
Transfer of Antibiotic Resistance Plasmid from Commensal E. coli towards Human Intestinal Microbiota in the M-SHIME: Effect of E. coli dosis, Human Individual and Antibiotic Use


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
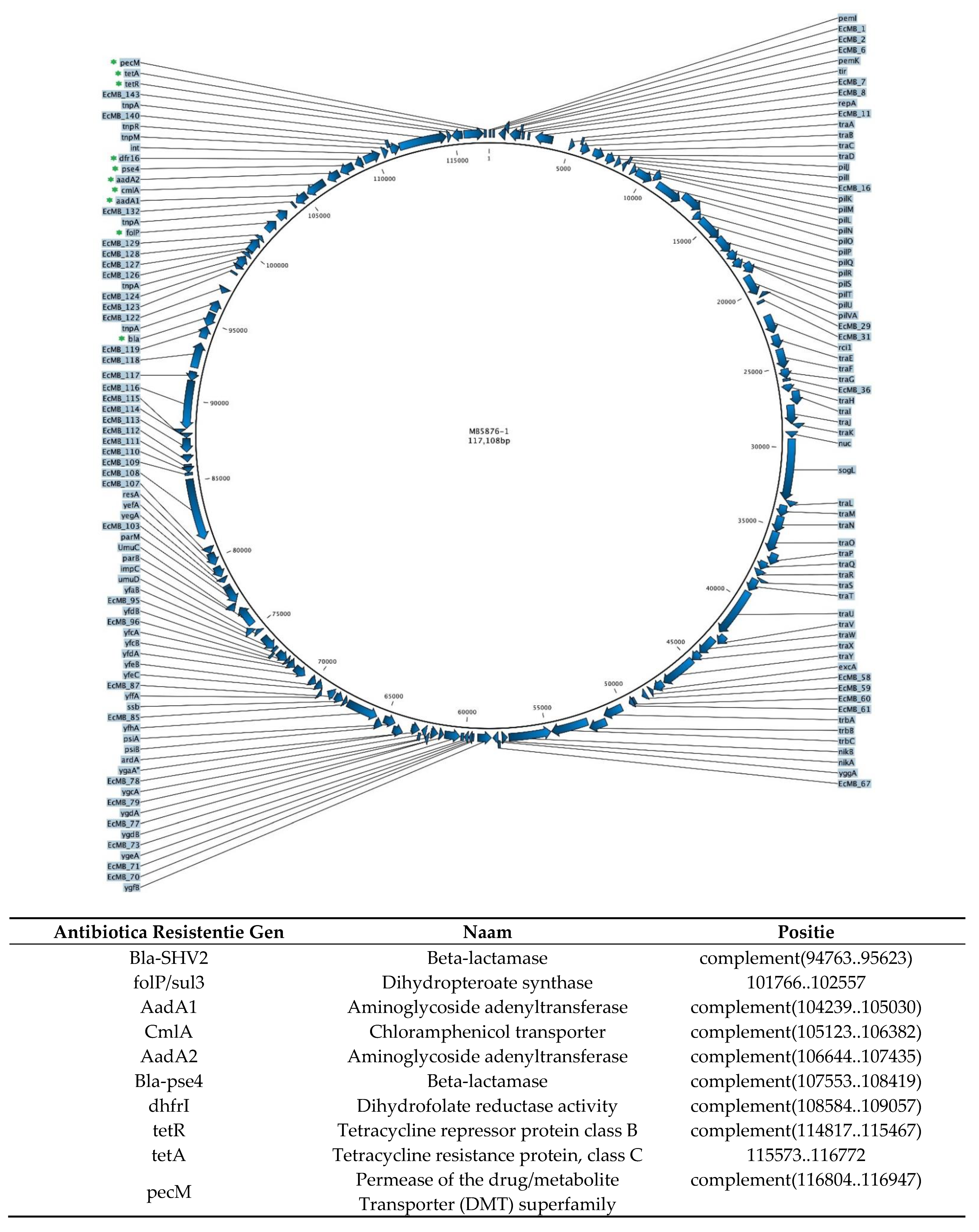
2.1. E. coli MB6212 Plasmid Donor Strain
2.2. Human Fecal Donors
2.3. Mucosal Simulator of the Human Intestinal Ecosystem (M-SHIME)
2.3.1. General Setup
2.3.2. Cefotaxime Treatment
2.4. Analyses of Lumen Samples
2.4.1. Bacterial Counting
2.4.2. Short Chain Fatty Acids
2.4.3. Quantification of Total 16S rRNA Gene Abundance and of p5876 by qPCR
2.4.4. UHPLC-MS/MS
2.5. Statistical Analysis of Bacterial Counts and SCFA Concentrations
2.6. S Amplicon Sequencing and Bioinformatics Analysis
3. Results
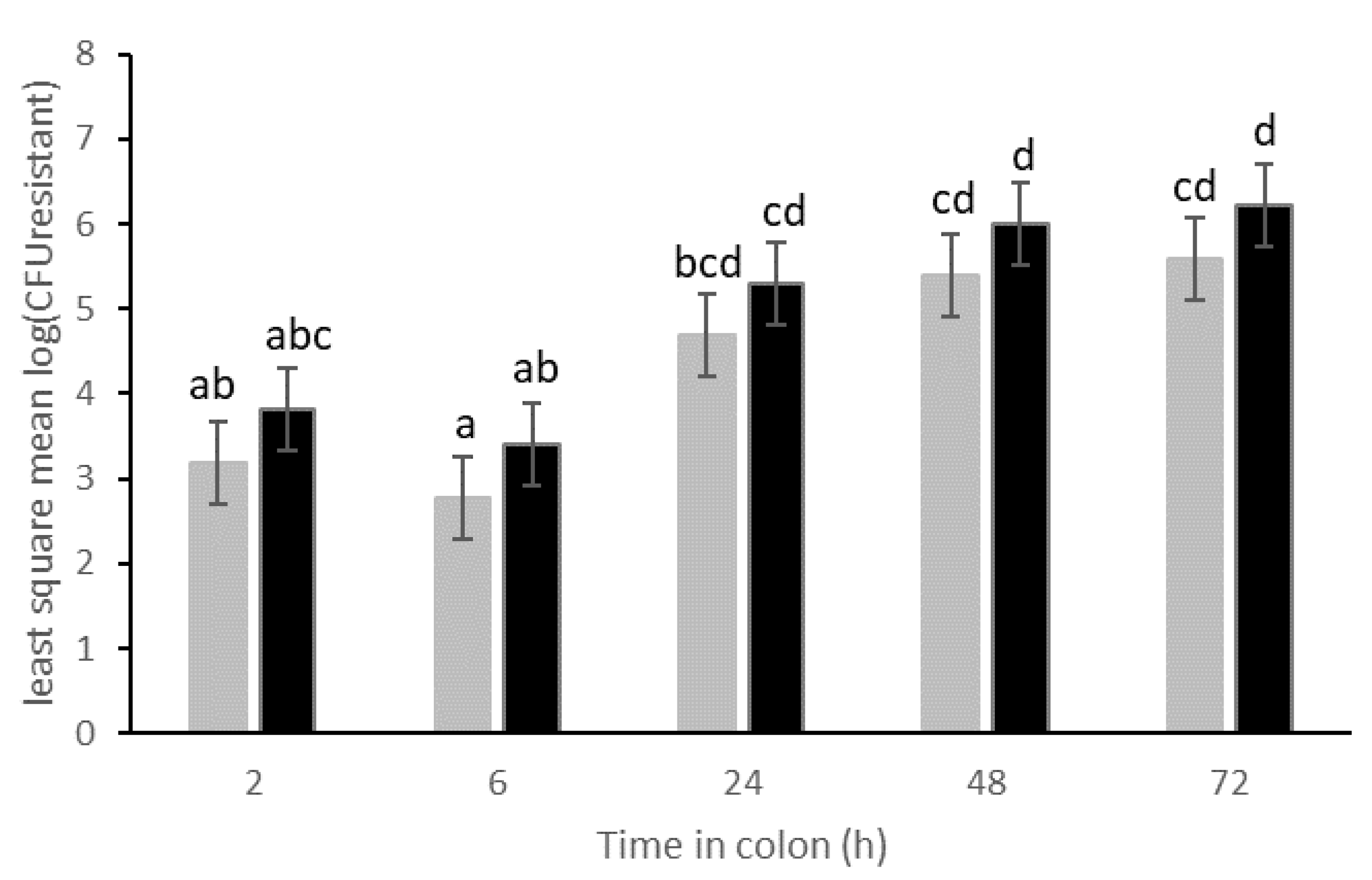
3.1. Transfer of Resistance Plasmid p5876: Effect of Dose and Human Individual in the Absence of Antibiotics
3.2. Transfer of Plasmid p5876 after Cefotaxime Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- European Food Safety Authority. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16, e05182. [Google Scholar]
- Hu, Y.; Gao, G.F.; Zhu, B. The antibiotic resistome: Gene flow in environments, animals and human beings. Front. Med. 2017, 11, 161–168. [Google Scholar] [CrossRef] [PubMed]
- ICF. EU Insights—Perceptions on the human health impact of antimicrobial resistance (AMR) and antibiotics use in animals across the EU. EFSA Supporting Publ. 2017, 14, 1183E. [Google Scholar]
- Hu, Y.; Yang, X.; Baoli, Z.; Lv, N.; Liu, F.; Wu, J.; Lin, I.Y.C.; Wu, N.; Weimer, B.C.; Gao, G.F.; et al. The Bacterial Mobile Resistome Transfer Network Connecting the Animal and Human Microbiomes. Appl. Environ. Microbiol. 2016, 82, 6672–6681. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhu, B. The Human Gut Antibiotic Resistome in the Metagenomic Era: Progress and Perspectives. Infect Dis. Transl. Med. 2016, 2, 41–47. [Google Scholar]
- Stokes, H.W.; Gillings, M.R. Gene flow, mobile genetic elements and the recruitment of antibiotic resistance genes into Gram-negative pathogens. FEMS Microbiol. Rev. 2011, 35, 790–819. [Google Scholar] [CrossRef]
- Jans, C.; Sarno, E.; Collineau, L.; Meile, L.; Stärk, K.D.C.; Stephan, R. Consumer Exposure to Antimicrobial Resistant Bacteria From Food at Swiss Retail Level. Front. Microbiol. 2018, 9, 362. [Google Scholar] [CrossRef]
- De Been, M.; Lanza, V.F.; De Toro, M.; Scharringa, J.; Dohmen, W.; Du, Y.; Hu, J.; Lei, Y.; Li, N.; Tooming-Klunderud, A.; et al. Dissemination of Cephalosporin Resistance Genes between Escherichia coli Strains from Farm Animals and Humans by Specific Plasmid Lineages. PLoS Genet. 2014, 10, e1004776. [Google Scholar] [CrossRef] [PubMed]
- Karanika, S.; Karantanos, T.; Arvanitis, M.; Grigoras, C.; Mylonakis, E. Fecal Colonization with Extended-spectrum Beta-lactamase–ProducingEnterobacteriaceaeand Risk Factors Among Healthy Individuals: A Systematic Review and Metaanalysis. Clin. Infect. Dis. 2016, 63, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Avershina, E.; Rudi, K. Confusion about the species richness of human gut microbiota. Benef. Microbes 2015, 6, 657–659. [Google Scholar] [CrossRef]
- Porse, A.; Gumpert, H.; Kubicek-Sutherland, J.Z.; Karami, N.; Adlerberth, I.; Wold, A.E.; Andersson, D.I.; Sommer, M.O.A. Genome dynamics of Escherichia coli during antibiotic treatment: Transfer, loss, and persistence of genetic elements in situ of the infant gut. Front. Cell. Infect. Microbiol. 2017, 7, 126. [Google Scholar] [CrossRef] [PubMed]
- Bidet, P.; Burghoffer, B.; Gautier, V.; Brahimi, N.; Mariani-Kurkdjian, P.; El-Ghoneimi, A.; Bingen, E.; Arlet, G. In Vivo Transfer of Plasmid-Encoded ACC-1 AmpC from Klebsiella pneumoniae to Escherichia coli in an Infant and Selection of Impermeability to Imipenem in K. pneumoniae. Antimicrob. Agents Chemother. 2005, 49, 3562–3565. [Google Scholar] [CrossRef]
- Karami, N.; Martner, A.; Enne, V.I.; Swerkersson, S.; Adlerberth, I.; Wold, A.E. Transfer of an ampicillin resistance gene between two Escherichia coli strains in the bowel microbiota of an infant treated with antibiotics. J. Antimicrob. Chemother. 2007, 60, 1142–1145. [Google Scholar] [CrossRef]
- Goren, M.G.; Carmeli, Y.; Schwaber, M.J.; Chmelnitsky, I.; Schechner, V.; Navon-Venezia, S. Transfer of Carbapenem-Resistant Plasmid from Klebsiella pneumoniae ST258 to Escherichia coli in Patient. Emerg. Infect. Dis. 2010, 16, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Lopatkin, A.J.; Sysoeva, T.A.; You, L. Dissecting the effects of antibiotics on horizontal gene transfer: Analysis suggests a critical role of selection dynamics. BioEssays 2016, 38, 1283–1292. [Google Scholar] [CrossRef]
- Lambrecht, E.; Van Coillie, E.; Van Meervenne, E.; Boon, N.; Heyndrickx, M.; Van De Wiele, T. Commensal E. coli rapidly transfer antibiotic resistance genes to human intestinal microbiota in the Mucosal Simulator of the Human Intestinal Microbial Ecosystem (M-SHIME). Int. J. Food Microbiol. 2019, 311, 108357. [Google Scholar] [CrossRef]
- Gumpert, H.; Kubicek-Sutherland, J.Z.; Porse, A.; Karami, N.; Munck, C.; Linkevicius, M.; Adlerberth, I.; Wold, A.E.; Andersson, D.I.; Sommer, M.O.A. Transfer and Persistence of a Multi-Drug Resistance Plasmid in situ of the Infant Gut Microbiota in the Absence of Antibiotic Treatment. Front. Microbiol. 2017, 8, 1852. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, E.; Solimini, A.; Pantanella, F.; De Giusti, M.; Cummins, E. Human exposure to antibiotic resistant-Escherichia coli through irrigated lettuce. Environ. Int. 2019, 122, 270–280. [Google Scholar] [CrossRef]
- Depoorter, P.; Persoons, D.; Uyttendaele, M.; Butaye, P.; De Zutter, L.; Dierick, K.; Herman, L.; Imberechts, H.; Van Huffel, X.; Dewulf, J. Assessment of human exposure to 3rd generation cephalosporin resistant E. coli (CREC) through consumption of broiler meat in Belgium. Int. J. Food Microbiol. 2012, 159, 30–38. [Google Scholar] [CrossRef]
- De Boever, P.; Deplancke, B.; Verstraete, W. Fermentation by Gut Microbiota Cultured in a Simulator of the Human Intestinal Microbial Ecosystem Is Improved by Supplementing a Soygerm Powder. J. Nutr. 2000, 130, 2599–2606. [Google Scholar] [CrossRef] [PubMed]
- Vermeiren, J.; Abbeele, P.V.D.; Laukens, D.; Vigsnaes, L.K.; De Vos, M.; Boon, N.; Van De Wiele, T. Decreased colonization of fecal Clostridium coccoides/Eubacterium rectale species from ulcerative colitis patients in an in vitro dynamic gut model with mucin environment. FEMS Microbiol. Ecol. 2011, 79, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Sanofi_aventis. Product Monograph Claforan. 2014. Available online: http://products.sanofi.ca/en/claforan.pdf (accessed on 8 February 2021).
- EUCAST. Available online: https://www.eucast.org/mic_distributions_and_ecoffs/ (accessed on 8 February 2021).
- Andersen, S.J.; Hennebel, T.; Gildemyn, S.; Coma, M.; Desloover, J.; Berton, J.; Tsukamoto, J.; Stevens, C.; Rabaey, K. Electrolytic Membrane Extraction Enables Production of Fine Chemicals from Biorefinery Sidestreams. Environ. Sci. Technol. 2014, 48, 7135–7142. [Google Scholar] [CrossRef]
- De Paepe, K.; Verspreet, J.; Verbeke, K.; Raes, J.; Courtin, C.M.; Van De Wiele, T. Introducing insoluble wheat bran as a gut microbiota niche in an in vitro dynamic gut model stimulates propionate and butyrate production and induces colon region specific shifts in the luminal and mucosal microbial community. Environ. Microbiol. 2018, 20, 3406–3426. [Google Scholar] [CrossRef]
- Ovreås, L.; Forney, L.; Daae, F.L.; Torsvik, V. Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 1997, 63, 3367–3373. [Google Scholar] [CrossRef]
- Geirnaert, A.; Wang, J.; Tinck, M.; Steyaert, A.; Abbeele, P.V.D.; Eeckhaut, V.; Vilchez-Vargas, R.; Falony, G.; Laukens, D.; De Vos, M.; et al. Interindividual differences in response to treatment with butyrate-producing Butyricicoccus pullicaecorum 25–3T studied in an in vitro gut model. FEMS Microbiol. Ecol. 2015, 91, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L. Assessing and Improving Methods Used in Operational Taxonomic Unit-Based Approaches for 16S rRNA Gene Sequence Analysis. Appl. Environ. Microbiol. 2011, 77, 3219–3226. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Hawinkel, S.; Kerckhof, F.-M.; Bijnens, L.; Thas, O. A unified framework for unconstrained and constrained ordination of microbiome read count data. PLoS ONE 2019, 14, e0205474. [Google Scholar] [CrossRef]
- Arbizu, P.M. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 0.3. 2019. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 8 February 2021).
- Dimitriu, T.; Marchant, L.; Buckling, A.; Raymond, B. Bacteria from natural populations transfer plasmids mostly towards their kin. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191110. [Google Scholar] [CrossRef] [PubMed]
- Forns, N.; Baños, R.C.; Balsalobre, C.; Juárez, A.; Madrid, C. Temperature-Dependent Conjugative Transfer of R27: Role of Chromosome- and Plasmid-Encoded Hha and H-NS Proteins. J. Bacteriol. 2005, 187, 3950–3959. [Google Scholar] [CrossRef]
- Frost, L.S.; Manchak, J. F- phenocopies: Characterization of expression of the F transfer region in stationary phase. Microbiology 1998, 144 Pt 9, 2579–2587. [Google Scholar] [CrossRef]
- Zatyka, M.; Thomas, C.M. Control of genes for conjugative transfer of plasmids and other mobile elements. FEMS Microbiol. Rev. 1998, 21, 291–319. [Google Scholar] [CrossRef]
- Serna, A.; Espinosa, E.; Camacho, E.M.; Casadesús, J. Regulation of Bacterial Conjugation in Microaerobiosis by Host-Encoded Functions ArcAB and SdhABCD. Genetics 2010, 184, 947–958. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B.; Schluederberg, S.; Rowse, D.; Davis, J. High frequency of antimicrobial resistance in human fecal flora. Antimicrob. Agents Chemother. 1988, 32, 1801–1806. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef]
- Masco, L.; Van Hoorde, K.; De Brandt, E.; Swings, J.; Huys, G. Antimicrobial susceptibility of Bifidobacterium strains from humans, animals and probiotic products. J. Antimicrob. Chemother. 2006, 58, 85–94. [Google Scholar] [CrossRef]
- Karapetkov, N.; Georgieva, R.; Rumyan, N.; Karaivanova, E. Antibiotic susceptibility of different lactic acid bacteria strains. Benef. Microbes 2011, 2, 335–339. [Google Scholar] [CrossRef]
- Manaia, C.M. Assessing the Risk of Antibiotic Resistance Transmission from the Environment to Humans: Non-Direct Proportionality between Abundance and Risk. Trends Microbiol. 2017, 25, 173–181. [Google Scholar] [CrossRef]
- Frazão, N.; Sousa, A.; Lässig, M.; Gordo, I. Horizontal gene transfer overrides mutation in Escherichia coli colonizing the mammalian gut. Proc. Natl. Acad. Sci. USA 2019, 116, 17906–17915. [Google Scholar] [CrossRef]
- Van den Bogaard, A.E.; Stobberingh, E.E. Epidemiology of resistance to antibiotics. Links between animals and humans. Int. J. Antimicrob. Agents 2000, 14, 327–335. [Google Scholar] [CrossRef]
- Courvalin, P. Transfer of antibiotic resistance genes between gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 1994, 38, 1447–1451. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Hammarén, R.; Pal, C.; Östman, M.; Björlenius, B.; Flach, C.-F.; Fick, J.; Kristiansson, E.; Tysklind, M.; Larsson, D.J. Elucidating selection processes for antibiotic resistance in sewage treatment plants using metagenomics. Sci. Total Environ. 2016, 572, 697–712. [Google Scholar] [CrossRef]
- Murray, A.K.; Zhang, L.; Yin, X.; Zhang, T.; Buckling, A.; Snape, J.; Gaze, W.H. Novel Insights into Selection for Antibiotic Resistance in Complex Microbial Communities. mBio 2018, 9, e00969-18. [Google Scholar] [CrossRef]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef]
- Machado, A.M.D.; Sommer, M.O.A. Human Intestinal Cells Modulate Conjugational Transfer of Multidrug Resistance Plasmids between Clinical Escherichia coli Isolates. PLoS ONE 2014, 9, e100739. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambrecht, E.; Van Coillie, E.; Boon, N.; Heyndrickx, M.; Van de Wiele, T. Transfer of Antibiotic Resistance Plasmid from Commensal E. coli towards Human Intestinal Microbiota in the M-SHIME: Effect of E. coli dosis, Human Individual and Antibiotic Use. Life 2021, 11, 192. https://doi.org/10.3390/life11030192
Lambrecht E, Van Coillie E, Boon N, Heyndrickx M, Van de Wiele T. Transfer of Antibiotic Resistance Plasmid from Commensal E. coli towards Human Intestinal Microbiota in the M-SHIME: Effect of E. coli dosis, Human Individual and Antibiotic Use. Life. 2021; 11(3):192. https://doi.org/10.3390/life11030192
Chicago/Turabian StyleLambrecht, Ellen, Els Van Coillie, Nico Boon, Marc Heyndrickx, and Tom Van de Wiele. 2021. "Transfer of Antibiotic Resistance Plasmid from Commensal E. coli towards Human Intestinal Microbiota in the M-SHIME: Effect of E. coli dosis, Human Individual and Antibiotic Use" Life 11, no. 3: 192. https://doi.org/10.3390/life11030192
APA StyleLambrecht, E., Van Coillie, E., Boon, N., Heyndrickx, M., & Van de Wiele, T. (2021). Transfer of Antibiotic Resistance Plasmid from Commensal E. coli towards Human Intestinal Microbiota in the M-SHIME: Effect of E. coli dosis, Human Individual and Antibiotic Use. Life, 11(3), 192. https://doi.org/10.3390/life11030192