
Ticks, Hair Loss, and Non-Clinging Babies: A Novel Tick-Based Hypothesis for the Evolutionary Divergence of Humans and Chimpanzees


{kind=link}
Abstract
1. Introduction
2. Forest Fragmentation and Ticks
2.1. Forest Fragmentation in Hominin Paleoenvironments
- (1)
- (2)
2.2. Importance of Ticks
2.3. Ticks and Forest Fragmentation
2.4. Hominin Exposure to Ticks in Fragmented Paleoenvironments
3. Evolution of Alternative Anti-Tick Strategies
3.1. Hominin Hair Loss
Evolutionary Timing of Hominin Hair Loss
3.2. Chimpanzee Grooming
The Social Brain Hypothesis for Primate Allogrooming
4. The Locomotor Divergence of Humans and Chimpanzees
4.1. The Locomotor Repertoire of the LCA
4.2. The Hylobatian Model for the LCA
4.3. Evidence in Support of the Hylobatian Model
4.4. Hominin Straight-Legged Bipedalism
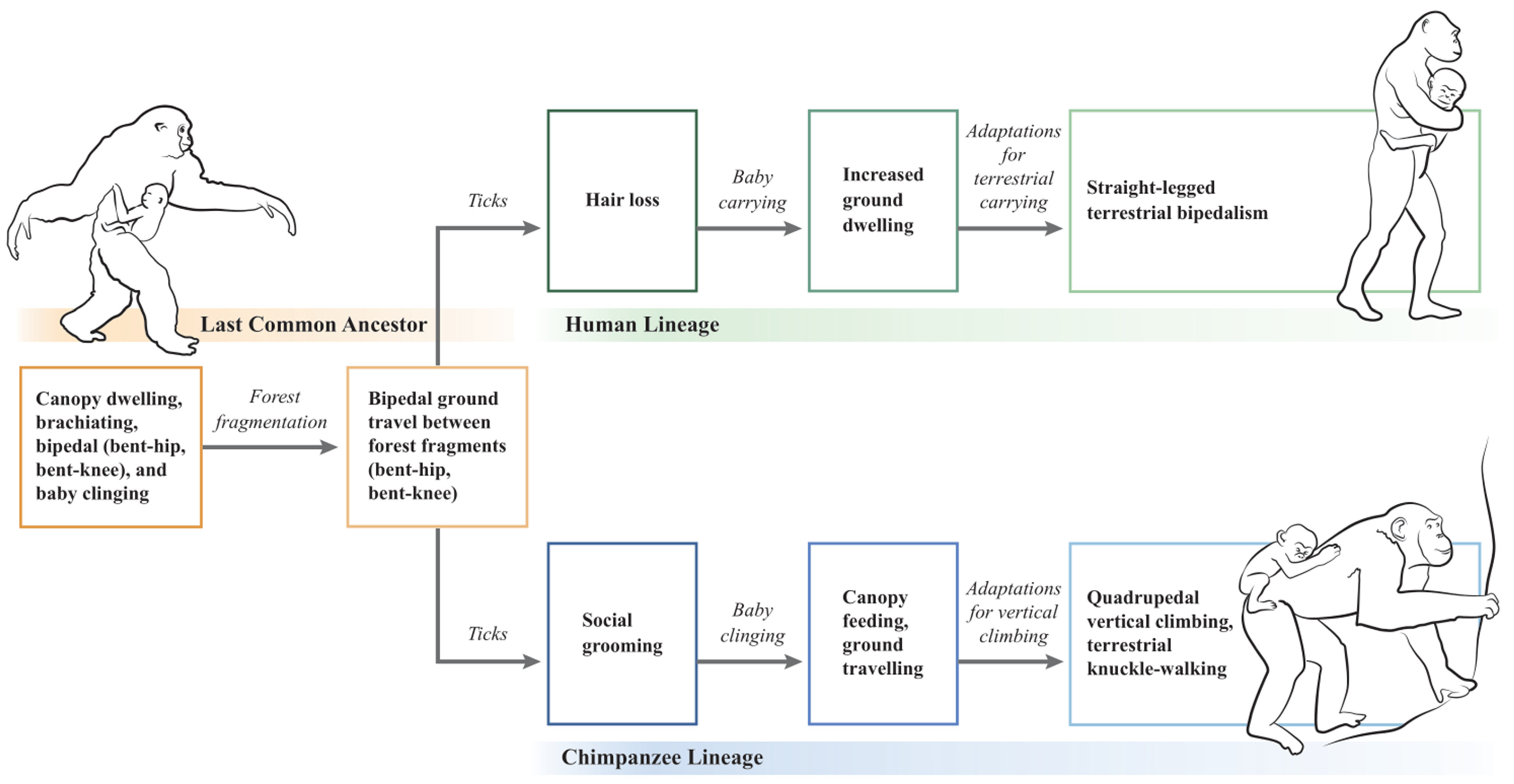
5. A Tick-Based Hypothesis for the Evolutionary Divergence of Humans and Chimpanzees
6. Limitations of the Tick-Based Hypothesis
7. How Can We Test This Hypothesis
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brunet, M.; Guy, F.; Pilbeam, D.; Mackaye, H.T.; Likius, A.; Ahounta, D.; Beauvilain, A.; Blondel, C.; Bocherens, H.; Boisserie, J.R.; et al. A new hominid from the Upper Miocene of Chad, Central Africa. Nature 2002, 418, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Langergraber, K.E.; Prufer, K.; Rowney, C.; Boesch, C.; Crockford, C.; Fawcett, K.; Inoue, E.; Inoue-Muruyama, M.; Mitani, J.C.; Muller, M.N.; et al. Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 15716–15721. [Google Scholar] [CrossRef] [PubMed]
- Michel, L.A.; Peppe, D.J.; Lutz, J.A.; Driese, S.G.; Dunsworth, H.M.; Harcourt-Smith, W.E.H.; Horner, W.H.; Lehmann, T.; Nightingale, S.; McNulty, K.P. Remnants of an ancient forest provide ecological context for Early Miocene fossil apes. Nat. Commun. 2014, 5, 3236. [Google Scholar] [CrossRef] [PubMed]
- Moorjani, P.; Amorim, C.E.G.; Arndt, P.F.; Przeworski, M. Variation in the molecular clock of primates. Proc. Natl. Acad. Sci. USA 2016, 113, 10607–10612. [Google Scholar] [CrossRef] [PubMed]
- Keith, A. Hunterian lectures on man’s posture: Its evolution and disorders: Given at the Royal College of Surgeons of England. Br. Med. J. 1923, 1, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Reno, P.L. Genetic and developmental basis for parallel evolution and its significance for hominoid evolution. Evol. Anthropol. Issues News Rev. 2014, 23, 188–200. [Google Scholar] [CrossRef]
- Thorpe, S.K.S.; Holder, R.L.; Crompton, R.H. Origin of Human Bipedalism As an Adaptation for Locomotion on Flexible Branches. Science 2007, 316, 1328–1331. [Google Scholar] [CrossRef]
- Böhme, M.; Spassov, N.; Fuss, J.; Tröscher, A.; Deane, A.S.; Prieto, J.; Kirscher, U.; Lechner, T.; Begun, D.R. A new Miocene ape and locomotion in the ancestor of great apes and humans. Nature 2019. [Google Scholar] [CrossRef]
- Pilbeam, D.R. Perspectives on the Miocene Hominoidea. In The Primate Fossil Record; Hartwig, W.C., Ed.; Cambridge University Press: Cambridge, UK, 2002; pp. 301–310. [Google Scholar]
- Tuttle, R.H. Apes and Human Evolution; Harvard University Press: Cambridge, MA, USA, 2014; p. 1056. [Google Scholar]
- Potts, R. Environmental hypotheses of hominin evolution. Am. J. Phys. Anthropol. 1998, 107, 93–136. [Google Scholar] [CrossRef]
- Reynolds, S.C.; Wilkinson, D.M.; Marston, C.G.; O’Regan, H.J. The ‘mosaic habitat’ concept in human evolution: Past and present. Trans. R. Soc. S. Afr. 2015, 70, 57–69. [Google Scholar] [CrossRef]
- Cheyne, S.M. Gibbon locomotion research in the field: Problems, possibilities, and benefits for conservation. In Primate locomotion: Linking Field and Laboratory Research; D’Août, K., Vereecke, E.E., Eds.; Springer Science: New York, NY, USA, 2011; pp. 201–213. [Google Scholar]
- Ishida, H. Current thoughts on terrestrialization in African apes and the origin of human bipedalism. In Human Origins and Environmental Backgrounds; Ishida, H., Tuttle, R., Pickford, M., Ogihara, N., Nakatsukasa, M., Eds.; Springer: New York, NY, USA, 2006; pp. 259–266. [Google Scholar]
- Niemitz, C. The evolution of the upright posture and gait—a review and a new synthesis. Die Nat. 2010, 97, 241–263. [Google Scholar] [CrossRef]
- Dávid-Barrett, T.; Dunbar, R.I.M. Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling. J. Hum. Evol. 2016, 94, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, P.E. The loss of functional body hair in man: The influence of thermal environment, body form and bipedality. J. Hum. Evol. 1985, 14, 23–28. [Google Scholar] [CrossRef]
- Rodman, P.S.; McHenry, H.M. Bioenergetics and the origin of hominid bipedalism. Am. J. Phys. Anthropol. 1980, 52, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, N.G.; Chaplin, G. Origin of habitual terrestrial bipedalism in the ancestor of the Hominidae. J. Hum. Evol. 1993, 24, 259–280. [Google Scholar] [CrossRef]
- Hunt, K.D. The evolution of human bipedality: Ecology and functional morphology. J. Hum. Evol. 1994, 26, 183–202. [Google Scholar] [CrossRef]
- Carrier, D.R. The Advantage of Standing Up to Fight and the Evolution of Habitual Bipedalism in Hominins. PLoS ONE 2011, 6, e19630. [Google Scholar] [CrossRef]
- Berecz, B.; Cyrille, M.; Casselbrant, U.; Oleksak, S.; Norholt, H. Carrying human infants—An evolutionary heritage. Infant Behav. Dev. 2020, 60, 101460. [Google Scholar] [CrossRef]
- Iwamoto, M. Bipedalism of Japanese monkeys and carrying models of hominization. In Primate Morphophysiology. Locomotor Analyses and Human Bipedalism, Kondo, S., Ed.; Tokyo University Press: Tokyo, Japan, 1985; pp. 251–260. [Google Scholar]
- Reno, P.L.; McCollum, M.A.; Meindl, R.S.; Lovejoy, C.O. An enlarged postcranial sample confirms Australopithecus afarensis dimorphism was similar to modern humans. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3355. [Google Scholar] [CrossRef]
- Calvin, W. A Brief History of the Mind: From Apes to Intellect and Beyond; Oxford University Press: Oxford, UK, 2005; p. 240. [Google Scholar]
- Sutou, S. Hairless mutation: A driving force of humanization from a human–ape common ancestor by enforcing upright walking while holding a baby with both hands. Genes Cells 2012, 17, 264–272. [Google Scholar] [CrossRef]
- Amaral, L.Q. Early hominoid physical evolution. Hum. Evol. 1989, 4, 33–44. [Google Scholar] [CrossRef]
- Amaral, L.Q. Mechanical analysis of infant carrying in hominoids. Naturwissenschaften 2008, 95, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Hoff, M.P.; Nadler, R.D.; Maple, T.L. Maternal transport and infant motor development in a captive group of lowland gorillas. Primates 1983, 24, 77–85. [Google Scholar] [CrossRef]
- Schultz, A.H. The Life of Primates; Universe Books: New York, NY, USA, 1969; p. 281. [Google Scholar]
- Van Lawick-Goodall, J. The Behaviour of Free-living Chimpanzees in the Gombe Stream Reserve. In Animal Behaviour Monographs; Cullen, J.M., Beer, C.G., Eds.; Ballière, Tindall & Cassell: London, UK, 1968; Volume 1, pp. 161–311. [Google Scholar]
- Rantala, M.J. Human nakedness: Adaptation against ectoparasites? Int. J. Parasitol. 1999, 29, 1987–1989. [Google Scholar] [CrossRef]
- Belt, T. The Naturalist in Nicaragua; J.M. Dent & Sons: London, UK, 1874. [Google Scholar]
- Kupfer, T.R.; Fessler, D.M.T. Ectoparasite defence in humans: Relationships to pathogen avoidance and clinical implications. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170207. [Google Scholar] [CrossRef] [PubMed]
- Pagel, M.; Bodmer, W. A naked ape would have fewer parasites. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, S117–S119. [Google Scholar] [CrossRef]
- Rantala, M.J. Evolution of nakedness in Homo sapiens. J. Zool. 2007, 273, 1–7. [Google Scholar] [CrossRef]
- Korch, G.W. Geographic dissemination of tick-borne zoonoses. In Ecological Dynamics of Tick-Borne Disease; Sonenshine, D.E., Mather, T.N., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 139–197. [Google Scholar]
- Rodriguez, I.A.; Rasoazanabary, E.; Godfrey, L.R. Seasonal variation in the abundance and distribution of ticks that parasitize Microcebus griseorufus at the Bezà Mahafaly Special Reserve, Madagascar. Int. J. Parasitol. Parasites Wildl. 2015, 4, 408–413. [Google Scholar] [CrossRef]
- Fumagalli, M.; Sironi, M.; Pozzoli, U.; Ferrer-Admettla, A.; Pattini, L.; Nielsen, R. Signatures of environmental genetic adaptation pinpoint pathogens as the main selective pressure through human evolution. PLoS Genet. 2011, 7, e1002355. [Google Scholar] [CrossRef]
- Karvonen, A.; Seehausen, O. The role of parasitism in adaptive radiations—when might parasites promote and when might they constrain ecological speciation? Int. J. Ecol. 2012, 2012, 280169. [Google Scholar] [CrossRef]
- Sironi, M.; Cagliani, R.; Forni, D.; Clerici, M. Evolutionary insights into host-pathogen interactions from mammalian sequence data. Nat. Rev. Genet. 2015, 16, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Buckling, A.; Rainey, P.B. The role of parasites in sympatric and allopatric host diversification. Nature 2002, 420, 496–499. [Google Scholar] [CrossRef]
- Kwiatkowski, D.P. How malaria has affected the human genome and what human genetics can teach us about malaria. Am. J. Hum. Genet. 2005, 77, 171–192. [Google Scholar] [CrossRef]
- Nunn, C.L.; Altizer, S.; Sechrest, W.; Jones, K.E.; Barton, R.A.; Gittleman, J.L. Parasites and the evolutionary diversification of primate clades. Am. Nat. 2004, 164, S90–S103. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.; Ko, W.Y.; Davis, A.; Tishkoff, S.A. Impact of natural selection due to malarial disease on human genetic variation. In Primates, Pathogens and Evolution; Brinkworth, J.F., Pechenkina, K., Eds.; Springer: New York, NY, USA, 2013; pp. 161–186. [Google Scholar]
- Lopez, C.; Saravia, C.; Gomez, A.; Hoebeke, J.; Patarroyo, M.A. Mechanisms of genetically-based resistance to malaria. Gene 2010, 467, 1–12. [Google Scholar] [CrossRef]
- Piel, F.B.; Patil, A.P.; Howes, R.E.; Nyangiri, O.A.; Gething, P.W.; Williams, T.N.; Weatherall, D.J.; Hay, S.I. Global distribution of the sickle cell gene and geographical confirmation of the malaria hypothesis. Nat. Commun. 2010, 1, 104. [Google Scholar] [CrossRef] [PubMed]
- Ghiglieri, M.P. The Chimpanzees of Kibale Forest: A Field Study of Ecology and Social Structure; Columbia University Press: New York, NY, USA, 1984. [Google Scholar]
- Nunn, C.L.; Altizer, S. Infectious Diseases in Primates: Behavior, Ecology and Evolution; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Obenchain, F.D.; Galun, R. Physiology of Ticks. Current Themes in Tropical Science; Pergamon Press: Oxford, UK, 1982. [Google Scholar]
- Darwin, C. The Descent of Man and Selection in Relation to Sex, 2nd ed.; Murray: London, UK, 1888. [Google Scholar]
- Perry, G.H. Parasites and human evolution. Evol. Anthropol. Issues News Rev. 2014, 23, 218–228. [Google Scholar] [CrossRef]
- Primack, R.; Corlett, R. Tropical Rain Forests: An Ecological and Biogeographical Comparison; Blackwell Publishing: Malden, MA, USA, 2005. [Google Scholar]
- Marsh, L.K. The nature of fragmentation. In Primates in Fragments: Ecology and Conservation; Marsh, L.K., Ed.; Kluwer Academic: New York, NY, USA, 2003; pp. 1–10. [Google Scholar]
- deMenocal, P.B.; Bloemendal, J. Plio-pleistocene climate variability in subtropical Africa and the paleoenvironment of hominid evolution: A combined data-model approach. In Paleoclimate and Evolution: With Emphasis on Human Origins; Vrba, E.S., Denton, G.H., Partridge, T.C., Burckle, L.H., Eds.; Yale University Press: New Haven, CT, USA, 1995; pp. 262–288. [Google Scholar]
- Maslin, M.A.; Brierley, C.M.; Milner, A.M.; Shultz, S.; Trauth, M.H.; Wilson, K.E. East African climate pulses and early human evolution. Quat. Sci. Rev. 2014, 101, 1–17. [Google Scholar] [CrossRef]
- Wesselman, H.B. Of mice and men: Regional paleoecology and human evolution in the Turkana Basin. In Paleoclimate and Evolution: With Emphasis on Human Origins; Vrba, E.S., Denton, G.H., Partridge, T.C., Burckle, L.H., Eds.; Yale University Press: New Haven, CT, USA, 1995; pp. 356–384. [Google Scholar]
- Coppens, Y. L’évolution des hominidés, de leur locomotion et de leurs environnements. In Origine(s) de la Bipédie Chez les Hominidés; Coppens, Y., Senut, B., Eds.; Centre National de la Recherche Scietifique: Paris, France, 1991; pp. 295–301. [Google Scholar] [CrossRef]
- Pickford, M. What caused the first steps towards the evolution of walkie-talkie primates? In Origine(s) de la Bipédie Chez les Hominidés; Coppens, Y., Senut, B., Eds.; Centre National de la Recherche Scietifique: Paris, France, 1991; pp. 275–293. [Google Scholar] [CrossRef]
- Senut, B. Arboreal Origins of Bipedalism. In Human Origins and Environmental Backgrounds; Ishida, H., Tuttle, R., Pickford, M., Ogihara, N., Nakatsukasa, M., Eds.; Springer: New York, NY, USA, 2006; pp. 199–208. [Google Scholar]
- Zihlman, A.; LaPorte, L. Plates, climates and hominoid evolution. S. Afr. J. Sci. 1983, 79, 96–110. [Google Scholar]
- Martin, R.D. Primates: A definition. In Major Topics in Primate and Human Evolution; Wood, B., Martin, L., Andrews, P., Eds.; Academic Press: London, UK, 1986; pp. 1–31. [Google Scholar]
- Lewin, R.; Foley, R.A. Principles of Human Evolution, 2nd ed.; Blackwell Publishing: Malden, MA, USA, 2004. [Google Scholar]
- Le Fur, S.; Fara, E.; Mackaye, H.T.; Vignaud, P.; Brunet, M. The mammal assemblage of the hominid site TM266 (Late Miocene, Chad Basin): Ecological structure and paleoenvironmental implications. Naturwissenschaften 2009, 96, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Pickford, M. Paleoenvironments, paleoecology, adaptations and the origins of bipedalism in Hominidae. In Human Origins and Environmental Backgrounds; Ishida, H., Tuttle, R., Pickford, M., Ogihara, N., Nakatsukasa, M., Eds.; Springer: New York, NY, USA, 2006; pp. 175–198. [Google Scholar]
- Roche, D.; Ségalen, L.; Senut, B.; Pickford, M. Stable isotope analyses of tooth enamel carbonate of large herbivores from the Tugen Hills deposits: Palaeoenvironmental context of the earliest Kenyan Hominids. Earth Planet. Sci. Lett. 2013, 381, 39–51. [Google Scholar] [CrossRef]
- Senut, B.; Pickford, M.; Gommery, D.; Ségalen, L. Palaeoenvironments and the origin of hominid bipedalism. Hist. Biol. 2018, 30, 284–296. [Google Scholar] [CrossRef]
- WoldeGabriel, G.; Haile-Selassie, Y.; Renne, P.R.; Hart, W.K.; Ambrose, S.H.; Asfaw, B.; Heiken, G.; White, T. Geology and palaeontology of the Late Miocene Middle Awash valley, Afar rift, Ethiopia. Nature 2001, 412, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Goosem, M. Internal fragmentation: The effect of roads, highways, and powerline clearings on movements and mortality of rainforest vertebrates. In Tropical Rain Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R., Jr., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 241–255. [Google Scholar]
- Döbert, T.F.; Webber, B.L.; Barnes, A.D.; Dickinson, K.J.M.; Didham, R.K. Forest fragmentation and biodiversity conservation in human-dominated landscapes In Global Forest Fragmentation; Kettle, C.J., Koh, L.P., Eds.; CAB International: Wallingford, Oxfordshire, UK, 2014; pp. 28–49. [Google Scholar]
- Laurance, W.F. Section III Tropical Forest Faunas. Introduction. In In Tropical Rain Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R.O., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 85–90. [Google Scholar]
- Gascon, C.; Lovejoy, T.E.; Bierregaard, R.O., Jr.; Malcolm, J.R.; Stouffer, P.C.; Vasconcelos, H.L.; Laurance, W.F.; Zimmerman, B.; Tocher, M.; Borges, S. Matrix habitat and species richness in tropical forest remnants. Biol. Conserv. 1999, 91, 223–229. [Google Scholar] [CrossRef]
- Garcia, C.A.; Feintrenie, L. Beyond the mirror: Tropical forest fragmentation and its impact on rural livelihoods. In Global Forest Fragmentation; Kettle, C.J., Koh, L.P., Eds.; CABI: Wallingford, Oxfordshire, UK, 2014; pp. 115–131. [Google Scholar]
- Halos, L.; Bord, S.; Cotté, V.; Gasqui, P.; Abrial, D.; Barnouin, J.; Boulouis, H.J.; Vayssier-Taussat, M.; Vourc’h, G. Ecological factors characterizing the prevalence of bacterial tick-borne pathogens in Ixodes ricinus ticks in pastures and woodlands. Appl. Environ. Microbiol. 2010, 76, 4413–4420. [Google Scholar] [CrossRef]
- Laurance, W.F. Section II Physical Processes and Edge Effects. Introduction: Edge Effects in Fragmented Forests. In Tropical Rain Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R.O., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 29–31. [Google Scholar]
- Boyard, C.; Vourc’h, G.; Barnouin, J. The relationships between Ixodes ricinus and small mammal species at the woodland–pasture interface. Exp. Appl. Acarol. 2008, 44, 61–76. [Google Scholar] [CrossRef]
- Bender, D.J.; Contreras, T.A.; Fahrig, L. Habitat loss and population decline: A meta-analysis of the patch size effect. Ecology 1998, 79, 517–533. [Google Scholar] [CrossRef]
- Cheyne, S.M.; Thompson, C.J.H.; Chivers, D.J. Travel adaptations of Bornean Agile Gibbons Hylobates albibarbis (Primates: Hylobatidae) in a degraded secondary forest, Indonesia. J. Threat. Taxa 2013, 5, 3963–3968. [Google Scholar] [CrossRef]
- Das, J.; Biswas, J.; Bhattacherjee, P.C.; Rao, S.S. Canopy Bridges: An Effective Conservation Tactic for Supporting Gibbon Populations in Forest Fragments. In The Gibbons: New Perspectives on Small Ape Socioecology and Population Biology; Lappan, S., Whittaker, D.J., Eds.; Springer: New York, NY, USA, 2009; pp. 467–496. [Google Scholar]
- Sati, J.P.; Alfred, J.R.B. Locomotion and posture in Hoolock gibbon. Ann. For. 2002, 10, 298–306. [Google Scholar] [CrossRef]
- Vereecke, E.E.; D’Aout, K.; Aerts, P. Locomotor versatility in the white-handed gibbon (Hylobates lar): A spatiotemporal analysis of the bipedal, tripedal, and quadrupedal gaits. J. Hum. Evol. 2006, 50, 552–567. [Google Scholar] [CrossRef]
- Chapman, C.A.; Speirs, M.L.; Gillespie, T.R.; Holland, T.; Austad, K.M. Life on the edge: Gastrointestinal parasites from the forest edge and interior primate groups. Am. J. Primatol. 2006, 68, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Fiennes, R. Zoonoses of Primates; Cornell University Press: Ithaca, NY, USA, 1967. [Google Scholar]
- Malcolm, J.R. Edge Effects in Central Amazonian Forest Fragments. Ecology 1994, 75, 2438–2445. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- García-Marmolejo, G.; Chapa-Vargas, L.; Weber, M.; Huber-Sannwald, E. Landscape composition influences abundance patterns and habitat use of three ungulate species in fragmented secondary deciduous tropical forests, Mexico. Glob. Ecol. Conserv. 2015, 3, 744–755. [Google Scholar] [CrossRef]
- Malcolm, J.R. Biomass and diversity of small mammals in Amazonian forest fragments. In Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R.O., Jr., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 207–221. [Google Scholar]
- Allan, B.F.; Keesing, F.; Ostfeld, R.S. Effect of forest fragmentation on lyme disease risk. Conserv. Biol. 2003, 17, 267–272. [Google Scholar] [CrossRef]
- Levi, T.; Kilpatrick, A.M.; Mangel, M.; Wilmers, C.C. Deer, predators, and the emergence of Lyme disease. Proc. Natl. Acad. Sci. USA 2012, 109, 10942–10947. [Google Scholar] [CrossRef]
- Terbough, J.; Lopez, L.; Nuñez, P.; Rao, M.; Shahabuddin, G.; Orihuela, G.; Riveros, M.; Ascanio, R.; Adler, G.H.; Lambert, T.D.; et al. Ecologic meltdown in predator-free forest fragments. Science 2001, 294, 1923–1926. [Google Scholar] [CrossRef]
- Randolph, S.E. Tick ecology: Processes and patterns behind the epidemiological risk posed by ixodid ticks as vectors. Parasitology 2004, 129, S37–S65. [Google Scholar] [CrossRef]
- Raši, T.; Majláth, I.; Bogdziewicz, M.; Dudek, K.; Majláthová, V.; Włodarek, J.; Almáši, M.; Vargová, B.; Tryjanowski, P. Tick distribution along animal tracks: Implication for preventative medicine. Ann. Agric. Environ. Med. 2018, 25, 360–363. [Google Scholar] [CrossRef]
- Broekhuis, F.; O’Meara, R.H.; O’Meara, S.; Barton, M.; Harrell, C.; Western, G.; Elliot, N.B. An assessment of mammals in Naimina Enkiyio Forest, Kenya. Afr. J. Ecol. 2018, 56, 755–758. [Google Scholar] [CrossRef]
- Sonenshine, D.E. The biology of tick vectors of human disease. In Tick-Borne Diseases of Humans; Goodman, J.L., Dennis, D.T., Sonenshine, D.E., Eds.; ASM Press: Washington, DC, USA, 2005; pp. 12–36. [Google Scholar]
- Sonenshine, D.E.; Roe, R.M. Overview: Ticks, people and animals. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2013; pp. 3–16. [Google Scholar]
- Dennis, D.T.; Piesman, J.F. Overview of tick-borne infections of humans. In Tick-Borne Diseases of Humans; Goodman, J.L., Dennis, D.T., Sonenshine, D.E., Eds.; American Society for Microbiology: Washington, DC, USA, 2005; pp. 3–11. [Google Scholar]
- Apanaskevich, D.A.; Oliver, J.H., Jr. Life cycles and natural history of ticks. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, New York, USA, 2013; Volume 1, pp. 59–73. [Google Scholar]
- Randolph, S.E. The impact of tick ecology on pathogen transmission dynamics. In Ticks: Biology, Disease and Control; Bowman, A.S., Nuttall, P., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 40–72. [Google Scholar]
- Sonenshine, D.E. Introduction. In Ecological Dynamics of Tick-Borne Disease; Sonenshine, D.E., Mather, T.N., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 3–19. [Google Scholar]
- Mejlon, H.A.; Jaenson, T.G.T. Questing behaviour of Ixodes ricinus ticks (Acari: Ixodidae). Exp. Appl. Acarol. 1997, 21, 747–754. [Google Scholar] [CrossRef]
- Oorebeek, M.; Sharrad, R.; Kleindorfer, S. What attracts larval Ixodes hirsti (Acari: Ixodidae) to their host? Parasitol. Res. 2009, 104, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Mather, T.N.; Ginsberg, H.S. Vector-host-pathogen relationships: Transmission dynamics of tick-borne infections. In Ecological Dynamics of Tick-Borne Disease; Sonenshine, D.E., Mather, T.N., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 68–90. [Google Scholar]
- Levin, M.L.; Ford, S.L.; Hartzer, K.; Krapiunaya, L.; Stanley, H.; Snellgrove, A.N. Minimal Duration of Tick Attachment Sufficient for Transmission of Infectious Rickettsia rickettsii (Rickettsiales: Rickettsiaceae) by Its Primary Vector Dermacentor variabilis (Acari: Ixodidae): Duration of Rickettsial Reactivation in the Vector Revisited. J. Med. Entomol. 2019, 57, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Falco, R.C.; Fish, D.; Piesman, J. Duration of tick bites in a Lyme disease-endemic area. Am. J. Epidemiol. 1996, 143, 187–192. [Google Scholar] [CrossRef]
- Fournier, P.E.; Raoult, D. Mediterranean spotted fever and other tick-borne rickettsioses. In Tick-Borne Diseases of Humans; Goodman, J.L., Dennis, D.T., Sonenshine, D.E., Eds.; American Society for Microbiology: Washington, DC, USA, 2005; pp. 302–327. [Google Scholar]
- Piesman, J.; Mather, T.N.; Sinsky, R.J.; Spielman, A. Duration of tick attachment and Borrelia burgdorferi transmission. J. Clin. Microbiol. 1987, 25, 557–558. [Google Scholar] [CrossRef]
- Katavolos, P.; Armstrong, P.M.; Dawson, J.E.; Telford, S.R., III. Duration of Tick Attachment Required for Transmission of Granulocytic Ehrlichiosis. J. Infect. Dis. 1998, 177, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.J. Lyme borreliosis: A review of data on transmission time after tick attachment. Int. J. Gen. Med. 2015, 8, 1–8. [Google Scholar] [CrossRef]
- Akinyi, M.Y.; Tung, J.; Jeneby, M.; Patel, N.B.; Altmann, J.; Alberts, S.C. Role of grooming in reducing tick load in wild baboons (Papio cynocephalus). Anim. Behav. 2013, 85, 559–568. [Google Scholar] [CrossRef]
- Bourliere, F. Mammals, large and small: The ecological implications of size. In Small Mammals: Their Productiivity and Population Dynamics; Petrusewitz, K., Ryszkowski, L., Eds.; Cambridge University Press: Cambridge, London, UK, 1975; pp. 1–8. [Google Scholar]
- Robinson, S.J.; Neitzel, D.F.; Moen, R.A.; Craft, M.E.; Hamilton, K.E.; Johnson, L.B.; Mulla, D.J.; Munderloh, U.G.; Redig, P.T.; Smith, K.E.; et al. Disease Risk in a Dynamic Environment: The Spread of Tick-Borne Pathogens in Minnesota, USA. EcoHealth 2015, 12, 152–163. [Google Scholar] [CrossRef]
- Daniel, M.; Dusbábek, F. Micrometeorological and Microhabitat Factors Affecting Maintenance and Dissemination of Tick-borne Diseases in the Environment. In Ecological Dynamics of Tick-Borne Disease; Sonenshine, D., Mather, T., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 91–138. [Google Scholar]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Stein, K.J.; Waterman, M.; Waldon, J.L. The effects of vegetation density and habitat disturbance on the spatial distribution of ixodid ticks (Acari: Ixodidae). Geospat Health 2008, 2, 241–252. [Google Scholar] [CrossRef]
- Estrada-Pena, A. The relationships between habitat topology, critical scales of connectivity and tick abundance Ixodes ricinus in a heterogeneous landscape in northern Spain. Ecography 2003, 26, 661–671. [Google Scholar] [CrossRef]
- Vanwambeke, S.O.; Sumilo, D.; Boremane, A.; Lambin, E.F.; Randolph, S.E. Landscape predictors of tick-borne encephalitis in Latvia: Land cover, land use, and land ownership. Vector Borne Zoonotic Dis. 2010, 10, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.G. The relevance of forest fragmentation on the incidence of human babesiosis: Investigating the landscape epidemiology of an emerging tick-borne disease. Vector Borne Zoonotic Dis. 2013, 13, 250–255. [Google Scholar] [CrossRef]
- Wimberly, M.C.; Yabsley, M.J.; Baer, A.D.; Dugan, V.G.; Davidson, W.R. Spacial heterogeneity of climate and land-cover constraints on distributions of tick-borne pathogens. Glob. Ecol. Biogeogr. 2008, 17, 189–202. [Google Scholar] [CrossRef]
- Brownstein, J.S.; Skelly, D.K.; Holdford, T.R.; Fish, D. Forest fragmentation predicts local scale hereogeneity of Lyme disease risk. Oecologia 2005, 146, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Nupp, T.E.; Swihart, R.K. Effect of forest patch area on population attributes of white-footed mice (Peromyscus leucopus) in fragmented habitats. Can. J. Zool. 1996, 74, 467–472. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Canham, C.D.; Oggenfuss, K.; Winchcombe, R.J.; Keesing, F. Climate, deer, rodents, and acorns as determinants of variation in lyme-disease risk. PLoS Biol. 2006, 4, e145. [Google Scholar] [CrossRef] [PubMed]
- Nupp, T.E.; Swihart, R.K. Effects of forest fragmentation on population attributes of white-footed mice and eastern chipmunks. J. Mammal. 1998, 79, 1234–1243. [Google Scholar] [CrossRef]
- Wilson, M.L. Correlation between abundance of deer and that of the deer tick, Ixodes dammini (Acari: Ixodidae). Ann. Entomol. Soc. Am. 1985, 78, 172–176. [Google Scholar] [CrossRef]
- Horak, I.G.; Fourie, L.J.; Heyne, H.; Walker, J.B.; Needham, G.R. Ixodid Ticks Feeding on Humans in South Africa: With Notes on Preferred Hosts, Geographic Distribution, Seasonal Occurrence and Transmission of Pathogens. Exp. Appl. Acarol. 2002, 27, 113–136. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Vatansever, Z.; Gargili, A.; Ergönul, O. The trend towards habitat fragmentation is the key factor driving the spread of Crimean-Congo haemorrhagic fever. Epidemiol. Infect. 2010, 138, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.E. Early hominid evolution and ecological change through the African Plio-Pleistocene. J. Hum. Evol. 1997, 32, 289–322. [Google Scholar] [CrossRef]
- Beinart, W.; Brown, K. African Local Knowledge & Livestock Health: Diseases & Treatments in South Africa; James Currey: Melton, UK, 2013; p. 304. [Google Scholar]
- Veríssimo, C.J.; Vasques, F.; Duarte, K.M.; Paulino, V.T.; Ambrósio, L.A. Management and control of parasites on dairy farms in northwestern region of São Paulo state. Rev. Bras. Parasitol. Vet. 2016, 25, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Poiner, G. Spirochete-like cells in a Dominican amber Ambylomma tick (Arachnida: Ixodidae). Hist. Biol. 2015, 27, 565–570. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; de la Fuente, J. The fossil record and the origin of ticks revisited. Exp. Appl. Acarol. 2018, 75, 255–261. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J. The fossil record and the origin of ticks (Acari: Parasitiformes: Ixodida). Exp. Appl. Acarol. 2003, 29, 331–344. [Google Scholar] [CrossRef]
- Ndip, L.M.; Fokam, E.B.; Bouyer, D.H.; Ndip, R.N.; Titanji, V.P.K.; Walker, D.H.; McBride, J.W. Detection of Rickettsia africae in patients and ticks along the coastal region of Cameroon. Am. J. Trop. Med. Hyg. 2004, 71, 363–366. [Google Scholar] [CrossRef]
- Shayan, S.; Bokaean, M.; Shahrivar, M.R.; Chinikar, S. Crimean-Congo Hemorrhagic Fever. Lab. Med. 2015, 46, 180–189. [Google Scholar] [CrossRef]
- Guglielimone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G. The Hard Ticks of the World: (Acari: Ixodida: Ixodidae); Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Brown, R.N.; Lane, R.S.; Dennis, D.T. Geographic distributions of tick-borne diseases and their vectors. In Tick-Borne Diseases of Humans; Goodman, J.L., Dennis, D.T., Sonenshine, D.E., Eds.; American Society for Microbiology: Washington, DC, USA, 2005; pp. 363–391. [Google Scholar]
- Bishop, R.; Musoke, A.; Skilton, R.; Morzaria, S.; Gardner, M.; Nene, V. Theileia: Life cycle stages associated with the ixodid tick vector. In Ticks: Biology, Disease and Control; Bowman, A.S., Nuttall, P., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 308–324. [Google Scholar]
- Maclatchy, L.M.; DeSilva, J.M.; Sanders, W.J.; Wood, B. Hominini. In Cenozoic Mammals of Africa; Werdelin, L., Sanders, W.J., Eds.; University of California Press: Berkeley, CA, USA, 2010; pp. 471–540. [Google Scholar]
- Monadjem, A.; Taylor, P.J.; Denys, C.; Cotterill, F.P.D. Rodents of Sub-Saharan Africa: A Biogeographic and Taxonomic Synthesis; De Gruyter: Berlin, Germany, 2015. [Google Scholar]
- Matthysse, J.G.; Colbo, M.H. The Ixodid Ticks of Uganda; Entomological Society of America: Annapolis, MD, USA, 1987. [Google Scholar]
- Estrada-Pena, A.; de la Fuente, J. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 2014, 108, 104–128. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Sprong, H.; Cabezas-Cruz, A.; de la Fuente, J.; Ramo, A.; Coipan, E.C. Nested coevolutionary networks shape the ecological relationships of ticks, hosts, and the Lyme disease bacteria of the Borrelia burgdorferi (s.l.) complex. Parasit. Vectors 2016, 9, 517. [Google Scholar] [CrossRef]
- Hoogstraal, H.; Kim, K.C. Tick and mammal coevolution, with emphasis on Haemaphysalis. In Coevolution of Parasitic Arthropods and Mammals; Kim, K.C., Ed.; John Wiley & Sons: Chichester, West Sussex, UK, 1985; pp. 505–568. [Google Scholar]
- Voordouw, M.J.; Lachish, S.; Dolan, M.C. The Lyme Disease Pathogen Has No Effect on the Survival of Its Rodent Reservoir Host. PLoS ONE 2015, 10, e0118265. [Google Scholar] [CrossRef] [PubMed]
- McKnight, D.T.; Schwarzkopf, L.; Alford, R.A.; Bower, D.S.; Zenger, K.R. Effects of emerging infectious diseases on host population genetics: A review. Conserv. Genet. 2017, 18, 1235–1245. [Google Scholar] [CrossRef]
- Galili, U. Natural anti-carbohydrate antibodies contributing to evolutionary survival of primates in viral epidemics? Glycobiology 2016, 26, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.A. History of Bovine Theileriosis in Southern Africa. In The Epidemiology of Theileriosis in Africa; Norval, R.A.I., Perry, B.D., Young, A.S., Eds.; Academic Press: London, UK, 1992; pp. 1–39. [Google Scholar]
- Ali, M.; de Castro, J.J. Host resistance to ticks (Acari: Ixodidae) in different breeds of cattle at Bako, Ethiopia. Trop. Anim. Health Prod. 1993, 25, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Spickett, A.M.; De Klerk, D.; Enslin, C.B.; Scholtz, M.M. Resistance of Nguni, Bonsmara and Hereford cattle to ticks in a Bushveld region of South Africa. Onderstepoort J. Vet. Res. 1989, 56, 245–250. [Google Scholar]
- Zhang, X.; Rayner, J.G.; Blaxter, M.; Bailey, N.W. Rapid parallel adaptation despite gene flow in silent crickets. Nat. Commun. 2021, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Perspectives on hair evolution based on some comparative studies on vertebrate cornification. J. Exp. Zool. Part B Mol. Dev. Evol. 2012, 318, 325–343. [Google Scholar] [CrossRef]
- Allen, J.M.; Worman, C.O.; Light, J.E.; Reed, D.L. Parasitic Lice Help to Fill in the Gaps of Early Hominid History. In Primates, Pathogens and Evolution; Brinkworth, J.F., Pechenkina, K., Eds.; Springer: New York, NY, USA, 2013; pp. 161–186. [Google Scholar]
- Kamberov, Y.G.; Guhan, S.M.; DeMarchis, A.; Jiang, J.; Wright, S.S.; Morgan, B.A.; Sabeti, P.C.; Tabin, C.J.; Lieberman, D.E. Comparative evidence for the independent evolution of hair and sweat gland traits in primates. J. Hum. Evol. 2018, 125, 99–105. [Google Scholar] [CrossRef]
- Szabo, G. The Regional Anatomy of the Human Integument with Special Reference to the Distribution of Hair Follicles, Sweat Glands and Melanocytes. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1967, 252, 447–485. [Google Scholar]
- Hardy, A. Was man more aquatic in the past? New Sci. 1960, 7, 642–645. [Google Scholar]
- Leo, N.P. Cryptic species and biodiversity of lice from primates. In Primate Parasite Ecolology; Huffman, M.A., Chapman, C.A., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 251–269. [Google Scholar]
- Voigt, D.; Gorb, S. Functional morphology of tarsal adhesive pads and attachment ability in ticks Ixodes ricinus (Arachnida, Acari, Ixodidae). J. Exp. Biol. 2017, 220, 1984. [Google Scholar] [CrossRef]
- Keene, S.A.; Rassman, W.R.; Harris, J.A. Determining Safe Excision Limits in FUE: Factors That Affect, and a Simple Way to Maintain, Aesthetic Donor Density. Int. Soc. Hair Restor. Surg. 2018, 28, 1. [Google Scholar] [CrossRef]
- Rassman, W.; Pak, J.; Kim, J. Combining Follicular Unit Extraction and Scalp Micropigmentation for the Cosmetic Treatment of Alopecias. Plast. Reconstr. Surg. Glob. Open 2017, 5, e1420. [Google Scholar] [CrossRef] [PubMed]
- Beeler, T.; Bickel, B.; Noris, G.; Beardsley, P.; Marschner, S.; Sumner, R.W.; Gross, M. Coupled 3D reconstruction of sparse facial hair and skin. ACM Trans. Graph. 2012, 31, 1–10. [Google Scholar] [CrossRef]
- Lachiewicz, A.M.; Berwick, M.; Wiggins, C.L.; Thomas, N.E. Survival differences between patients with scalp or neck melanoma and those with melanoma of other sites in the Surveillance, Epidemiology, and End Results (SEER) program. Arch. Dermatol. 2008, 144, 515–521. [Google Scholar] [CrossRef]
- Sebastian, S. Does preoperative scalp shaving result in fewer postoperative wound infections when compared with no scalp shaving? A systematic review. J. Neurosci. Nurs. 2012, 44, 149–156. [Google Scholar] [CrossRef]
- Walker, A.R.; Bouattour, A.; Camicas, J.L.; Estrada-Peña, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; The University of Edinburgh: Edinburgh, UK, 2003; p. 221. [Google Scholar]
- Stafford, K.C. Tick Management Handbook; The Connecticut Agricultural Experiment Station: New Haven, CT, USA, 2007. [Google Scholar]
- Barbedo, J.G.; Gomes, C.C.; Cardoso, F.F.; Domingues, R.; Ramos, J.V.; McManus, C.M. The use of infrared images to detect ticks in cattle and proposal of an algorithm for quantifying the infestation. Vet. Parasitol. 2017, 235, 106–112. [Google Scholar] [CrossRef]
- Cortivo, P.D.; Dias, E.; Barcellos, J.O.J.; Peripolli, V.; Costa, J.B.G.; Dallago, B.S.L.; McManus, C.M. Use of thermographic images to detect external parasite load in cattle. Comput. Electron. Agric. 2016, 127, 413–417. [Google Scholar] [CrossRef]
- Felz, M.W.; Smith, C.D.; Swift, T.R. A six-year-old girl with tick paralysis. N. Engl. J. Med. 2000, 342, 90–94. [Google Scholar] [CrossRef]
- Jablonski, N.G.; Chaplin, G. Human skin pigmentation as an adaptation to UV radiation. Proc. Natl. Acad. Sci. USA 2010, 107, 8962–8968. [Google Scholar] [CrossRef]
- Toups, M.A.; Kitchen, A.; Light, J.E.; Reed, D.L. Origin of Clothing Lice Indicates Early Clothing Use by Anatomically Modern Humans in Africa. Mol. Biol. Evol. 2011, 28, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Dean, I.; Siva-Jothy, M.T. Human fine body hair enhances ectoparasite detection. Biol. Lett. 2012, 8, 358–361. [Google Scholar] [CrossRef]
- Rogers, A.R.; Iltis, D.; Wooding, S. Genetic variation at the MC1R locus and the time since loss of human body hair. Curr. Anthropol. 2004, 45, 105–124. [Google Scholar] [CrossRef]
- Dror, Y.; Hopp, M. Hair for brain trade-off, a metabolic bypass for encephalization. Springerplus 2014, 3, 562. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Durden, L.A.; Musser, G.G. The sucking lice (Insecta, Anoplura) of the world: A taxonomic checklist with records of mammalian hosts and geographical distributions. Bull. Am. Mus. Nat. Hist. 1994, 218, 1–90. [Google Scholar]
- Reed, D.L.; Light, J.E.; Allen, J.M.; Kirchman, J.J. Pair of lice lost or parasites regained: The evolutionary history of anthropoid primate lice. BMC Biol. 2007, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Sandel, A.A. Brief communication: Hair density and body mass in mammals and the evolution of human hairlessness. Am. J. Phys. Anthropol. 2013, 152, 145–150. [Google Scholar] [CrossRef]
- Cohen, A.; Campisano, C.; Arrowsmith, R.; Asrat, A.; Behrensmeyer, A.K.; Deino, A.; Feibel, C.; Hill, A.; Johnson, R.; Kingston, J.; et al. The hominin sites and paleolakes drilling project: Inferring the environmental context of human evolution from eastern African rift lake deposits. Sci. Drill. 2016, 21, 1–16. [Google Scholar] [CrossRef]
- Thiery, W.; Davin, E.L.; Panitz, H.; Demuzere, M.; Lhermitte, S.; van Lipzig, N. The impact of the African great lakes on the regional climate. J. Clim. 2015, 28, 4061–4085. [Google Scholar] [CrossRef]
- Cedar Lake Ventures. Weather Spark: The Typical Weather Anywhere on Earth. Available online: https://weatherspark.com/ (accessed on 11 August 2020).
- Fruth, B.; Hohmann, G. Nest building behavior in the great apes: The great leap forward? In Great Ape Societies; McGrew, W.C., Marchant, L.F., Nishida, T., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 225–239. [Google Scholar]
- Anderson, J.R. Sleep-related behavioural adaptations in free-ranging anthropoid primates. Sleep Med. Rev. 2000, 4, 355–373. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, M.; Motesharrei, S.; Mu, Q.; Kalnay, E.; Li, S. Local cooling and warming effects of forests based on satellite observations. Nat. Commun. 2015, 6, 6603. [Google Scholar] [CrossRef] [PubMed]
- DeSilva, J.M. A shift toward birthing relatively large infants early in human evolution. Proc. Natl. Acad. Sci. USA 2011, 108, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Pawŀwski, B. Why are human newborns so big and fat? Hum. Evol. 1998, 13, 65–72. [Google Scholar] [CrossRef]
- Kuzawa, C.W. Adipose tissue in human infancy and childhood: An evolutionary perspective. Am. J. Phys. Anthropol. 1998, 107, 177–209. [Google Scholar] [CrossRef]
- Dattani, M.T.; Gevers, E.F. Endocrinology of fetal development. In Williams Textbook of Endocrinology; Melmed, S., Polonsky, K.S., Larsen, P.R., Kronenberg, H.M., Eds.; Elsevier: Philadelphia, PA, USA, 2017; pp. 849–892. [Google Scholar]
- Rowlatt, U.; Mrosovsky, N.; English, A. A comparative survey of brown fat in the neck and axilla of mammals at birth. Biol. Neonate 1971, 17, 53–83. [Google Scholar] [CrossRef] [PubMed]
- Correia, H.R.; Balseiro, S.C.; Correia, E.R.; Mota, P.G.; de Areia, M.L. Why are human newborns so fat? Relationship between fatness and brain size at birth. Am. J. Hum. Biol. 2004, 16, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Crawford, M.A. Survival of the fattest: Fat babies were the key to evolution of the large human brain. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 136, 17–26. [Google Scholar] [CrossRef]
- Blount, Z.D.; Lenski, R.E.; Losos, J.B. Contingency and determinism in evolution: Replaying life’s tape. Science 2018, 362, eaam5979. [Google Scholar] [CrossRef]
- Karasuyama, H.; Miyake, K.; Yoshikawa, S. Immunobiology of Acquired Resistance to Ticks. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Mooring, M.S.; Blumstein, D.T.; Stoner, C.J. The evolution of parasite-defence grooming in ungulates. Biol. J. Linn. Soc. 2004, 81, 17–37. [Google Scholar] [CrossRef]
- Osuna-Mascaró, A.J.; Ortiz, C.; Stolz, C.; Musgrave, S.; Sanz, C.M.; Morgan, D.B.; Fragaszy, D.M. Dexterity and technique in termite fishing by chimpanzees (Pan troglodytes troglodytes) in the Goualougo Triangle, Republic of Congo. Am. J. Primatol. 2021, 83, e23215. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T. Form Perception and Visual Acuity in a Chimpanzee. Folia Primatol. 1990, 55, 24–32. [Google Scholar] [CrossRef]
- Bettridge, C.; Dunbar, R.I.M. Predation as a determinant of minimum group size in baboons. Folia Primatol. 2012, 83, 332–352. [Google Scholar] [CrossRef]
- Reichard, U.; Sommer, V. Grooming Site Preferences in Wild White-handed Gibbons (Hylobates lar). Primates 1994, 35, 369–374. [Google Scholar] [CrossRef]
- Terry, R.L. Primate grooming as a tension reduction mechanism. J. Psychol. Interdiscip. Appl. 1970, 76, 129–136. [Google Scholar] [CrossRef]
- Barton, R.A. Grooming site preferences in primates and their functional implications. Int. J. Primatol. 1985, 6, 519–532. [Google Scholar] [CrossRef]
- Goodall, J. The Chimpanzees of Gombe: Patterns of Behavior; The Belknap Press of Harvard University: Cambridge, MA, USA, 1986. [Google Scholar]
- Hutchins, M.; Barash, D.P. Grooming in primates: Implications for its utilitarian function. Primates 1976, 17, 145–150. [Google Scholar] [CrossRef]
- Durden, L.A.; Lloyd, J.E. Lice (Phthiraptera). In Medical and Veterinary Entomology; Mullen, G.R., Durden, L.A., Eds.; Academic Press: London, UK, 2009; pp. 59–82. [Google Scholar]
- Nicholson, W.L.; Sonenshine, D.E.; Lane, R.S.; Uilenberg, G. Ticks (Ixodida). In Medical and Veterinary Entomology; Mullen, G.R., Durden, L.A., Eds.; Academic Press: London, UK, 2009; pp. 493–542. [Google Scholar]
- Durden, L.A.; Musser, G.G. The mammalian hosts of the sucking lice (Anoplura) of the world: A host-parasite list. Bull. Soc. Vector Ecol. 1994, 19, 130–168. [Google Scholar]
- Kuhn, H.J. Parasites and the phylogeny of the catarrhine primates. In Taxonomy and Phylogeny of Old World Primates with References to the Origin of Man; Chiarelli, B., Ed. Rosenberg & Sellier: Torino, Italy, 1968; pp. 187–195. [Google Scholar]
- Freeland, W.J. Functional aspects of primate grooming. Ohio Acad. Sci. 1981, 81, 173–177. [Google Scholar]
- Brain, C.; Bohrmann, R. Tick infestation of baboons (Papio ursinus) in the Namib Desert. J. Wildl. Dis. 1992, 28, 188–191. [Google Scholar] [CrossRef]
- Young, P.R.; Ng, L.F.P.; Hall, R.A.; Smith, D.W.; Johansen, C.A. 14—Arbovirus Infections. In Manson’s Tropical Infectious Diseases, 23rd ed.; Farrar, J., Hotez, P.J., Junghanss, T., Kang, G., Lalloo, D., White, N.J., Eds.; W.B. Saunders: London, UK, 2014; pp. 129–161.e123. [Google Scholar] [CrossRef]
- Chakraborty, S.; Andrade, F.C.D.; Ghosh, S.; Uelmen, J.; Ruiz, M.O. Historical Expansion of Kyasanur Forest Disease in India From 1957 to 2017: A Retrospective Analysis. GeoHealth 2019, 3, 44–55. [Google Scholar] [CrossRef] [PubMed]
- McCoy, K.D.; Léger, E.; Dietrich, M. Host specialization in ticks and transmission of tick-borne diseases: A review. Front. Cell. Infect. Microbiol. 2013, 3, 57. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Harvey, P.H. Primate ecology and social organization. J. Zool. 1977, 183, 1–39. [Google Scholar] [CrossRef]
- Spruijt, B.M.; van Hooff, J.A.; Gispen, W.H. Ethology and neurobiology of grooming behavior. Physiol. Rev. 1992, 72, 825–852. [Google Scholar] [CrossRef] [PubMed]
- Ruch, T.C. Diseases of the skin. In Diseases of Laboratory Primates; Ruch, T.C., Ed.; Saunders: Philadelphia, PA, USA, 1959; pp. 501–528. [Google Scholar]
- Wrangham, R.W. Feeding behaviour of chimpanzees in the Gombe National Park, Tanzania. In Primate Ecology; Clutton-Brock, T.H., Ed.; Academic Press: London, UK, 1977; pp. 503–538. [Google Scholar]
- Zamma, K.; Nakamura, M. Grooming: Its hygienic and social aspects. In Mahale Chimpanzees: 50 Years of Research; Nakamura, M., Hosaka, K., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 472–484. [Google Scholar]
- Hamer, S.A.; Bernard, A.B.; Donovan, R.M.; Hartel, J.A.; Wrangham, R.W.; Otali, E.; Goldberg, T.L. Coincident tick infestations in the nostrils of wild chimpanzees and a human in Uganda. Am. J. Trop. Med. Hyg. 2013, 89, 924–927. [Google Scholar] [CrossRef]
- Dunbar, R.I.M. Functional Significance of Social Grooming in Primates. Folia Primatol. 1991, 57, 121–131. [Google Scholar] [CrossRef]
- Clark, R.P.; Hu, L.T. Prevention of Lyme Disease and Other Tick-Borne Infections. Infect. Dis. Clin. N. Am. 2008, 22, 381–396. [Google Scholar] [CrossRef]
- Biggs, H.M.; Behravesh, C.B.; Bradley, K.K.; Dahlgren, F.S.; Drexler, N.A.; Dumler, J.S.; Folk, S.M.; Kato, C.Y.; Lash, R.R.; Levin, M.L.; et al. Diagnosis and management of tickborne rickettsial diseases: Rocky mountain spotted fever and other spotted fever group rickettsioses, ehrlichioses, and anaplasmosis—United States. Mmwr Recomm. Rep. 2016, 65, 1–44. [Google Scholar] [CrossRef]
- Jones, H.; Pekins, P.; Kantar, L.; Sidor, I.; Ellingwood, D.; Lichtenwalner, A.; O’Neal, M. Mortality assessment of moose (Alces alces) calves during successive years of winter tick (Dermacentor albipictus) epizootics in New Hampshire and Maine (USA). Can. J. Zool. 2018, 97, 22–30. [Google Scholar] [CrossRef]
- Hart, B.L. Behavioural defense against parasites: Interaction with parasite invasiveness. Parasitology 1994, 109, S139–S151. [Google Scholar] [CrossRef]
- Lehmann, T. Ectoparasites: Direct impact on host fitness. Parasitol. Today 1993, 9, 8–13. [Google Scholar] [CrossRef]
- Grueter, C.C.; Bissonnette, A.; Isler, K.; van Schaik, C.P. Grooming and group cohesion in primates: Implications for the evolution of language. Evol. Hum. Behav. 2013, 34, 61–68. [Google Scholar] [CrossRef]
- Jaeggi, A.V.; Kramer, K.L.; Hames, R.; Kiely, E.J.; Gomes, C.; Kaplan, H.; Gurven, M. Human grooming in comparative perspective: People in six small-scale societies groom less but socialize just as much as expected for a typical primate. Am. J. Phys. Anthropol. 2017, 162, 810–816. [Google Scholar] [CrossRef]
- Ginath Yuh, Y.; N’Goran, P.K.; Dongmo, Z.N.; Tracz, W.; Tangwa, E.; Agunbiade, M.; Kühl, H.S.; Sop, T.; Fotang, C. Mapping suitable great ape habitat in and around the Lobéké National Park, South-East Cameroon. Ecol. Evol. 2020, 10, 14282–14299. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, X.; Huang, Z. Behavioral adjustments and support use of François’ langur in limestone habitat in Fusui, China: Implications for behavioral thermoregulation. Ecol. Evol. 2020, 10, 4956–4967. [Google Scholar] [CrossRef]
- Alberts, S.C. Social influences on survival and reproduction: Insights from a long-term study of wild baboons. J. Anim. Ecol. 2019, 88, 47–66. [Google Scholar] [CrossRef]
- Hill, R.A.; Dunbar, R.I.M. An evaluation of the roles of predation rate and predation risk as selective pressures on primate grouping behaviour. Behaviour 1998, 135, 411–430. [Google Scholar] [CrossRef]
- Roberts, S.G.B.; Roberts, A.I. Social Brain Hypothesis: Vocal and Gesture Networks of Wild Chimpanzees. Front. Psychol. 2016, 7, 1756. [Google Scholar] [CrossRef]
- Dunbar, R.I.M. The Social Brain Hypothesis and Human Evolution. Available online: https://oxfordre.com/psychology/view/10.1093/acrefore/9780190236557.001.0001/acrefore-9780190236557-e-44 (accessed on 4 May 2021).
- Dunbar, R.I.M. The social role of touch in humans and primates: Behavioural function and neurobiological mechanisms. Neurosci. Biobehav. Rev. 2010, 34, 260–268. [Google Scholar] [CrossRef]
- Graves, F.; Wallen, K.; Maestripieri, D. Opioids and attachment in rhesus macaque (Macaca mulatta) abusive mothers. Behav. Neurosci. 2002, 116, 489–493. [Google Scholar] [CrossRef]
- Keverne, E.B.; Martensz, N.D.; Tuite, B. Beta-endorphin concentrations in cerebrospinal fluid of monkeys are influenced by grooming relationships. Psychoneuroendocrinology 1989, 14, 155–161. [Google Scholar] [CrossRef]
- Martel, F.L.; Nevison, C.M.; Simpson, M.J.; Keverne, E.B. Effects of opioid receptor blockade on the social behavior of rhesus monkeys living in large family groups. Dev. Psychobiol. 1995, 28, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C. The Ape in Our House; Harper: New York, NY, USA, 1951; p. 247. [Google Scholar]
- Falk, J.L. The grooming behavior of the chimpanzee as a reinforcer. J. Exp. Anal. Behav. 1958, 1, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.; Graham, K.E.; Sakamaki, T.; Furuichi, T. Long-sightedness in old wild bonobos during grooming. Curr. Biol. 2016, 26, R1131–R1132. [Google Scholar] [CrossRef][Green Version]
- Fujisawa, M.; Matsubayashi, K.; Soumah, A.G.; Kasahara, Y.; Nakatsuka, M.; Matsuzawa, T. Farsightedness (presbyopia) in a wild elderly chimpanzee: The first report. Geriatr. Gerontol. Int. 2010, 10, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, K. Who’s who. In Mahale Chimpanzees: 50 Years of Research; Nakamura, M., Hosaka, K., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 48–67. [Google Scholar]
- Nakamura, M.; Nishie, H. Death of the oldest female at mahale and some notes about longevity of wild chimpanzees. Pan Afr. News 2012, 19, 19–21. [Google Scholar] [CrossRef]
- Lampert, R.M.S.; de Azevedo, C.V.M.; Menezes, A.A.L. Influence of different light intensities on the daily grooming distribution of common marmosets Callithrix jacchus. Folia Primatol. 2011, 82, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Charles-Dominique, P. Ecology and Behavior of Nocturnal Primates; Columbia University Press: New York, NY, USA, 1977; p. 277. [Google Scholar]
- Boesch, C. The Real Chimpanzee: Sex Strategies in the Forest; Cambridge University Press: Cambridge, UK, 2009; p. 196. [Google Scholar]
- Nishida, T.; Mitani, J.C.; Watts, D.P. Variable grooming behaviours in wild chimpanzees. Folia Primatol. 2004, 75, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Zamma, K. Leaf-grooming by a wild chimpanzee in Mahale. Primates 2002, 43, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Yerkes, R.M. Genetic Aspects of Grooming, a Socially Important Primate Behavior Pattern. J. Soc. Psychol. 1933, 4, 3–23. [Google Scholar] [CrossRef]
- Tremblay, L.; Schultz, W. Relative reward preference in primate orbitofrontal cortex. Nature 1999, 398, 704–708. [Google Scholar] [CrossRef]
- Shah, S.Z.; Jabbar, B.; Ahmed, N.; Rehman, A.; Nasir, H.; Nadeem, S.; Jabbar, I.; Rahman, Z.U.; Azam, S. Epidemiology, Pathogenesis, and Control of a Tick-Borne Disease- Kyasanur Forest Disease: Current Status and Future Directions. Front. Cell Infect. Microbiol. 2018, 8, 149. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Shultz, S. Why are there so many explanations for primate brain evolution? Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160244. [Google Scholar] [CrossRef] [PubMed]
- Shultz, S.; Finlayson, L.V. Large body and small brain and group sizes are associated with predator preferences for mammalian prey. Behav. Ecol. 2010, 21, 1073–1079. [Google Scholar] [CrossRef]
- Shultz, S.; Noë, R.; McGraw, W.; Dunbar, R.I.M. A community-level evaluation of the impact of prey behavioural and ecological characteristics on predator diet composition. Proc. R. Soc. Lond. Biol. Sci. 2004, 271, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, S.K.S.; McClymont, J.M.; Crompton, R.H. The arboreal origins of human bipedalism. Antiquity 2014, 88, 906–914. [Google Scholar] [CrossRef]
- Schmitt, D. Insights into the evolution of human bipedalism from experimental studies of humans and other primates. J. Exp. Biol. 2003, 206, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Filler, A.G. Homeotic Evolution in the Mammalia: Diversification of Therian Axial Seriation and the Morphogenetic Basis of Human Origins. PLoS ONE 2007, 2, e1019. [Google Scholar] [CrossRef] [PubMed]
- Prang, T.C. The African ape-like foot of Ardipithecus ramidus and its implications for the origin of bipedalism. Elife 2019, 8. [Google Scholar] [CrossRef]
- Pilbeam, D.R.; Lieberman, D.E. Reconstructing the Last Common Ancestor of Chimpanzees and Humans. In Chimpanzees and Human Evolution; Muller, M.N., Wrangham, R.W., Pilbeam, D.R., Eds.; Harvard University Press: Cambridge, MA, USA, 2017; p. 839. [Google Scholar]
- Püschel, T.A.; Marcé-Nogué, J.; Chamberlain, A.T.; Yoxall, A.; Sellers, W.I. The biomechanical importance of the scaphoid-centrale fusion during simulated knuckle-walking and its implications for human locomotor evolution. Sci. Rep. 2020, 10, 3526. [Google Scholar] [CrossRef]
- Tuttle, R.H. Evolution of hominid bipedalism and prehensile capabilities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1981, 292, 89–94. [Google Scholar] [CrossRef]
- Morton, D.J. Evolution of man’s erect posture (preliminary report). J. Morphol. 1926, 43, 147–179. [Google Scholar] [CrossRef]
- Cannon, C.H.; Leighton, M. Comparative locomotor ecology of gibbons and macaques: Selection of canopy elements for crossing gaps. Am. J. Phys. Anthropol. 1994, 93, 505–524. [Google Scholar] [CrossRef]
- Vereecke, E.E.; D’Août, K.; Aerts, P. Studying Captive Ape Locomotion: Past, Present, and Future. In Primate Locomotion: Linking Field and Laboratory Research; D’Août, K., Vereecke, E.E., Eds.; Springer Science: New York, NY, USA, 2011; pp. 29–46. [Google Scholar]
- Fan, P.; Scott, M.B.; Fei, H.; Ma, C. Locomotion behavior of cao vit gibbon (Nomascus nasutus) living in karst forest in Bangliang Nature Reserve, Guangxi, China. Integr. Zool. 2013, 8, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Vereecke, E.E.; Août, K.; Aerts, P. The dynamics of hylobatid bipedalism: Evidence for an energy-saving mechanism? J. Exp. Biol. 2006, 209, 2829. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schmitt, D. Compliant walking in primates. J. Zool. 1999, 248, 149–160. [Google Scholar] [CrossRef]
- Demes, B.; Jungers, W.; Nieschalk, U. Size- and speed related aspects of quadrupedal walking in slender and slow lorises. In Gravity, Posture and Locomotion in Primates; Jouffrey, F., Stack, M., Niemitz, C., Eds.; Editrice Il Sedicesimo Firenze: Florence, Italy, 1990; pp. 175–197. [Google Scholar]
- McLester, J.; St. Pierre, P. Applied Biomechanics: Concepts and Connections, 2nd ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2019; p. 506. [Google Scholar]
- Pratt, J. Exploiting Natural Dynamics in the Control of a 3D Bipedal Walking Simulation. In Proceedings of the International Conference on Climbing and Walking Robots(CLAWAR99), Portsmouth, UK; Randall, M.J., Ed.; John Wiley & Sons: Chichester, West Sussex, UK, 1999. [Google Scholar]
- Vereecke, E.E.; Aerts, P. The mechanics of the gibbon foot and its potential for elastic energy storage during bipedalism. J. Exp. Biol. 2008, 211, 3661. [Google Scholar] [CrossRef] [PubMed]
- Crompton, R.H.; Sellers, W.I.; Thorpe, S.K.S. Arboreality, Terrestriality and Bipedalism. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3301–3314. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, R.H.; Cortright, G.W. Positional behaviour, adaptive complexes, and evolution. In Orang-Utan Biology; Schwartz, J.H., Ed.; Oxford University Press: Oxford, UK, 1988; pp. 311–330. [Google Scholar]
- Susman, R.L. Facultative terrestrial hand postures in an orangutan (Pongo pygmaeus) and pongid evolution. Am. J. Phys. Anthropol. 1974, 40, 27–37. [Google Scholar] [CrossRef]
- Böhme, M.; Spassov, N.; DeSilva, J.M.; Begun, D.R. Reply to: Reevaluating bipedalism in Danuvius. Nature 2020, 586, E4–E5. [Google Scholar] [CrossRef]
- Kivell, T.L. Fossil ape hints at how walking on two feet evolved. Nature 2019, 575, 445–446. [Google Scholar] [CrossRef]
- Richmond, B.G.; Strait, D.S. Evidence that humans evolved from a knuckle-walking ancestor. Nature 2000, 404, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Kivell, T.L.; Schmitt, D. Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor. Proc. Natl. Acad. Sci. USA 2009, 106, 14241. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.W.; Latimer, B.; Lovejoy, C.O. Why Do Knuckle-Walking African Apes Knuckle-Walk? Anat. Rec. 2018, 301, 496–514. [Google Scholar] [CrossRef] [PubMed]
- Prang, T.C.; Ramirez, K.; Grabowski, M.; Williams, S.A. Ardipithecus hand provides evidence that humans and chimpanzees evolved from an ancestor with suspensory adaptations. Sci. Adv. 2021, 7, eabf2474. [Google Scholar] [CrossRef]
- Corruccini, R.S. Morphometric affinities in the forelimb of Anthropoid primates. Z. Für Morphol. Und Anthropol. 1975, 67, 19–31. [Google Scholar]
- Crompton, R.H.; Vereecke, E.E.; Thorpe, S.K.S. Locomotion and posture from the common hominoid ancestor to fully modern hominins, with special reference to the last common panin/hominin ancestor. J. Anat. 2008, 212, 501–543. [Google Scholar] [CrossRef]
- McCollum, M.A.; Rosenman, B.A.; Suwa, G.; Meindl, R.S.; Lovejoy, C.O. The vertebral formula of the last common ancestor of African apes and humans. J. Exp. Zool. B Mol. Dev. Evol. 2010, 314, 123–134. [Google Scholar] [CrossRef]
- Williams, S.A.; Middleton, E.R.; Villamil, C.I.; Shattuck, M.R. Vertebral numbers and human evolution. Am. J. Phys. Anthropol. 2016, 159, 19–36. [Google Scholar] [CrossRef]
- Machnicki, A.L.; Reno, P.L. Great apes and humans evolved from a long-backed ancestor. J. Hum. Evol. 2020, 144, 102791. [Google Scholar] [CrossRef]
- Grüneberg, C.; Bloem, B.R.; Honegger, F.; Allum, J.H.J. The influence of artificially increased hip and trunk stiffness on balance control in man. Exp. Brain Res. 2004, 157, 472–485. [Google Scholar] [CrossRef]
- Poliero, T.; Lazzaroni, M.; Toxiri, S.; Di Natali, C.; Caldwell, D.G.; Ortiz, J. Applicability of an active back-support exoskeleton to carrying activities. Front. Robot. Ai 2020, 7, 157. [Google Scholar] [CrossRef] [PubMed]
- DeSilva, J.M. First Steps: How Upright Walking Made Us Human; Harper: New York, NY, USA, 2021; p. 346. [Google Scholar]
- Williams, S.A.; Prang, T.C.; Meyer, M.R.; Russo, G.A.; Shapiro, L.J. Reevaluating bipedalism in Danuvius. Nature 2020, 586, E1–E3. [Google Scholar] [CrossRef]
- Alexander, R.M. Characteristics and advantages of human bipedalism. In Biomechanics in Evolution; Rayner, J.M.V., Wooton, R.J., Eds.; Cambridge Univesity Press: Cambridge, UK, 1991; pp. 225–266. [Google Scholar]
- Barak, M.M.; Lieberman, D.E.; Raichlen, D.; Pontzer, H.; Warrener, A.G.; Hublin, J.J. Trabecular evidence for a human-like gait in Australopithecus africanus. PLoS ONE 2013, 8, e77687. [Google Scholar] [CrossRef]
- Carey, T.S.; Crompton, R.H. The metabolic costs of ‘bent-hip, bent-knee’ walking in humans. J. Hum. Evol. 2005, 48, 25–44. [Google Scholar] [CrossRef]
- Raichlen, D.A.; Gordon, A.D.; Harcourt-Smith, W.E.H.; Foster, A.D.; Haas Jr, W.R. Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics. PLoS ONE 2010, 5, e9769. [Google Scholar] [CrossRef]
- Ward, C.V.; Kimbel, W.H.; Johanson, D.C. Complete fourth metatarsal and arches in the foot of Australopithecus afarensis. Science 2011, 331, 750–753. [Google Scholar] [CrossRef] [PubMed]
- LeVeau, B.F. Biomechanics of Human Motion (Basics and Beyond for the Health Professions), 1st ed.; Slack Incorporated: Thorofare, NJ, USA, 2010. [Google Scholar]
- McNutt, E.J.; Zipfel, B.; DeSilva, J.M. The evolution of the human foot. Evol. Anthropol. Issues News Rev. 2018, 27, 197–217. [Google Scholar] [CrossRef] [PubMed]
- Farris, D.J.; Kelly, L.A.; Cresswell, A.G.; Lichtwark, G.A. The functional importance of human foot muscles for bipedal locomotion. Proc. Natl. Acad. Sci. USA 2019, 116, 1645. [Google Scholar] [CrossRef]
- Griffin, N.L.; Miller, C.E.; Schmitt, D.; D’aout, K. Understanding the evolution of the windlass mechanism of the human foot from comparative anatomy: Insights, obstacles, and future directions. Am. J. Phys. Anthropol. 2015, 156, 1–10. [Google Scholar] [CrossRef]
- Fleagle, J.G. Primate Adaptation and Evolution, 3rd ed.; Elsevier: New York, NY, USA, 2013. [Google Scholar]
- Chan, C.W.; Rudins, A. Foot biomechanics during walking and running. Mayo Clin. Proc. 1994, 69, 448–461. [Google Scholar] [CrossRef]
- Kuo, A.D.; Donelan, J.M.; Ruina, A. Energetic Consequences of Walking Like an Inverted Pendulum: Step-to-Step Transitions. Exerc. Sport Sci. Rev. 2005, 33, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.D.; Raichlen, D.A.; Pontzer, H. Muscle force production during bent-hip, bent-knee walking in humans. J. Hum. Evol. 2013, 65, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Griffin, T.M.; Roberts, T.J.; Kram, R. Metabolic cost of generating muscular force in human walking: Insights from load-carrying and speed experiments. J. Appl. Physiol. 2003, 95, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Crompton, R.H. The role of load-carrying in the evolution of modern body proportions. J. Anat. 2004, 204, 417–430. [Google Scholar] [CrossRef]
- Watson, J.; Payne, R.; Chamberlain, A.; Jones, R.; Sellers, W.I. The kinematics of load carrying in humans and great apes: Implications for the evolution of human bipedalism. Folia Primatol. 2009, 80, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.M. Stride length and speed for adults, children, and fossil hominids. Am. J. Phys Anthr. 1984, 63, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, C.O. The natural history of human gait and posture. Part 1. Spine and pelvis. Gait Posture 2005, 21, 95–112. [Google Scholar] [CrossRef]
- Susman, R.L.; Stern, J.T.; Jungers, W.L. Arboreality and bipedality in the hadar hominids. Folia Primatol. 1984, 43, 113–156. [Google Scholar] [CrossRef]
- Watson, J.C.; Payne, R.C.; Chamberlain, A.T.; Jones, R.K.; Sellers, W.I. The energetic costs of load-carrying and the evolution of bipedalism. J. Hum. Evol. 2008, 54, 675–683. [Google Scholar] [CrossRef]
- Rosenberg, K.R.; Golinkoff, R.M.; Zosh, J.M. Did australopithecines (or early Homo) sling? Behav. Brain Sci. 2004, 27, 522. [Google Scholar] [CrossRef]
- Wall-Scheffler, C.M.; Geiger, K.; Steudel-Numbers, K.L. Infant carrying: The role of increased locomotory costs in early tool development. Am. J. Phys. Anthropol. 2007, 133, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.D.; Cant, J.G.H.; Gebo, D.L.; Rose, M.D.; Walker, S.E.; Youlatos, D. Standardized descriptions of primate locomotor and postural modes. Primates 1996, 37, 363–387. [Google Scholar] [CrossRef]
- Hunt, K.D. Ecological Morphology of Australopithecus afarensis. In Primate Locomotion: Recent Advances; Strasser, E., Fleagle, J.G., Rosenberger, A.L., McHenry, H.M., Eds.; Springer US: Boston, MA, USA, 1998; pp. 397–418. [Google Scholar]
- Strait, D.S.; Weber, G.W.; Neubauer, S.; Chalk, J.; Richmond, B.G.; Lucas, P.W.; Spencer, M.A.; Schrein, C.; Dechow, P.C.; Ross, C.F.; et al. The feeding biomechanics and dietary ecology of Australopithecus africanus. Proc. Natl. Acad. Sci. USA 2009, 106, 2124–2129. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.D. Chimpanzee: Lessons from our Sister Species; Cambridge University Press: Cambridge, UK, 2020; p. 592. [Google Scholar]
- Pontzer, H.; Wrangham, R.W. Climbing and the daily energy cost of locomotion in wild chimpanzees: Implications for hominoid locomotor evolution. J. Hum. Evol. 2004, 46, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Pontzer, H. Locomotor Ecology and Evolution in Chimpanzees and Humans. In Chimpanzees and Human Evolution; Muller, M.N., Wrangham, R.W., Pilbeam, D.R., Eds.; Harvard University Press: Cambridge, MA, USA, 2017; p. 839. [Google Scholar]
- DeSilva, J.M. Functional morphology of the ankle and the likelihood of climbing in early hominins. Proc. Natl. Acad. Sci. USA 2009, 106, 6567–6572. [Google Scholar] [CrossRef]
- Venkataraman, V.V.; Kraft, T.S.; Dominy, N.J. Tree climbing and human evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 1237–1242. [Google Scholar] [CrossRef]
- Stern, J.T.; Larson, S.G. Electromyographic study of the obturator muscles in non-human primates: Implications for interpreting the obturator externus groove of the femur. J. Hum. Evol. 1993, 24, 403–427. [Google Scholar] [CrossRef]
- Crompton, R.H.; Li, Y.; Thorpe, S.K.; Wang, W.J.; Savage, R.; Payne, R. The biomechanical evolution of erect bipedality. Cour. Forsch. Senckenberg 2003, 243, 115–126. [Google Scholar]
- Isler, K. Characteristics of vertical climbing in African apes. Senckenbergiana Lethaea 2002, 82, 115–124. [Google Scholar] [CrossRef]
- Nakano, Y.; Hirasaki, E.; Kumakura, H. Patterns of Vertical Climbing in Primates. In Human Origins and Environmental Backgrounds; Ishida, H., Tuttle, R., Pickford, M., Ogihara, N., Nakatsukasa, M., Eds.; Springer: New York, NY, USA, 2006; pp. 97–104. [Google Scholar]
- Preuschoft, H.; Witte, H. Biomechanical Reasons for the Evolution of the Hominid Body Plan. In Origine(s) de la Bipédie Chez Les Hominidés; Coppens, Y., Senut, B., Eds.; Centre National de la Recherche Scientifique: Paris, France, 1991; pp. 59–77. [Google Scholar]
- Adams, M.E.; Jenkins, D.L. Earliest record of the spinose ear tick, Otobius megnini (Dugès) (Ixodida: Argasidae), from the Paisley Caves site (Oregon, USA), with implications for prehistoric human ectoparasitism. J. Archaeol. Sci. 2020, 116, 105114. [Google Scholar] [CrossRef]
- Abbasi, A.A. Molecular evolution of HR, a gene that regulates the postnatal cycle of the hair follicle. Sci. Rep. 2011, 1, 32. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Z.; Xu, S.; Zhou, K.; Yang, G. Characterization of hairless (Hr) and FGF5 genes provides insights into the molecular basis of hair loss in Cetaceans. BMC Evol. Biol. 2013, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Gaudry, M.J.; Campbell, K.L.; Jastroch, M. Evolution of UCP1. Handb. Exp. Pharm. 2019, 251, 127–141. [Google Scholar] [CrossRef]
- Rogers, J.; Gibbs, R.A. Comparative primate genomics: Emerging patterns of genome content and dynamics. Nat. Rev. Genet. 2014, 15, 347–359. [Google Scholar] [CrossRef]
- Galili, U. Evolution in primates by “Catastrophic-selection” interplay between enveloped virus epidemics, mutated genes of enzymes synthesizing carbohydrate antigens, and natural anti-carbohydrate antibodies. Am. J. Phys. Anthropol. 2019, 168, 352–363. [Google Scholar] [CrossRef]
- Galili, U.; Swanson, K. Gene sequences suggest inactivation of alpha-1,3-galactosyltransferase in catarrhines after the divergence of apes from monkeys. Proc. Natl. Acad. Sci. USA 1991, 88, 7401–7404. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Hodžić, A.; Román-Carrasco, P.; Mateos-Hernández, L.; Duscher, G.G.; Sinha, D.K.; Hemmer, W.; Swoboda, I.; Estrada-Peña, A.; de la Fuente, J. Environmental and Molecular Drivers of the α-Gal Syndrome. Front. Immunol. 2019, 10, 1210. [Google Scholar] [CrossRef]
- Schultz, W.; Tremblay, L.; Hollerman, J.R. Reward Processing in Primate Orbitofrontal Cortex and Basal Ganglia. Cereb. Cortex 2000, 10, 272–283. [Google Scholar] [CrossRef]
- Cooper, J.E. Gorilla Pathology and Health: With a Catalogue of Preserved Materials; Academic Press: London, UK, 2017; p. 666. [Google Scholar]
- Huang, C.; Li, Y. How does the white-headed langur (Trachypithecus leucocephalus) adapt locomotor behavior to its unique limestone hill habitat? Primates 2005, 46, 261–267. [Google Scholar] [CrossRef]
- Nelson, H.; Geher, G. Mutual Grooming in Human Dyadic Relationships: An Ethological Perspective. Curr. Psychol. 2007, 26, 121–140. [Google Scholar] [CrossRef]
- Schwartz, A.M.; Hinckley, A.F.; Mead, P.S.; Hook, S.A.; Kugeler, K.J. Surveillance for Lyme Disease—United States, 2008–2015. Mmwr Surveill. Summ. 2017, 66, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.J.; Hurd, P.L.; Crespi, B.J. Genes underlying altruism. Biol. Lett. 2013, 9, 20130395. [Google Scholar] [CrossRef] [PubMed]
- Bryant, K.A.; Marshall, G.S. Clinical manifestations of tick-borne infections in children. Clin. Diagn Lab. Immunol 2000, 7, 523–527. [Google Scholar] [CrossRef][Green Version]
- Stanton, M.A.; Lonsdorf, E.V.; Pusey, A.E.; Goodall, J.; Murray, C.M. Maternal Behavior by Birth Order in Wild Chimpanzees (Pan troglodytes): Increased Investment by First-Time Mothers. Curr. Anthropol. 2014, 55, 483–489. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, J.G. Ticks, Hair Loss, and Non-Clinging Babies: A Novel Tick-Based Hypothesis for the Evolutionary Divergence of Humans and Chimpanzees. Life 2021, 11, 435. https://doi.org/10.3390/life11050435
Brown JG. Ticks, Hair Loss, and Non-Clinging Babies: A Novel Tick-Based Hypothesis for the Evolutionary Divergence of Humans and Chimpanzees. Life. 2021; 11(5):435. https://doi.org/10.3390/life11050435
Chicago/Turabian StyleBrown, Jeffrey G. 2021. "Ticks, Hair Loss, and Non-Clinging Babies: A Novel Tick-Based Hypothesis for the Evolutionary Divergence of Humans and Chimpanzees" Life 11, no. 5: 435. https://doi.org/10.3390/life11050435
APA StyleBrown, J. G. (2021). Ticks, Hair Loss, and Non-Clinging Babies: A Novel Tick-Based Hypothesis for the Evolutionary Divergence of Humans and Chimpanzees. Life, 11(5), 435. https://doi.org/10.3390/life11050435