
Zinc Sulfate Stimulates Osteogenic Phenotypes in Periosteum-Derived Cells and Co-Cultures of Periosteum-Derived Cells and THP-1 Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture and Differentiation of hPDCs
2.2. Measurement of hPDC Viability in the Presence of Zinc Suflate
2.3. Evaluation of Osteoblastic Differentiation of hPDCs Treated with Zinc Sulfate
2.4. Measurement of MAPK Phosphorylation in hPDCs
2.5. Culture and Osteoclasitc Differentiation of THP-1 Cells
2.6. Measurement of Effects of Zinc Sulfate on Osteoclast Differentiation
2.7. Direct Co-Culture of hPDCs and THP-1 Cells and Effects of Zinc Sulfate on Osteoblastic Phenotypes
2.8. Measurement of RANKL and OPG Levels and the RANKL/OPG Ratio in hPDC and THP-1 Co-Cultures
2.9. Statistical Analysis
3. Results
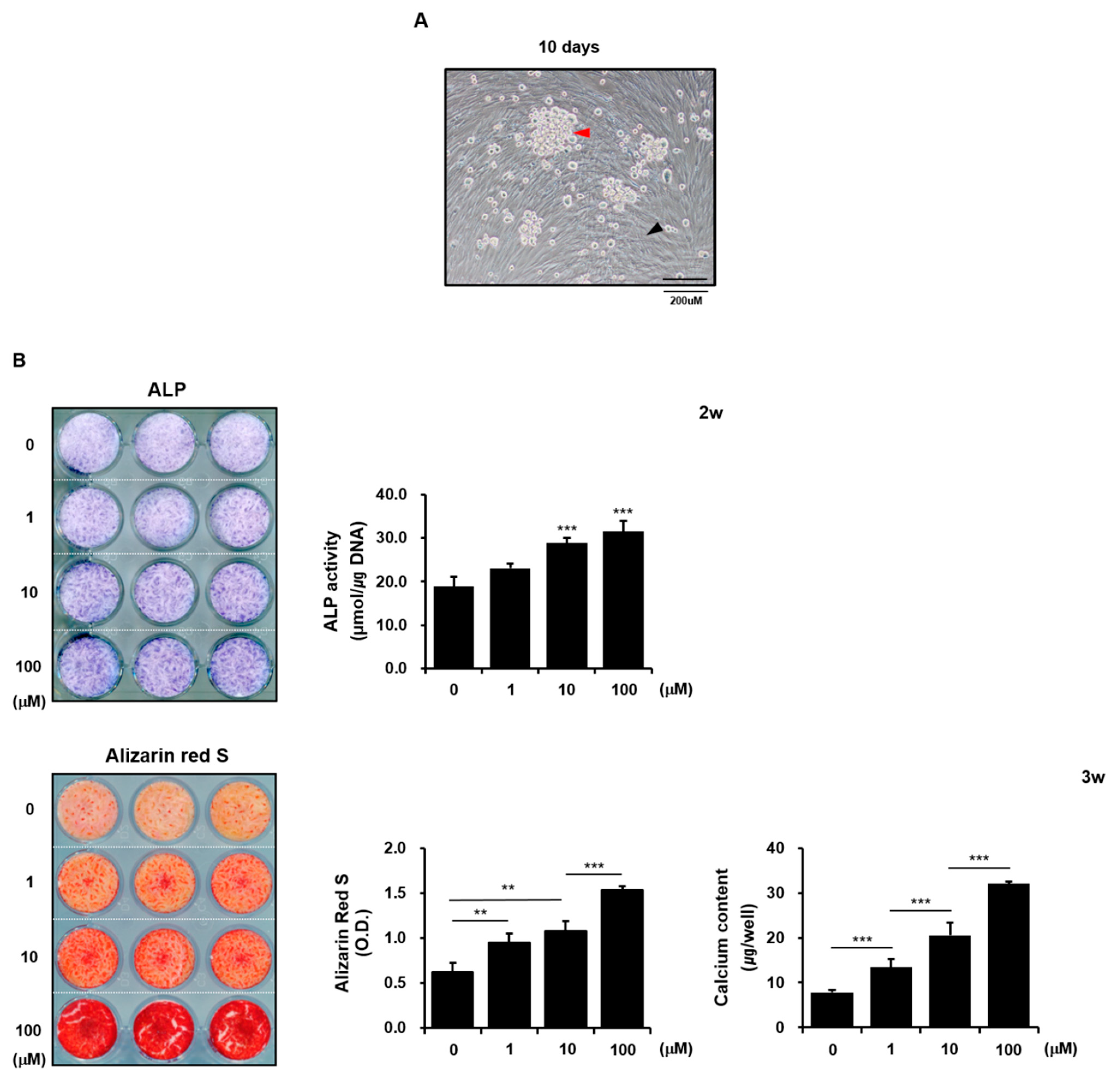
3.1. Zinc Sulfate Promoted Osteoblast Differentiation
3.2. ERK Phosphorylation Was Strongly Induced after Zinc Sulfate Treatment in hPDCs
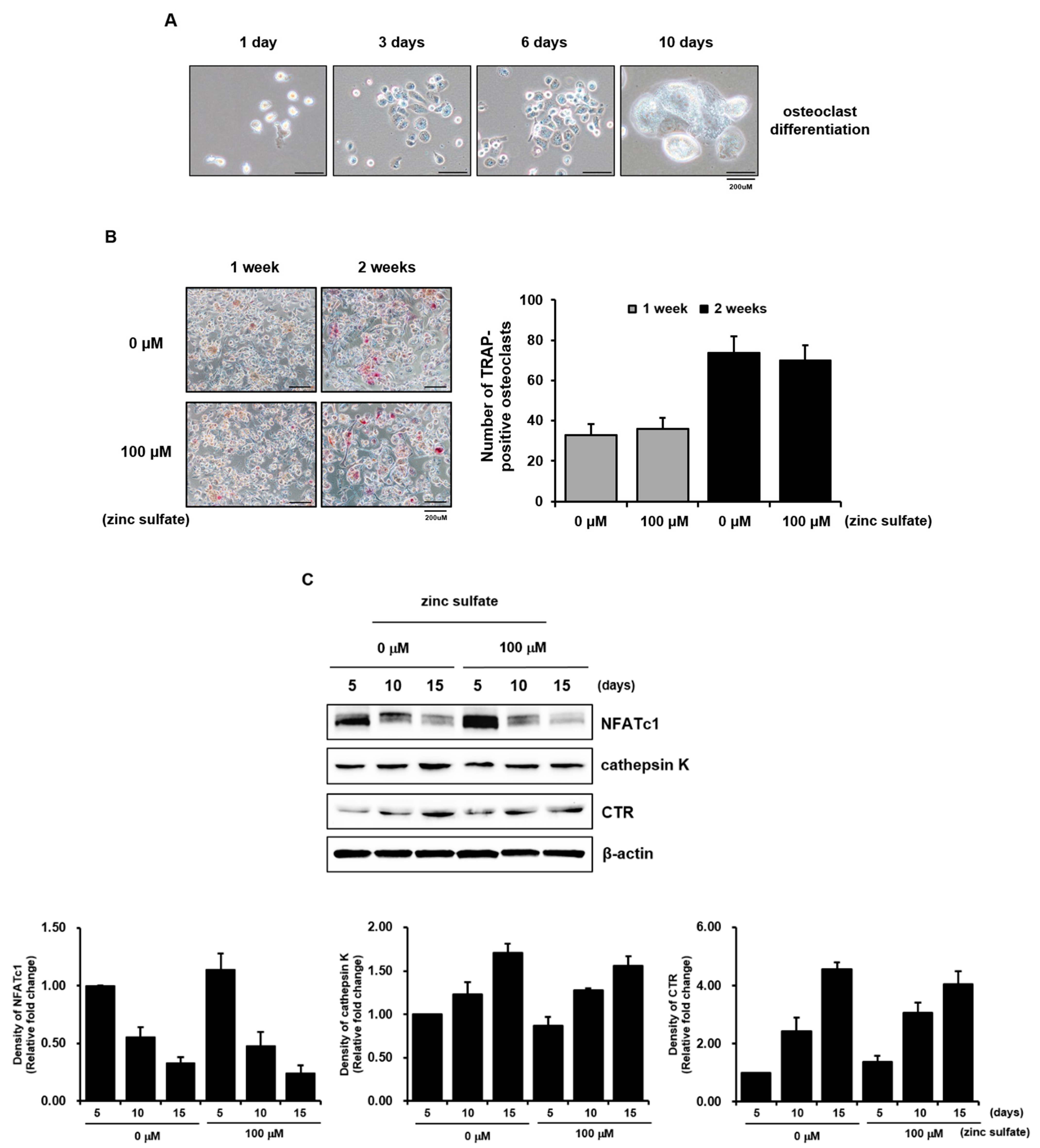
3.3. Zinc Sulfate Did Not Affect the Osteoclastic Differentiation of THP-1 cells
3.4. Zinc Sulfate Promoted Osteogenic Phenotypes in Co-Cultures of hPDCs and THP-1 Cells
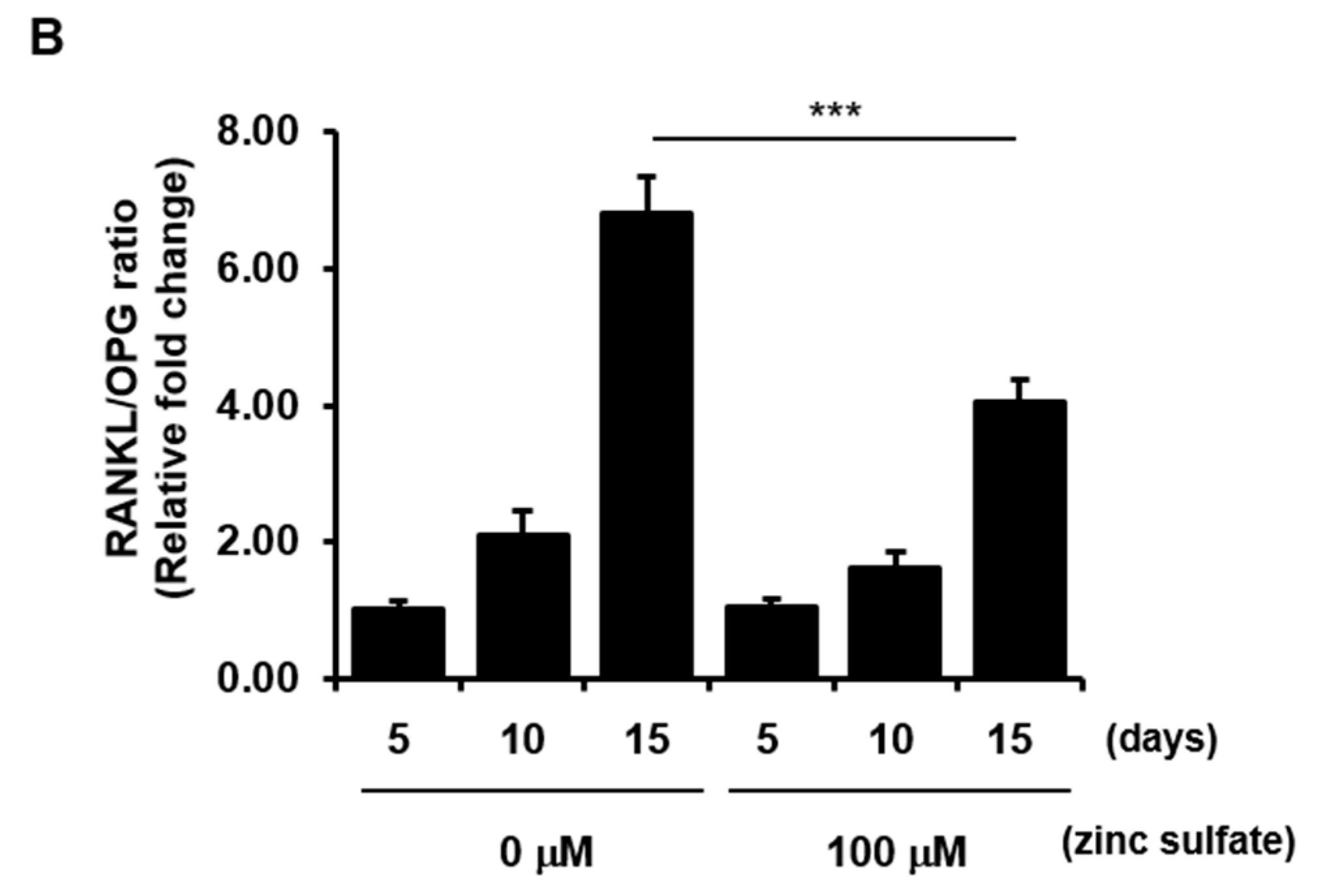
3.5. Effects of Zinc Sulfate on the Levels of RANKL, OPG, and the RANKL/OPG Ratio in Co-Culture of hPDCs and THP-1 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, B.J.; Koh, J.M. Coupling factors involved in preserving bone balance. Cell Mol. Life Sci. 2019, 76, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Roger, P.M. From Crosstalk between Immune and Bone Cells to Bone Erosion in Infection. Int. J. Mol. Sci. 2019, 20, 5154. [Google Scholar] [CrossRef]
- Trohatou, O.; Roubelakis, M.G. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: Past, Present, and Future. Cell Reprogram 2017, 19, 217–224. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise Review: Mesenchymal Stem Cells: From Roots to Boost. Stem Cells 2019, 37, 855–864. [Google Scholar] [CrossRef]
- Yoon, D.K.; Park, J.S.; Rho, G.J.; Lee, H.J.; Sung, I.Y.; Son, J.H.; Park, B.W.; Kang, Y.H.; Byun, S.H.; Hwang, S.C.; et al. The involvement of histone methylation in osteoblastic differentiation of human periosteum-derived cells cultured in vitro under hypoxic conditions. Cell Biochem. Funct. 2017, 35, 441–452. [Google Scholar] [CrossRef]
- Park, B.W.; Hah, Y.S.; Kim, D.R.; Kim, J.R.; Byun, J.H. Osteogenic phenotypes and mineralization of cultured human periosteal-derived cells. Arch. Oral Biol. 2007, 52, 983–989. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J. Bone Tissue Regeneration—Application of Mesenchymal Stem Cells and Cellular and Molecular Mechanisms. Curr. Stem Cell Res. Ther. 2017, 12, 357–364. [Google Scholar] [CrossRef]
- Kim, M.K.; Kim, S.G.; Lee, S.K. 4-Hexylresorcinol-induced angiogenesis potential in human endothelial cells. Maxillofac. Plast Reconstr. Surg. 2020, 42, 23. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M. Role of nutritional zinc in the prevention of osteoporosis. Mol. Cell Biochem. 2010, 338, 241–254. [Google Scholar] [CrossRef]
- Qi, S.; He, J.; Zheng, H.; Chen, C.; Jiang, H.; Lan, S. Zinc Supplementation Increased Bone Mineral Density, Improves Bone Histomorphology, and Prevents Bone Loss in Diabetic Rat. Biol. Trace Elem. Res. 2020, 194, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Chou, J.; Hao, J.; Hatoyama, H.; Ben-Nissan, B.; Milthorpe, B.; Otsuka, M. The therapeutic effect on bone mineral formation from biomimetic zinc containing tricalcium phosphate (ZnTCP) in zinc-deficient osteoporotic mice. PLoS ONE 2013, 8, e71821. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Weitzmann, M.N. Zinc stimulates osteoblastogenesis and suppresses osteoclastogenesis by antagonizing NF-κB activation. Mol. Cell Biochem. 2011, 355, 179–186. [Google Scholar] [CrossRef]
- Park, K.H.; Choi, Y.; Yoon, D.; Lee, K.M.; Kim, D.; Lee, J.W. Zinc Promotes Osteoblast Differentiation in Human Mesenchymal Stem Cells Via Activation of the cAMP-PKA-CREB Signaling Pathway. Stem Cells Dev. 2018, 27, 1125–1135. [Google Scholar] [CrossRef]
- Yusa, K.; Yamamoto, O.; Takano, H.; Fukuda, M.; Iino, M. Zinc-modified titanium surface enhances osteoblast differentiation of dental pulp stem cells in vitro. Sci. Rep. 2016, 6, 29462. [Google Scholar] [CrossRef] [PubMed]
- Dorst, K.; Rammelkamp, D.; Hadjiargyrou, M.; Meng, Y. The Effect of Exogenous Zinc Concentration on the Responsiveness of MC3T3-E1 Pre-Osteoblasts to Surface Microtopography: Part II (Differentiation). Materials 2014, 7, 1097–1112. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, M.B.; Shim, J.H.; Glimcher, L.H. Mitogen-activated protein kinase pathways in osteoblasts. Annu. Rev. Cell Dev. Biol. 2013, 29, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xiao, L.; Peng, J.; Qian, Y.Q.; Huang, C.C. Exosomes derived from bone marrow mesenchymal stem cells improve osteoporosis through promoting osteoblast proliferation via MAPK pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3962–3970. [Google Scholar]
- Kim, B.S.; Kang, H.J.; Park, J.Y.; Lee, J. Fucoidan promotes osteoblast differentiation via JNK- and ERK-dependent BMP2-Smad 1/5/8 signaling in human mesenchymal stem cells. Exp. Mol. Med. 2015, 47, e128. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef]
- Li, Z.H.; Si, Y.; Xu, G.; Chen, X.M.; Xiong, H.; Lai, L.; Zheng, Y.Q.; Zhang, Z.G. High-dose PMA with RANKL and MCSF induces THP-1 cell differentiation into human functional osteoclasts in vitro. Mol. Med. Rep. 2017, 16, 8380–8384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, C.; Liu, C.; Wu, W. NF-κB promotes osteoclast differentiation by overexpressing MITF via down regulating microRNA-1276 expression. Life Sci. 2020, 258, 118093. [Google Scholar] [CrossRef] [PubMed]
- Scian, R.; Barrionuevo, P.; Rodriguez, A.M.; Arriola Benitez, P.C.; García Samartino, C.; Fossati, C.A.; Giambartolomei, G.H.; Delpino, M.V. Brucella abortus invasion of synoviocytes inhibits apoptosis and induces bone resorption through RANKL expression. Infect Immun. 2013, 81, 1940–1951. [Google Scholar] [CrossRef] [PubMed]
- Hayden, R.S.; Quinn, K.P.; Alonzo, C.A.; Georgakoudi, I.; Kaplan, D.L. Quantitative characterization of mineralized silk film remodeling during long-term osteoblast-osteoclast co-culture. Biomaterials 2014, 35, 3794–3802. [Google Scholar] [CrossRef]
- Luo, G.; Li, F.; Li, X.; Wang, Z.G.; Zhang, B. TNF-α and RANKL promote osteoclastogenesis by upregulating RANK via the NF-κB pathway. Mol. Med. Rep. 2018, 17, 6605–6611. [Google Scholar] [CrossRef]
- Abdallah, D.; Jourdain, M.L.; Braux, J.; Guillaume, C.; Gangloff, S.C.; Jacquot, J.; Velard, F. An Optimized Method to Generate Human Active Osteoclasts from Peripheral Blood Monocytes. Front. Immunol. 2018, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.; Abradelo, C.; San Román, J.; Rojo, L. Bibliographic review on the state of the art of strontium and zinc based regenerative therapies. Recent developments and clinical applications. J. Mater. Chem. B 2019, 7, 1974–1985. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Honma, M.; Ikebuchi, Y.; Kariya, Y.; Suzuki, H. Regulatory mechanisms of RANKL presentation to osteoclast precursors. Curr. Osteoporos. Rep. 2014, 12, 115–120. [Google Scholar] [CrossRef]
- Sakamoto, M.; Fukunaga, T.; Sasaki, K.; Seiryu, M.; Yoshizawa, M.; Takeshita, N.; Takano-Yamamoto, T. Vibration enhances osteoclastogenesis by inducing RANKL expression via NF-κB signaling in osteocytes. Bone 2019, 123, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef]
- Liu, H.; Dong, Y.; Gao, Y.; Zhao, L.; Cai, C.; Qi, D.; Zhu, M.; Zhao, L.; Liu, C.; Guo, F.; et al. Hesperetin suppresses RANKL-induced osteoclastogenesis and ameliorates lipopolysaccharide-induced bone loss. J. Cell Physiol. 2019, 234, 11009–11022. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current Understanding of RANK Signaling in Osteoclast Differentiation and Maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar] [PubMed]
- Ihn, H.J.; Kim, K.; Cho, H.S.; Park, E.K. Pentamidine Inhibits Titanium Particle-Induced Osteolysis In Vivo and Receptor Activator of Nuclear Factor-κB Ligand-Mediated Osteoclast Differentiation In Vitro. Tissue Eng. Regen. Med. 2019, 16, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-H.; Park, S.A.; Kang, Y.-H.; Hwa, S.M.; Koh, E.-B.; Hwang, S.-C.; Oh, S.H.; Byun, J.-H. Zinc Sulfate Stimulates Osteogenic Phenotypes in Periosteum-Derived Cells and Co-Cultures of Periosteum-Derived Cells and THP-1 Cells. Life 2021, 11, 410. https://doi.org/10.3390/life11050410
Park J-H, Park SA, Kang Y-H, Hwa SM, Koh E-B, Hwang S-C, Oh SH, Byun J-H. Zinc Sulfate Stimulates Osteogenic Phenotypes in Periosteum-Derived Cells and Co-Cultures of Periosteum-Derived Cells and THP-1 Cells. Life. 2021; 11(5):410. https://doi.org/10.3390/life11050410
Chicago/Turabian StylePark, Jin-Ho, Su A Park, Young-Hoon Kang, So Myeong Hwa, Eun-Byeol Koh, Sun-Chul Hwang, Se Heang Oh, and June-Ho Byun. 2021. "Zinc Sulfate Stimulates Osteogenic Phenotypes in Periosteum-Derived Cells and Co-Cultures of Periosteum-Derived Cells and THP-1 Cells" Life 11, no. 5: 410. https://doi.org/10.3390/life11050410
APA StylePark, J.-H., Park, S. A., Kang, Y.-H., Hwa, S. M., Koh, E.-B., Hwang, S.-C., Oh, S. H., & Byun, J.-H. (2021). Zinc Sulfate Stimulates Osteogenic Phenotypes in Periosteum-Derived Cells and Co-Cultures of Periosteum-Derived Cells and THP-1 Cells. Life, 11(5), 410. https://doi.org/10.3390/life11050410