
Biotechnology Applications of Cell-Free Expression Systems



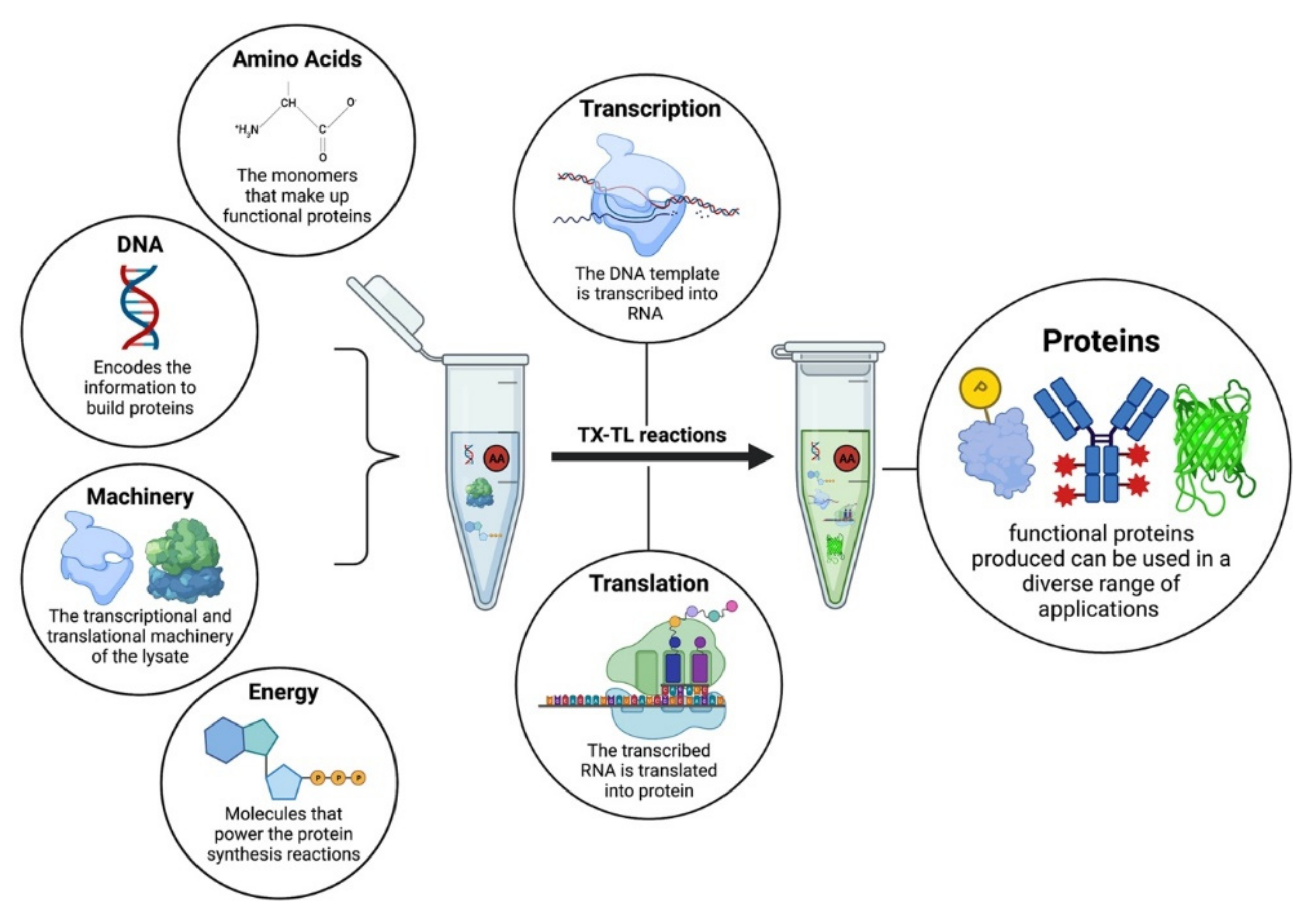{kind=link}
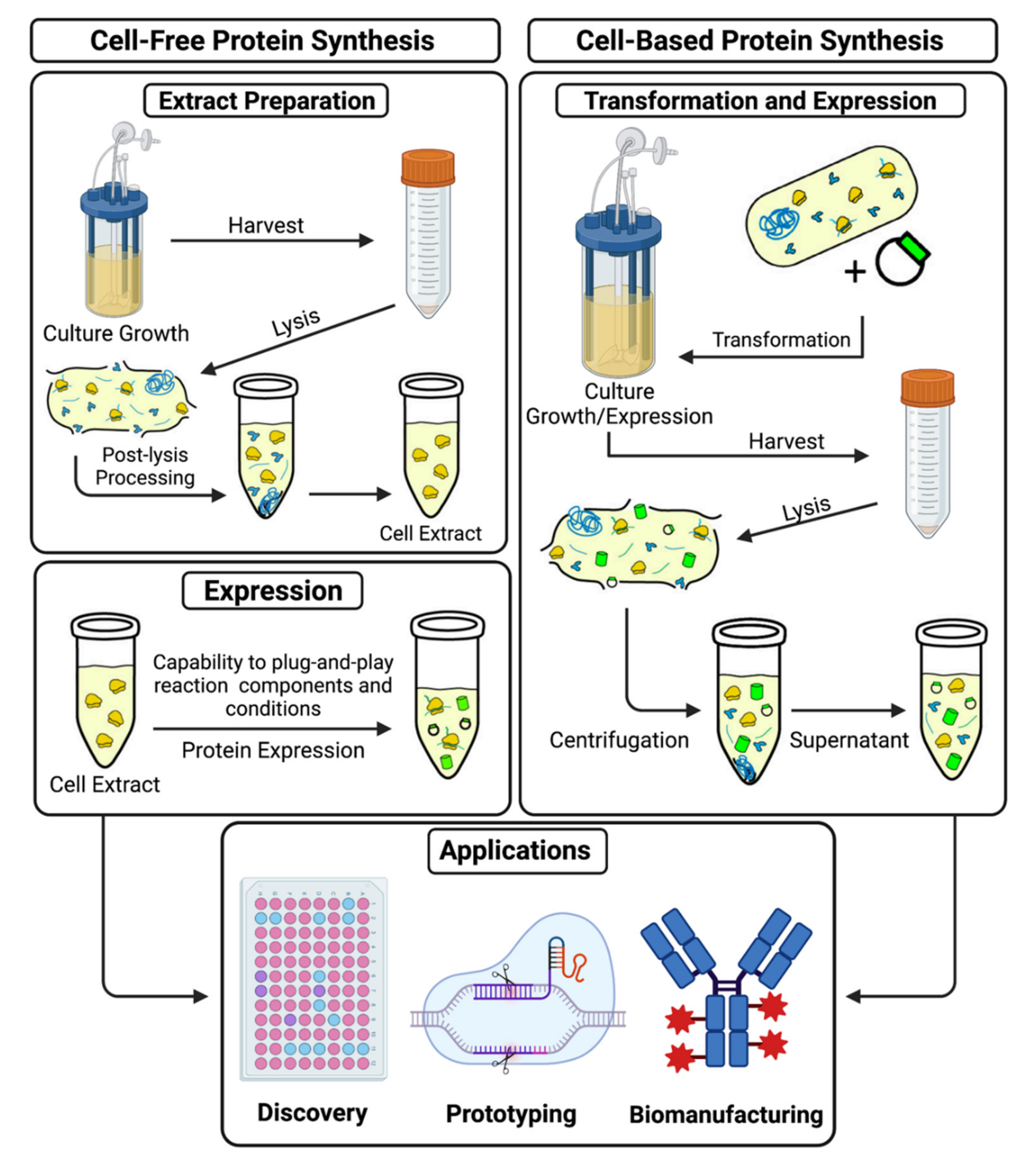{kind=link}
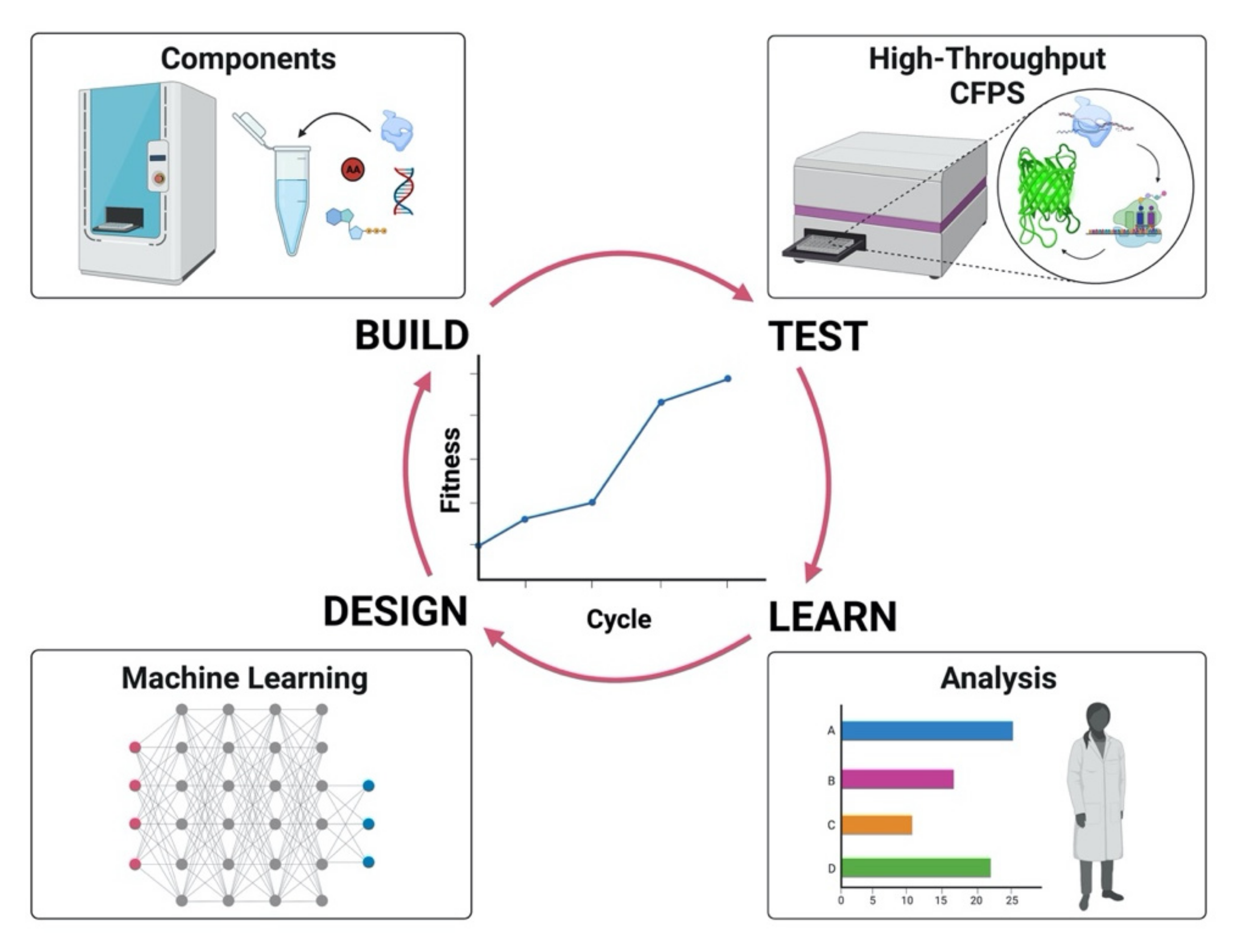{kind=link}
Abstract
:1. Introduction
2. Cell-Free Metabolic Engineering
3. Comparing Extract and Enzyme-Based CFME
4. Cell-Free Prototyping
5. Cell-Free Biosensing
6. Cell-Free Biomanufacturing
6.1. Pharmaceuticals
6.1.1. Monoclonal Antibodies
6.1.2. Antimicrobial Peptides
6.1.3. Vaccines
6.1.4. Small Molecules
6.1.5. Membrane Proteins
6.1.6. On-Demand Production
6.2. Food Biotechnology
6.3. Growing Industries & Industrially Relevant Biomolecules
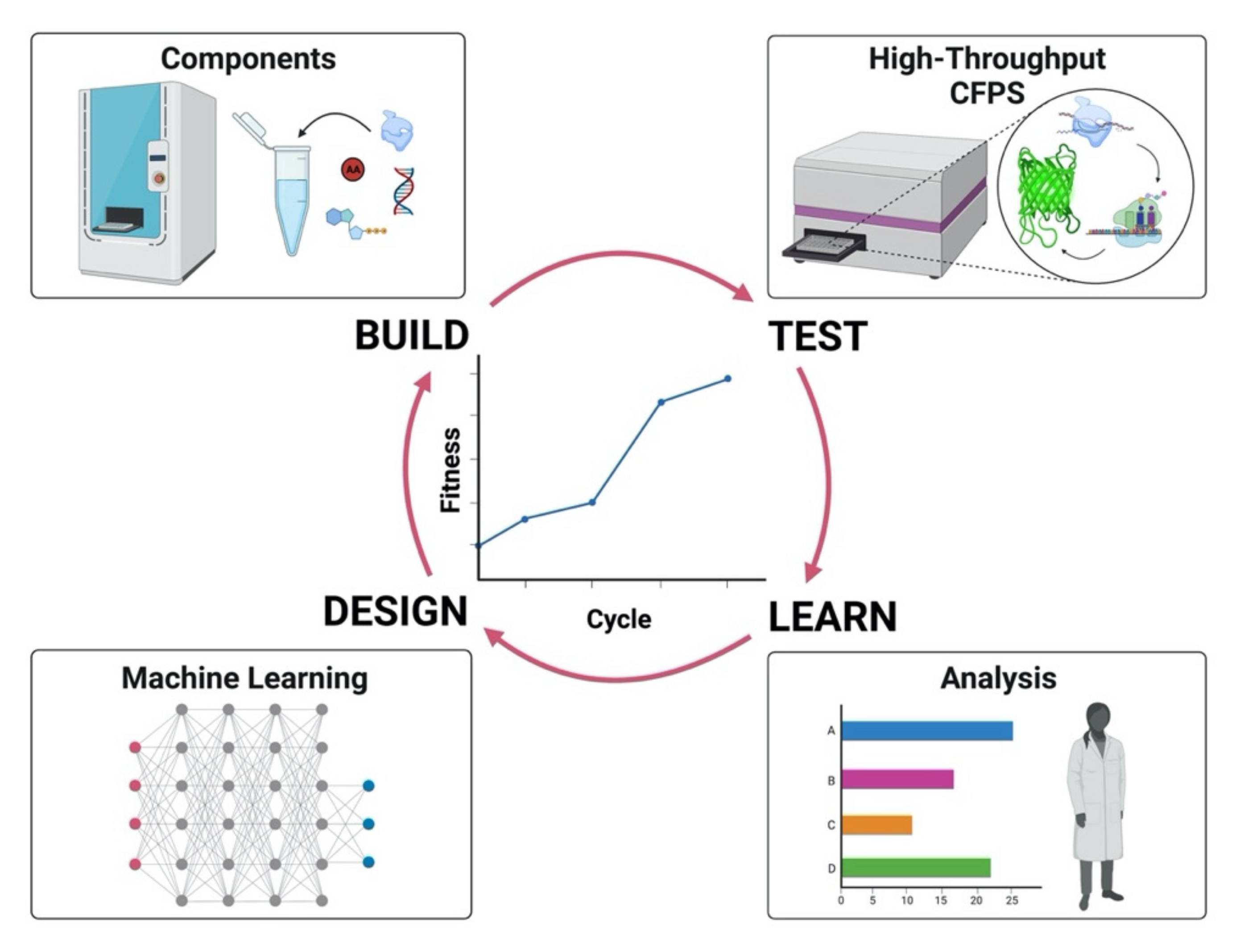
7. Utilizing Machine Learning in Cell-Free
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Koo, J.; Yang, J.; Park, H. Cell-free Systems: Recent Advances and Future Outlook. Biotechnol. Bioprocess. Eng. 2020, 25, 955–961. [Google Scholar] [CrossRef]
- Buchner, E. Alcoholic Fermentation without Yeast Cells. Rep. Ger. Chem. Soc. Berl. 1897, 1, 117–124. [Google Scholar]
- Eduard Buchner Nobel Lecture. Available online: https://www.nobelprize.org/prizes/chemistry/1907/buchner/lecture/ (accessed on 28 June 2021).
- Chong, S. Overview of Cell-Free Protein Synthesis: Historic Landmarks, Commercial Systems, and Expanding Applications. Curr. Protoc. Mol. Biol. 2014, 108, 16–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamecnik, P.C.; Frantz, I.D.; Loftfield, R.B.; Stephenson, M.L. Incorporation in vitro of radioactive carbon from carboxyl-labeled dl-alanine and glycine into proteins of normal and malignant rat livers. J. Biol. Chem. 1948, 175, 299–314. [Google Scholar] [CrossRef]
- Gale, E.F.; Folkes, J.P. Effect of Nucleic Acids on Protein Synthesis and Amino-Acid Incorporation in Disrupted Staphylococcal Cells. Nature 1954, 173, 1223–1227. [Google Scholar] [CrossRef]
- Lamborg, M.R.; Zamecnik, P.C. Amino acid incorporation into protein by extracts of E. coli. Biochim. Biophys. Acta 1960, 42, 206–211. [Google Scholar] [CrossRef]
- Bank, A.; Marks, P.A. Protein synthesis in a cell free human reticulocyte system: Ribosome function in thalassemia. J. Clin. Investig. 1966, 45, 330–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.L.; Ivashkiv, L.; Chen, H.Z.; Zubay, G.; Cashel, M. Cell-free coupled transcription-translation system for investigation of linear DNA segments. Proc. Natl. Acad. Sci. USA 1980, 77, 7029–7033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spirin, A.S.; Baranov, V.I.; Ryabova, L.A.; Ovodov, S.Y.; Alakhov, Y.B. A continuous cell-free translation system capable of producing polypeptides in high yield. Science 1988, 242, 1162–1164. [Google Scholar] [CrossRef]
- Gesteland, R.F. Isolation and characterization of ribonuclease I mutants of Escherichia coli. J. Mol. Biol. 1966, 16, 67–84. [Google Scholar] [CrossRef]
- Zhou, Y.; Asahara, H.; Gaucher, E.A.; Chong, S. Reconstitution of translation from Thermus thermophilus reveals a minimal set of components sufficient for protein synthesis at high temperatures and functional conservation of modern and ancient translation components. Nucleic Acids Res. 2012, 40, 7932–7945. [Google Scholar] [CrossRef] [Green Version]
- Michel-Reydellet, N.; Calhoun, K.; Swartz, J. Amino acid stabilization for cell-free protein synthesis by modification of the Escherichia coli genome. Metab. Eng. 2004, 6, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Jewett, M.C.; Calhoun, K.A.; Voloshin, A.; Wuu, J.J.; Swartz, J.R. An integrated cell-free metabolic platform for protein production and synthetic biology. Mol. Syst. Biol. 2008, 4, 220. [Google Scholar] [CrossRef]
- Shimizu, Y.; Inoue, A.; Tomari, Y.; Suzuki, T.; Yokogawa, T.; Nishikawa, K.; Ueda, T. Cell-free translation reconstituted with purified components. Nat. Biotechnol. 2001, 19, 751–755. [Google Scholar] [CrossRef]
- Carlson, E.D.; Gan, R.; Hodgman, C.E.; Jewett, M.C. Cell-free protein synthesis: Applications come of age. Biotechnol. Adv. 2012, 30, 1185–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzen, F.; Chang, G.; Kudlicki, W. The past, present and future of cell-free protein synthesis. Trends Biotechnol. 2005, 23, 150–156. [Google Scholar] [CrossRef]
- Kwok, R. Five hard truths for synthetic biology. Nature 2010, 463, 288–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, M.Z.; Gregorio, N.E.; Jewett, M.C.; Watts, K.R.; Oza, J.P. Escherichia coli-Based Cell-Free Protein Synthesis: Protocols for a robust, flexible, and accessible platform technology. JoVE 2019, 144, e58882. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-Q.; Wu, C.; Jewett, M.C.; Li, J. Cell-Free Protein Synthesis Enables One-Pot Cascade Biotransformation in an Aqueous-Organic Biphasic System. Biotechnol. Bioeng. 2020, 117, 4001–4008. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Faulon, J.-L.; Borkowski, O. Models for Cell-Free Synthetic Biology: Make Prototyping Easier, Better, and Faster. Front. Bioeng. Biotechnol. 2018, 6, 182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Guo, W.; Lu, Y. Advances in Cell-Free Biosensors: Principle, Mechanism, and Applications. Biotechnol. J. 2020, 15, 2000187. [Google Scholar] [CrossRef] [PubMed]
- Karig, D.K. Cell-free synthetic biology for environmental sensing and remediation. Curr. Opin. Biotechnol. 2017, 45, 69–75. [Google Scholar] [CrossRef]
- Zawada, J.F.; Yin, G.; Steiner, A.R.; Yang, J.; Naresh, A.; Roy, S.M.; Gold, D.S.; Heinsohn, H.G.; Murray, C.J. Microscale to manufacturing scale-up of cell-free cytokine production—A new approach for shortening protein production development timelines. Biotechnol. Bioeng. 2011, 108, 1570–1578. [Google Scholar] [CrossRef] [Green Version]
- Rasor, B.J.; Vögeli, B.; Landwehr, G.M.; Bogart, J.W.; Karim, A.S.; Jewett, M.C. Toward sustainable, cell-free biomanufacturing. Curr. Opin. Biotechnol. 2021, 69, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liu, Y.; Liu, W.-Q.; Wu, C.; Li, J. Designing modular cell-free systems for tunable biotransformation of l-phenylalanine to aromatic compounds. Front. Bioeng. Biotechnol. 2021, 9, 688. [Google Scholar] [CrossRef]
- Chiba, C.H.; Knirsch, M.C.; Azzoni, A.R.; Moreira, A.R.; Stephano, M.A. Cell-free protein synthesis: Advances on production process for biopharmaceuticals and immunobiological products. Biotechniques 2021, 70, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Beebe, E.T.; Makino, S.; Markley, J.L.; Fox, B.G. Automated Cell-Free Protein Production Methods for Structural Studies. Methods Mol. Biol. 2014, 1140, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Caschera, F.; Bedau, M.A.; Buchanan, A.; Cawse, J.; de Lucrezia, D.; Gazzola, G.; Hanczyc, M.M.; Packard, N.H. Coping with complexity: Machine learning optimization of cell-free protein synthesis. Biotechnol. Bioeng. 2011, 108, 2218–2228. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, J.; Lian, J.; Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synth. Syst. Biotechnol. 2018, 3, 90–96. [Google Scholar] [CrossRef]
- Nirenberg, M.W.; Matthaei, J.H. The dependence of cell-free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proc. Natl. Acad. Sci. USA 1961, 47, 1588–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.D.; Crick, F.H.C. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Crick, F. Central Dogma of Molecular Biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Swartz, J.R. Expanding biological applications using cell-free metabolic engineering: An overview. Metab. Eng. 2018, 50, 156–172. [Google Scholar] [CrossRef] [PubMed]
- McCormick, N.E.; Jakeman, D.L. On the mechanism of phosphoenolpyruvate synthetase (PEPs) and its inhibition by sodium fluoride: Potential magnesium and aluminum fluoride complexes of phosphoryl transfer. Biochem. Cell Biol. 2015, 93, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Dudley, Q.M.; Anderson, K.C.; Jewett, M.C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synth. Biol. 2016, 5, 1578–1588. [Google Scholar] [CrossRef]
- Moore, S.J.; MacDonald, J.T.; Freemont, P.S. Cell-free synthetic biology for in vitro prototype engineering. Biochem. Soc. Trans. 2017, 45, 785–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calhoun, K.A.; Swartz, J.R. Energy Systems for ATP Regeneration in Cell-Free Protein Synthesis Reactions. Vitr. Transcr. Transl. Protoc. 2007, 3–17. [Google Scholar] [CrossRef]
- Jensen, A.B.; Hubálek, F.; Stidsen, C.E.; Johansson, E.; Öberg, F.K.; Skjøt, M.; Kjeldsen, T. Cell free protein synthesis versus yeast expression—A comparison using insulin as a model protein. Protein Expr. Pur. 2021, 186, 105910. [Google Scholar] [CrossRef] [PubMed]
- Hartley, C.J.; Williams, C.C.; Scoble, J.A.; Churches, Q.I.; North, A.; French, N.G.; Nebl, T.; Coia, G.; Warden, A.C.; Simpson, G.; et al. Engineered enzymes that retain and regenerate their cofactors enable continuous-flow biocatalysis. Nat. Catal. 2019, 2, 1006–1015. [Google Scholar] [CrossRef]
- Kwon, Y.-C.; Jewett, M.C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Sci. Rep. 2015, 5, 8663. [Google Scholar] [CrossRef] [Green Version]
- Garamella, J.; Marshall, R.; Rustad, M.; Noireaux, V. The all, E. coli TX-TL Toolbox 2.0: A Platform for Cell-Free Synthetic Biology. ACS Synth. Biol. 2016, 5, 344–355. [Google Scholar] [CrossRef]
- Caschera, F.; Noireaux, V. Synthesis of 2.3 mg/mL of protein with an all Escherichia coli cell-free transcription-translation system. Biochimie 2014, 99, 162–168. [Google Scholar] [CrossRef]
- Garenne, D.; Thompson, S.; Brisson, A.; Khakimzhan, A.; Noireaux, V. The all-E. coli TXTL toolbox 3.0: New capabilities of a cell-free synthetic biology platform. Synth. Biol. 2021, 6, ysab017. [Google Scholar] [CrossRef] [PubMed]
- Caschera, F.; Noireaux, V. A cost-effective polyphosphate-based metabolism fuels an all E. coli cell-free expression system. Metab. Eng. 2015, 27, 29–37. [Google Scholar] [CrossRef]
- Lim, H.J.; Kim, D.-M. Cell-Free Metabolic Engineering: Recent Developments and Future Prospects. Methods Protoc. 2019, 2, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, A.S.; Dudley, Q.M.; Juminaga, A.; Yuan, Y.; Crowe, S.A.; Heggestad, J.T.; Garg, S.; Abdalla, T.; Grubbe, W.S.; Rasor, B.J.; et al. In vitro prototyping and rapid optimization of biosynthetic enzymes for cell design. Nat. Chem. Biol. 2020, 16, 912–919. [Google Scholar] [CrossRef]
- Karim, A.S.; Jewett, M.C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metab. Eng. 2016, 36, 116–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogart, J.W.; Cabezas, M.D.; Vögeli, B.; Wong, D.A.; Karim, A.S.; Jewett, M.C. Cell-Free Exploration of the Natural Product Chemical Space. ChemBioChem 2021, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.; Maxwell, C.S.; Collins, S.P.; Jacobsen, T.; Luo, M.L.; Begemann, M.B.; Gray, B.N.; January, E.; Singer, A.; He, Y. Rapid and Scalable Characterization of CRISPR Technologies Using an E. coli Cell-Free Transcription-Translation System. Mol. Cell 2018, 69, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.Z.; Yeung, E.; Hayes, C.A.; Noireaux, V.; Murray, R.M. Linear DNA for Rapid Prototyping of Synthetic Biological Circuits in an Escherichia coli Based TX-TL Cell-Free System. ACS Synth. Biol. 2013, 3, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Estrela, P.; Bhalla, N.; Jolly, P.; Formisano, N.; Estrela, P. Introduction to biosensors. Essays Biochem. 2016, 60, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beabout, K.; Bernhards, C.B.; Thakur, M.; Turner, K.B.; Cole, S.D.; Walper, S.A.; Chávez, J.L.; Lux, M.W. Optimization of heavy metal sensors based on transcription factors and cell-free expression systems. ACS Synth. Biol. 2021, 10, 3040–3054. [Google Scholar] [CrossRef] [PubMed]
- Arce, A.; Chavez, F.G.; Gandini, C.; Puig, J.; Matute, T.; Haseloff, J.; Dalchau, N.; Molloy, J.; Pardee, K.; Federici, F. Decentralizing Cell-Free RNA Sensing with the Use of Low-Cost Cell Extracts. Front. Bioeng. Biotechnol. 2021, 9, 727584. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Wang, M. Design, Optimization and Application of Small Molecule Biosensor in Metabolic Engineering. Front. Microbiol. 2017, 8, 2012. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Fong, S.S. Biosensors for Metabolic Engineering. Synth. Syst. Biotechnol. 2016, 53–70. [Google Scholar] [CrossRef]
- Brophy, K.; Davies, S.; Olenik, S.; Çotur, Y.; Ming, D.; Van Zalk, N.; Yetisen, A.K. The Future of Wearable Technologies. ICL 2021, 6, 1–20. [Google Scholar] [CrossRef]
- Nguyen, P.Q.; Soenksen, L.R.; Donghia, N.M.; Angenent-Mari, N.M.; de Puig, H.; Huang, A.; Lee, R.; Slomovic, S.; Galbersanini, T.; Lansberry, G.; et al. Wearable materials with embedded synthetic biology sensors for biomolecule detection. Nat. Biotechnol. 2021, 39, 1366–1374. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [Green Version]
- English, M.A.; Soenksen, L.R.; Gayet, R.V.; de Puig, H.; Angenent-Mari, N.M.; Mao, A.S.; Nguyen, P.Q.; Collins, J.J. Programmable CRISPR-responsive smart materials. Science 2019, 365, 780–785. [Google Scholar] [CrossRef]
- Huang, A.; Nguyen, P.Q.; Stark, J.C.; Takahashi, M.K.; Donghia, N.; Ferrante, T.; Dy, A.J.; Hsu, K.J.; Dubner, R.S.; Pardee, K.; et al. BiobitsTM explorer: A modular synthetic biology education kit. Sci. Adv. 2018, 4, 5105–5106. [Google Scholar] [CrossRef] [Green Version]
- Sharpes, C.E.; McManus, J.B.; Blum, S.M.; Mgboji, G.E.; Lux, M.W. Assessment of colorimetric reporter enzymes in the pure system. ACS Synth. Biol. 2021, 10, 3205–3208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.P.; Sun, J.; Ma, Y. Biomanufacturing: History and perspective. J. Ind. Microbiol. Biotechnol. 2017, 44, 773–784. [Google Scholar] [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nunez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.-S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [Green Version]
- Tamang, J.P.; Cotter, P.D.; Endo, A.; Han, N.S.; Kort, R.; Liu, S.Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented foods in a global age: East meets West. Compr. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, A.E. Elmer, L. Gaden, Jr., Father of biochemical engineering. Biotechnol. Bioeng. 1991, 37, 995–997. [Google Scholar] [CrossRef]
- Goto, M.; Akai, K.; Murakami, A.; Hashimoto, C.; Tsuda, E.; Ueda, M.; Kawanishi, G.; Takahashi, N.; Ishimoto, A.; Chiba, H.; et al. Production of Recombinant Human Erythropoietin in Mammalian Cells: Host–Cell Dependency of the Biological Activity of the Cloned Glycoprotein. Bio/Technology 1988, 6, 67–71. [Google Scholar] [CrossRef]
- Leniobio. Available online: https://www.leniobio.com/technology/ (accessed on 17 October 2021).
- Swiftscale Biologics. Available online: https://www.swiftscalebio.com/our-approach (accessed on 17 October 2021).
- Kykeon Biotech. Available online: https://kykeonbiotech.de/en/about/ (accessed on 17 October 2021).
- Technology—Cell Free Sciences. Available online: https://www.cfsciences.com/eg/ (accessed on 17 October 2021).
- Bundy, B.C.; Hunt, J.P.; Jewett, M.; Swartz, J.R.; Wood, D.W.; Frey, D.D.; Rao, G. Cell-free biomanufacturing. Curr. Opin. Chem. Eng. 2018, 22, 177–183. [Google Scholar] [CrossRef]
- O’Flaherty, R.; Bergin, A.; Flampouri, E.; Mota, L.M.; Obaidi, I.; Quigley, A.; Xie, Y.; Butler, M. Mammalian cell culture for production of recombinant proteins: A review of the critical steps in their biomanufacturing. Biotechnol. Adv. 2020, 43, 107552. [Google Scholar] [CrossRef]
- González Peña, O.I.; López Zavala, M.Á.; Cabral Ruelas, H. Pharmaceuticals Market, Consumption Trends and Disease Incidence Are Not Driving the Pharmaceutical Research on Water and Wastewater. Int. J. Environ. Res. Public Health 2021, 18, 2532. [Google Scholar] [CrossRef] [PubMed]
- Kitney, R.I.; Bell, J.; Philp, J. Build a sustainable vaccines industry with synthetic biology. Trends Biotechnol. 2021, 39, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, C.W. New 2016 data and statistics for global pharmaceutical products and projections through 2017. ACS Chem. Neurosci. 2017, 8, 1635–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, S.; Riminton, S. Monoclonal antibody therapy for non-malignant disease. Aust. Prescr. 2006, 29, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Drago, J.Z.; Modi, S.; Chandarlapaty, S. Unlocking the potential of antibody–drug conjugates for cancer therapy. Nat. Rev. Clin. Oncol. 2021, 18, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment. Int. J. Oncol. 2018, 54, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Walko, C.M.; West, H. Antibody drug conjugates for cancer treatment. JAMA Oncol. 2019, 5, 1648. [Google Scholar] [CrossRef] [Green Version]
- Rajewsky, K. The advent and rise of monoclonal antibodies. Nature 2019, 575, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Thaore, V.; Tsourapas, D.; Shah, N.; Kontoravdi, C. Techno-Economic Assessment of Cell-Free Synthesis of Monoclonal Antibodies Using CHO Cell Extracts. Processes 2020, 8, 454. [Google Scholar] [CrossRef]
- Stech, M.; Kubick, S. Cell-Free Synthesis Meets Antibody Production: A Review. Antibodies 2015, 4, 12–33. [Google Scholar] [CrossRef] [Green Version]
- Stech, M.; Nikolaeva, O.; Thoring, L.; Stöcklein, W.F.M.; Wüstenhagen, D.A.; Hust, M.; Dübel, S.; Kubick, S. Cell-free synthesis of functional antibodies using a coupled in vitro transcription-translation system based on CHO cell lysates. Sci. Rep. 2017, 7, 12030. [Google Scholar] [CrossRef] [Green Version]
- Merk, H.; Stiege, W.; Tsumoto, K.; Kumagai, I.; Erdmann, V.A. Cell-Free Expression of Two Single-Chain Monoclonal Antibodies against Lysozyme: Effect of Domain Arrangement on the Expression. J. Biochem. 1999, 125, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Sutro Biopharma. Available online: https://www.sutrobio.com/about-us (accessed on 21 October 2021).
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 2559. [Google Scholar] [CrossRef] [PubMed]
- Guilhelmelli, F.; Vilela, N.; Albuquerque, P.; Derengowski, L.; Silva-Pereira, I.; Kyaw, C. Antibiotic development challenges: The various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front. Microbiol. 2013, 4, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef]
- Lee, K.H.; Kwon, Y.C.; Yoo, S.J.; Kim, D.M. Ribosomal synthesis and in situ isolation of peptide molecules in a cell-free translation system. Protein Expr. Purif. 2010, 71, 16–20. [Google Scholar] [CrossRef]
- Pardee, K.; Slomovic, S.; Nguyen, P.; Lee, J.W.; Donghia, N.; Burrill, D.; Ferrante, T.; McSorley, F.R.; Furuta, Y.; Vernet, A.; et al. Portable, On-Demand Biomolecular Manufacturing. Cell 2016, 167, 248–259.e12. [Google Scholar] [CrossRef] [Green Version]
- Wang, G. Human Antimicrobial Peptides and Proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2020, 21, 83–100. [Google Scholar] [CrossRef]
- Maruggi, G.; Zhang, C.; Li, J.; Ulmer, J.B.; Yu, D. Mrna as a transformative technology for vaccine development to control infectious diseases. Mol. Ther. 2019, 27, 757–772. [Google Scholar] [CrossRef] [Green Version]
- De Sá Magalhães, S.; Keshavarz-Moore, E. Pichia pastoris (Komagataella phaffii) as a cost-effective tool for vaccine production for low- and middle-income countries(Lmics). Bioengineering 2021, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Gomez, P.L.; Robinson, J.M. Vaccine Manufacturing. Plotkin’s Vaccines 2018, 51-60.e1. [Google Scholar] [CrossRef]
- Stark, J.C.; Jaroentomeechai, T.; Moeller, T.D.; Hershewe, J.M.; Warfel, K.F.; Moricz, B.S.; Martini, A.M.; Dubner, R.S.; Hsu, K.J.; Stevenson, T.C.; et al. On-demand biomanufacturing of protective conjugate vaccines. Sci. Adv. 2021, 7, eabe9444. [Google Scholar] [CrossRef]
- CDC. Available online: https://www.cdc.gov/coronavirus/2019-ncov/vaccines/different-vaccines/mrna.html (accessed on 28 June 2021).
- Path. Available online: https://www.path.org/articles/mrna-and-future-vaccine-manufacturing/ (accessed on 28 June 2021).
- Adamo, A.; Beingessner, R.L.; Behnam, M.; Chen, J.; Jamison, T.F.; Jensen, K.F.; Monbaliu, J.-C.M.; Myerson, A.S.; Revalor, E.M.; Snead, D.R.; et al. On-demand continuous-flow production of pharmaceuticals in a compact, reconfigurable system. Science 2016, 352, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pinera, P.; Han, N.; Cleto, S.; Cao, J.; Purcell, O.; Shah, K.A.; Lee, K.; Ram, R.; Lu, T.K. Synthetic biology and microbioreactor platforms for programmable production of biologics at the point-of-care. Nat. Commun. 2016, 7, 12211. [Google Scholar] [CrossRef] [PubMed]
- The Medicine Maker. Available online: https://themedicinemaker.com/manufacture/why-small-molecules-are-still-a-big-deal (accessed on 4 September 2021).
- Tinafar, A.; Jaenes, K.; Pardee, K. Synthetic Biology Goes Cell-Free. BMC Biology 2019, 17, 64. [Google Scholar] [CrossRef] [Green Version]
- Kay, J.E.; Jewett, M.C. A cell-free system for production of 2,3-butanediol is robust to growth-toxic compounds. Metab. Eng. Commun. 2020, 10, e00114. [Google Scholar] [CrossRef] [PubMed]
- Design Pharmaceuticals. Available online: https://www.designpharmaceuticals.com/technology/our-technology.html (accessed on 21 October 2021).
- Yin, H.; Flynn, A.D. Drugging Membrane Protein Interactions. Annu. Rev. Biomed. Eng. 2016, 18, 51–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoring, L.; Wüstenhagen, D.A.; Borowiak, M.; Stech, M.; Sonnabend, A.; Kubick, S. Cell-Free Systems Based on CHO Cell Lysates: Optimization Strategies, Synthesis of “Difficult-to-Express” Proteins and Future Perspectives. PLoS ONE 2016, 11, e0163670. [Google Scholar] [CrossRef]
- Schneider, B.; Junge, F.; Shirokov, V.A.; Durst, F.; Schwarz, D.; Dötsch, V.; Bernhard, F. Membrane protein expression in cell-free systems. Methods Mol. Biol. 2010, 601, 165–186. [Google Scholar] [CrossRef]
- Synthelis. Available online: https://www.synthelis.com/ (accessed on 18 October 2021).
- Henrich, E.; Hein, C.; Dötsch, V.; Bernhard, F. Membrane protein production in Escherichia coli cell-free lysates. FEBS Letters 2015, 589, 1713–1722. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Lin, X.; Lu, Y.; Zhang, J. Flexible on-demand cell-free protein synthesis platform based on a tube-in-tube reactor. React. Chem. Eng. 2020, 5, 270–277. [Google Scholar] [CrossRef]
- Rao, G.; Kostov, Y.; Tolosa, L.; Ge, X.; Frey, D. System and Method for Production of On-Demand Proteins in a Portable Unit for Point of Care Delivery. U.S. Patent 10435664B2, 10 July 2019. [Google Scholar]
- Adiga, R.; Andar, A.; Borhani, S.; Burgenson, D.; Deldari, S.; Frey, D.; Ge, X.; Gopalakrishnan, M.; Gurramkonda, C.; Gutierrez, E.; et al. Manufacturing biological medicines on demand: Safety and efficacy of granulocyte colony-stimulating factor in a mouse model of total body irradiation. Biotechnol. Prog. 2020, 36, e2970. [Google Scholar] [CrossRef]
- Boles, K.S.; Kannan, K.; Gill, J.; Felderman, M.; Gouvis, H.; Hubby, B.; I Kamrud, K.; Venter, J.C.; Gibson, D.G. Digital-to-biological converter for on-demand production of biologics. Nat. Biotechnol. 2017, 35, 672–675. [Google Scholar] [CrossRef]
- Nuclera. Available online: https://www.nuclera.com/ (accessed on 17 October 2021).
- Liberumbio. Available online: http://www.liberumbio.com/ (accessed on 17 October 2021).
- Tierra Biosciences. Available online: https://www.tierrabiosciences.com/ (accessed on 17 October 2021).
- Eisenstein, M. Enzymatic DNA synthesis enters new phase. Nat. Biotechnol. 2020, 38, 1113–1115. [Google Scholar] [CrossRef]
- Jebrail, M.J.; Bartsch, M.S.; Patel, K.D. Digital microfluidics: A versatile tool for applications in chemistry, biology and medicine. Lab. A Chip 2012, 12, 2452–2463. [Google Scholar] [CrossRef] [PubMed]
- Garenne, D.; Haines, M.C.; Romantseva, E.F.; Freemont, P.; Strychalski, E.A.; Noireaux, V. Cell-free gene expression. Nat. Rev. Methods Primers 2021, 1, 1–18. [Google Scholar] [CrossRef]
- Silverman, A.D.; Karim, A.S.; Jewett, M.C. Cell-free gene expression: An expanded repertoire of applications. Nat. Rev. Genet. 2020, 21, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Sethuraman, N.; Stadheim, T.A. Challenges in therapeutic glycoprotein production. Curr. Opin. Biotechnol. 2006, 17, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Popa, M.E.; Mitelut, A.C.; Popa, E.E.; Matei, F. Creating products and services in food biotechnology. In Introduction to Biotech Entrepreneurship: From Idea to Business: A European Perspective; Matei, F., Zirra, D., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 141–178. [Google Scholar] [CrossRef]
- Claassens, N.J.; Burgener, S.; Vögeli, B.; Erb, T.J.; Bar-Even, A. A critical comparison of cellular and cell-free bioproduction systems. Curr. Opin. Biotechnol. 2019, 60, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, C.; Zhao, Z.; Xu, J.; Li, J.; Li, P. Application of cell-free protein synthesis system for the biosynthesis of l-theanine. ACS Synth. Biol. 2021, 10, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Müller-Maatsch, J.; Gras, C. The “Carmine Problem” and Potential Alternatives. Handb. Nat. Pigment. Food Beverages 2016, 18, 385–428. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Debut Biotech. Available online: https://debutbiotech.com/biomanufacturing/ (accessed on 4 September 2021).
- Ruhela, M.; Nagar, L.; Gupta, A.; Popli, H. Cosmetics: Regulatory and Market Scenario for US and India. Pharma Innov. J. 2018, 7, 164–169. [Google Scholar]
- UCLA Technology Development Group. Available online: https://techtransfer.universityofcalifornia.edu/NCD/23596.html (accessed on 28 June 2021).
- Secchi, G. Role of protein in cosmetics. Clin. Dermatol. 2008, 26, 321–325. [Google Scholar] [CrossRef]
- Diegelmann, R.F.; Bernstein, L.; Peterkofsky, B. Cell-free Collagen Synthesis on Membrane-bound Polysomes Chick Embryo Connective Tissue and the Localization of Prolyl Hydroxylase on the Polysome-Membrane Complex*. JBC 1973, 248, 6514–6521. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef] [Green Version]
- Rigoldi, F.; Donini, S.; Redaelli, A.; Parisini, E.; Gautieri, A. Review: Engineering of thermostable enzymes for industrial applications. APL Bioeng. 2018, 2, 011501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelwick, R.; Webb, A.J.; MacDonald, J.T.; Freemont, P.S. Development of a Bacillus subtilis cell-free transcription-translation system for prototyping regulatory elements. Metab. Eng. 2016, 38, 370–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowbray, M.; Savage, T.; Wu, C.; Song, Z.; Cho, B.A.; Del Rio-Chanona, E.A.; Zhang, D. Machine learning for biochemical engineering: A review. Biochem. Eng. J. 2021, 172, 108054. [Google Scholar] [CrossRef]
- Vamathevan, J.; Clark, D.; Czodrowski, P.; Dunham, I.; Ferran, E.; Lee, G.; Li, B.; Madabhushi, A.; Shah, P.; Spitzer, M.; et al. Applications of machine learning in drug discovery and development. Nat. Rev. Drug Discov. 2019, 18, 463–477. [Google Scholar] [CrossRef]
- Libbrecht, M.W.; Noble, W.S. Machine learning applications in genetics and genomics. Nat. Rev. Genet. 2015, 16, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caschera, F.; Karim, A.S.; Gazzola, G.; d’Aquino, A.E.; Packard, N.H.; Jewett, M.C. High-throughput optimization cycle of a cell-free ribosome assembly and protein synthesis system. ACS Synth. Biol. 2018, 7, 2841–2853. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, O.; Koch, M.; Zettor, A.; Pandi, A.; Batista, A.C.; Soudier, P.; Faulon, J.-L. Large scale active-learning-guided exploration for in vitro protein production optimization. Nat. Commun. 2020, 11, 1872. [Google Scholar] [CrossRef] [Green Version]
- Faulon, J.-L.; Faure, L. In silico, in vitro, and in vivo machine learning in synthetic biology and metabolic engineering. Curr. Opin. Chem. Biol. 2021, 65, 85–92. [Google Scholar] [CrossRef]
- Duran-Villalobos, C.A.; Ogonah, O.; Melinek, B.; Bracewell, D.G.; Hallam, T.; Lennox, B. Multivariate statistical data analysis of cell-free protein synthesis toward monitoring and control. AIChE J. 2021, 67, e17257. [Google Scholar] [CrossRef]
- Colant, N.; Melinek, B.; Teneb, J.; Goldrick, S.; Rosenberg, W.; Frank, S.; Bracewell, D.G. A rational approach to improving titer in Escherichia coli-based cell-free protein synthesis reactions. Biotechnol. Prog. 2021, 37, e3062. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Bickel, F.; Osberghaus, A.; Hubbuch, J. High-throughput characterization of an insect cell-free expression. Eng. Life Sci. 2014, 14, 409–417. [Google Scholar] [CrossRef]
- Gilman, J.; Walls, L.; Bandiera, L.; Menolascina, F. Statistical design of experiments for synthetic biology. ACS Synth. Biol. 2021, 10, 1–18. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brookwell, A.; Oza, J.P.; Caschera, F. Biotechnology Applications of Cell-Free Expression Systems. Life 2021, 11, 1367. https://doi.org/10.3390/life11121367
Brookwell A, Oza JP, Caschera F. Biotechnology Applications of Cell-Free Expression Systems. Life. 2021; 11(12):1367. https://doi.org/10.3390/life11121367
Chicago/Turabian StyleBrookwell, August, Javin P. Oza, and Filippo Caschera. 2021. "Biotechnology Applications of Cell-Free Expression Systems" Life 11, no. 12: 1367. https://doi.org/10.3390/life11121367
APA StyleBrookwell, A., Oza, J. P., & Caschera, F. (2021). Biotechnology Applications of Cell-Free Expression Systems. Life, 11(12), 1367. https://doi.org/10.3390/life11121367