle lumen (GO:0060205) | 26/129 | 8.41 × 10−6 | |
| MF | Protein homodimerization activity (GO:0042803) | 88/664 | 2.90 × 10−8 |
| Metal ion binding (GO:0046872) | 65/442 | 7.99 × 10−8 | |
| Transition metal ion binding (GO:0046914) | 56/399 | 5.30 × 10−6 | |
| Kinase binding (GO:0019900) | 56/418 | 2.01 × 10−5 | |
| Protein kinase binding (GO:0019901) | 60/495 | 1.73 × 10−4 | |
| KEGG | Cell cycle | 31/124 | 7.94 × 10−9 |
| Pathways in cancer | 72/530 | 1.08 × 10−7 | |
| Complement and coagulation cascades | 22/79 | 2.23 × 10−7 | |
| Focal adhesion | 37/199 | 2.27 × 10−7 | |
| Human T-cell leukemia virus 1 infection | 39/219 | 2.46 × 10−7 | |
| WikiPathways | VEGFA-VEGFR2 Signaling Pathway WP3888 | 48/236 | 1.93 × 10−10 |
| Cell Cycle WP179 | 31/120 | 2.20 × 10−9 | |
| Retinoblastoma Gene in Cancer WP2446 | 25/87 | 1.20 × 10−8 | |
| Nuclear Receptors Meta-Pathway WP2882 | 50/31 | 2.81 × 10−7 | |
| Human Complement System WP2806 | 24/97 | 4.75 × 10−7 |
| Gene Symbol | Full Name | Function |
|---|---|---|
| MCM2 | Minichromosome maintenance complex component 2 | Involved in the initiation of eukaryotic genome replication |
| TOP2A | Topoisomerase IIα | Controls the topology structure of DNA and cell cycle progression |
| CDC20 | Cell division cycle 20 | Regulatory protein interacting with several other proteins at multiple points in the cell cycle |
| CCNB2 | Cyclin B2 | Essential component of the cell cycle regulatory machinery |
| BUB1 | BUB1 mitotic checkpoint serine/threonine kinase | Serine/threonine-protein kinase that plays a central role in mitosis |
| TPX2 | TPX2 microtubule nucleation factor | Microtubule-associated protein linked to mitosis and spindle assembly |
| UBE2C | Ubiquitin-conjugating enzyme E2 C | Involved in ubiquitination; required for the destruction of mitotic cyclins and cell cycle progression |
| CENPF | Centromere protein F | Role in the centromere-kinetochore complex and chromosomal segregation |
| CDC6 | Cell division cycle 6 | Essential for the initiation of DNA replication |
| AURKA | Aurora kinase A | Cell cycle-regulated kinase involved in microtubule formation and/or stabilization at the spindle pole during chromosome segregation |
| MCM4 | Minichromosome maintenance complex component 4 | Essential for the initiation of eukaryotic genome replication |
| TYMS | Thymidylate synthetase | Catalyzes the methylation of deoxyuridylate to deoxythymidylate and maintains the dTMP pool critical for DNA replication and repair |
| CDKN3 | Cyclin-dependent kinase inhibitor 3 | Cyclin-dependent kinase inhibitor |
| PTTG1 | PTTG1 regulator of sister chromatid separation | Homolog of yeast securin proteins, which prevent separins from promoting sister chromatid separation |
| KIF11 | Kinesin family member 11 | Motor protein that belongs to the kinesin-like protein family |
| MELK | Maternal embryonic leucine zipper kinase | Serine/threonine-protein kinase involved in various processes such as cell cycle regulation, self-renewal of stem cells, apoptosis, and splicing regulation |
| FOXM1 | Forkhead box M1 | Transcriptional activator involved in cell proliferation |
| HMMR | Hyaluronan (HA) -mediated motility receptor | Binds native and fragmented HA, promotes its uptake |
| CHEK1 | Checkpoint kinase 1 | Required for checkpoint mediated cell cycle arrest in response to DNA damage or the presence of unreplicated DNA |
| TRIP13 | Thyroid hormone receptor interactor 13 | Interacts with thyroid hormone receptors |
| Category | Term | Count | Adj. p-Value |
|---|---|---|---|
| BP | Mitotic cell cycle phase transition (GO:0044772) | 9/221 | 7.22 × 10−11 |
| Regulation of mitotic cell cycle phase transition (GO:1901990) | 7/184 | 3.99 × 10−8 | |
| G1/S transition of the mitotic cell cycle (GO:0000082) | 5/105 | 4.03 × 10−6 | |
| Regulation of mitotic sister chromatid separation (GO:0010965) | 3/15 | 1.65 × 10−5 | |
| DNA metabolic process (GO:0006259) | 6/314 | 1.65 × 10−5 | |
| CC | Microtubule cytoskeleton (GO:0015630) | 8/388 | 7.67 × 10−8 |
| Spindle (GO:0005819) | 6/186 | 4.15 × 10−7 | |
| Spindle pole (GO:0000922) | 5/107 | 7.73 × 10−7 | |
| Condensed nuclear chromosome, centromeric region (GO:0000780) | 2/12 | 6.23 × 10−5 | |
| Mitotic spindle (GO:0072686) | 3/84 | 6.19 × 10−4 | |
| MF | Histone serine kinase activity (GO:0035174) | 2/7 | 0.0011 |
| Histone kinase activity (GO:0035173) | 2/9 | 0.0011 | |
| Kinase binding (GO:0019900) | 5/418 | 0.0011 | |
| Protein kinase binding (GO:0019901) | 5/495 | 0.0018 | |
| Protein serine/threonine kinase activity (GO:0004674) | 4/368 | 0.0059 | |
| KEGG | Cell cycle | 8/124 | 2.88 × 10−12 |
| Oocyte meiosis | 5/125 | 8.86 × 10−7 | |
| Human T-cell leukemia virus 1 infection | 4/219 | 2.76 × 10−4 | |
| Progesterone-mediated oocyte maturation | 3/99 | 4.42 × 10−4 | |
| Cellular senescence | 3/160 | 0.0014 | |
| WikiPathways | Cell Cycle WP179 | 8/120 | 3.37 × 10−12 |
| Gastric Cancer Network 1 WP2361 | 6/29 | 3.37 × 10−12 | |
| Retinoblastoma Gene in Cancer WP2446 | 5/87 | 2.25 × 10−7 | |
| Regulation of sister chromatid separation at the metaphase-anaphase transition WP4240 | 3/15 | 3.19 × 10−6 | |
| DNA Replication WP466 | 3/42 | 6.32 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samaržija, I. Site-Specific and Common Prostate Cancer Metastasis Genes as Suggested by Meta-Analysis of Gene Expression Data. Life 2021, 11, 636. https://doi.org/10.3390/life11070636
Samaržija I. Site-Specific and Common Prostate Cancer Metastasis Genes as Suggested by Meta-Analysis of Gene Expression Data. Life. 2021; 11(7):636. https://doi.org/10.3390/life11070636
Chicago/Turabian StyleSamaržija, Ivana. 2021. "Site-Specific and Common Prostate Cancer Metastasis Genes as Suggested by Meta-Analysis of Gene Expression Data" Life 11, no. 7: 636. https://doi.org/10.3390/life11070636
APA StyleSamaržija, I. (2021). Site-Specific and Common Prostate Cancer Metastasis Genes as Suggested by Meta-Analysis of Gene Expression Data. Life, 11(7), 636. https://doi.org/10.3390/life11070636