1. Introduction
Soybean cyst nematode (SCN) (
Heterodera glycines Ichinohe) infects soybean (
Glycine max) plantations and hence causes significant financial losses to the soybean production process globally. In China,
H. glycines infection causes an average soybean yield loss of as high as 30%, and the figure even reaches 70% in some areas in Northeast China [
1].
H. glycines infection causes similar yield losses in the USA [
2]. Several methods such as conventional crossbreeding and transgenic breeding have been developed to confer resistance to soybean varieties against
H. glycines [
3]; however, more efficient methods must be developed to effectively combat this pest. Currently, the use of biocontrol agents is a potential pest-control method, which involves coating seeds with microorganisms to protect plants against microbes [
4]. Seed treatment by introducing plant-parasitic nematodes (PPNs) has been extensively used to improve yield by parasitism, antibiosis, paralysis, and lytic enzymes [
1,
5,
6,
7,
8,
9,
10]. Seed treatment indirectly induces different host defense pathways such as salicylic acid (SA), jasmonic acid (JA), auxin, cytokinin, and reactive oxygen species (ROS) signaling and chemical defense components through the synthesis of plant secondary metabolites or different enzymes to facilitate their parasitic successes in plants, which constitutes an advantage for the plant growth [
11].
In the present study, we attempted to improve the root resistance of soybean against
H. glycines through seed coating.
H. glycines infection in soybean roots could be prevented by stimulating the roots with some active compounds isolated from microorganisms. These active compounds are typically evolutionarily conserved compounds such as saccharides, glycopeptides, lipids or lipopeptides, proteins, and low-molecular-weight metabolites [
12,
13], of which only protein-like or peptide-like plant immune elicitors can regulate plant resistance against nematodes [
14,
15]. The process of inducing plant immunity involves complete utilisation of the potential ability of plants to control disease occurrence. The regulation of plant defence and metabolic systems can induce the basic immune response and delay or reduce the occurrence and development of diseases [
14,
16].
The current research on the types of induced resistance responses to nematodes mainly focuses on two aspects. The first is the microscopic observation of the incompatible interactions between nematodes and their hosts [
17], mainly microscopic observation of the developmental aspects of the infection site (syncytia and giant cells). The second is the identification and observation of rapid allergic reactions. Allergic necrosis is the defense mechanism of plants against a variety of pathogens. Its typical feature is oxidation, which leads to ROS production and the accumulation of phenylpropanoid compounds [
18,
19]. Acibenzolar S-methyl (ASM) is a plant immune elicitor for inducing resistance to PPNs through interference with a pectin lyase during the formation of giant cells affecting the development of nematodes. In addition, after using ASM for five days, the increase in β-1,3-glucanase activity was also an important factor in inducing resistance in plant roots [
20,
21,
22]. The non-protein amino acid β-aminobutyric acid (BABA) inhibits the invasion of rice root-knot nematodes and delays the formation of giant cells and the development of nematodes depending on JA and ET pathways, while activating many basic defense responses, such as the accumulation of ROS, lignin formation, and callose deposition [
23,
24,
25]. Sclareol is an antimicrobial and defense-related substance inhibiting the invasion of Meloidogyne incognita and inducing the expression of related genes such as ET biosynthesis and signal transduction pathways and phenylalanine metabolic pathways. In addition, sclareol activates mitogen-activated protein kinase (MAPK) MAPK3 and MAPK6, which are related to ET biosynthesis [
26]. Silicon-induced defense reactions are related to phenylalanine lyase and lignin, peroxidase, and polyphenol oxidase. The silicon absorbed by plants can be quickly transferred to the roots to prevent the invasion of nematodes by activating the synthesis of phenols, lignin, cork and callosin, which are defense-related substances [
23,
26,
27,
28]. The induced resistance of plants is mainly regulated through the SA or JA/ET pathway. Therefore, exogenous application of SA, JA, or ET and other plant hormones and their analogues is a hot topic in the research of inducing resistance. Foliar spraying of epibrassinolide (BL) can inhibit the biosynthesis of BL in roots and negatively regulate the JA pathway to enhance rice resistance to root-knot nematodes [
29,
30,
31]. However, the active compounds involved in conferring
H. glycines resistance to soybean and the underlying mechanisms of seed treatment involved in stimulating the soybean immune system remain unknown.
In the present study, we aimed to (i) assess the biocontrol capability of Snef1650 towards H. glycines by performing pot and field tests; (ii) isolate and characterise scopoletin obtained from Snef1650; (iii) assess the direct in vitro antagonistic mechanism of scopoletin in juveniles of H. glycines; (iv) assess the biocontrol capability of scopoletin against H. glycines infection by performing field and pot experiments; and (v) confirm whether scopoletin inhibits syncytium formation through seed coating. The study can provide new insight into the utilisation of scopoletin as an anti-H. glycines biocontrol agent.
3. Discussion
In the present study, Snef1650, a potential seed treatment agent for controlling nematodes, was used to improve soybean resistance against
H. glycines. Soybean seeds of susceptible cultivars treated with the Snef1650 fermentation liquid demonstrated increased SCN tolerance and a boost in growth and yield. Studies have demonstrated that microorganism-treated seeds produce nematicides during root development and resist infection due to different parasitic nematodes through systemic or local induction of endogenous hormones and plant defense-related proteins in the plant immune system [
32,
33,
34,
35,
36,
37].
Scoploletin is a type of plant secondary metabolite produced by the phenylalanine metabolism pathway, which is abundantly present in plants and involved in the plant defense response to stress [
38]. Although scopoletin is found in more than 110,000 plants belonging to several families exhibiting direct or indirect resistance against other plant diseases [
10,
15,
39,
40,
41,
42,
43,
44,
45], it had directly detected nematocidal activity and has not yet indirectly induced resistance to nematode in microorganisms. In this study, scoploletin was obtained for the first time through separation and purification of active substances in the fermentation broth of Snef1650. This metabolite was found to be a potent compound and exhibited efficient agent and nematicidal activity in pot experiments and field trials. The presence of scopoletin in microorganisms could be explained by the similarities in enzymes, substrates, and pathways for scopoletin synthesis across various plants and microorganisms. Though the pathways for the biosynthesis of scoploletin exist in different microbes, its extracts outside the plant kingdom with the specific objective of its application to plants against nematodes were explored for the first time in this work. Additionally, the horizontal gene transfer due to long-term coexistence or interaction between microorganisms and plants could be responsible for plant immunity against PPNs and increased soybean yield [
46,
47,
48]. It will be interesting to explore the effect of scopoletin on the root zone dwelling microbes and insects (both pests and friendly ones) in future studies. Additionally, scopoletin might be used as a selection marker for the offspring in breeding
H. glycines-resistant soybean and the possible genetic players of scopoletin metabolism pathway in soybean that might be manipulated for breeding
H. glycines-resistant soybean.
The syncytium is a nurse attendant cell that is positioned inside the soybean roots by
H. glycines.
H. glycines’ survival depends on the development and maintenance of the syncytium. Under nematode stress conditions, resistant soybean cultivars confine the arrangement and advancement of syncytia through several mechanisms during the early phases of soybean–SCN interactions [
49,
50,
51,
52,
53,
54,
55,
56]. Nevertheless, the scopoletin-coating effect on suppressing the development of syncytia in susceptible soybean cultivars has been unclear. Scopoletin treatment slows down syncytium formation, leading to the inhibition of nematode development.
To summarise, both field and pot experiments in this study indicated that soybean seeds treated with scoploletin, obtained from the Snef1650 fermentation broth, exhibited remarkably decreased numbers of H. glycines juveniles and cysts. Scopoletin decreased nematode development in roots and reduced J2 activity in vitro. Additionally, scopoletin significantly promoted the growth of soybean. These results indicate that scopoletin from Penicillium janthinellum Snef1650 has the potential for controlling H. glycines infection.
4. Materials and Methods
4.1. Fungal Strain and Culture Conditions
Snef1650 (Penicillium janthinellum) was chosen from 4600 fungal strains detached from rhizosphere soil test samples, harvested from various regions of China, because of its ability to inhibit H. glycines. Snef1650 was obtained from Northern Nematode Research Institute (NINC), Shenyang Agricultural University, China (CGMCC No. 10487; Patent No.: CN201510607223.9 and CN201810553440.8). Snef1650 was cultured in potato dextrose agar (PDA) at 25 °C for 7 days to prepare the seed-coating agent. The Snef1650 culture plates were punched using a 5-mm diameter punch, and the strain was inoculated into a 50-mL flask for performing a sterilisation test. In an Erlenmeyer flask of liquid fermentation medium, the culture was kept in a shaker incubator (120 rpm) at 25 °C for 8 days. The cultured fermentation broth was filtered through a qualitative filter paper to remove the dense mycelium and then placed in a refrigerator at 4 °C for future use.
4.2. Preparation of Nematodes
H. glycines was isolated from the soil of the NINC experimental field. First, the soil was passed through a set of 420- and 250-μm sieves to collect cysts and was sterilised with 0.5% NaOCl. The surface was sterilised for 3 min, washed thrice with sterile distilled water, and placed in a 25 °C incubator to incubate the second-stage juvenile (J2) prior to collecting J2 (2000 J2 mL−1) for inoculation.
4.3. Biocontrol of H. glycines by Snef1650 in Pot Tests
Jidou 17, a soybean variety susceptible to
H. glycines, was used in all the tests (provided by researcher Zhang Mengchen, Hebei Academy of Agricultural Sciences, China, No. National Examination Bean 2013010). Snef1650 culture (10
9 CFU/mL) was used for the uniform coating of seeds at 70:1 (
w:v), according to the method described by Jing. Z [
57]. At 9 days post-inoculation (dpi), we calculated juvenile quantity within roots according to acid fuchsin staining proposed by Byrd and colleagues [
6]. The amount of juveniles inside the roots, fresh root/shoot weights, and root length were recorded. Ten replicates were set, and this experiment was performed in duplicates.
4.4. Biocontrol of Heterodera Glycines by Snef1650 in Field Tests
Snef1650 fermentation broth was used to coat soybean seeds in May 2016 and 2017 at NINC of Shenyang Agricultural University (123.65bE, 42.32bN). Field trials based on the well-recognised serious SCN infection (>50 cysts/100 mL soil) were performed in the naturally occurring test fields, according to the method described by Jing. Z [
57]. Fresh weight, root/shoot lengths of the above seedlings, juvenile quantity within roots, cyst number on roots, cysts per 100 mL, dry weight of the seedlings, seed/pod quantities in each plant, yield per plot, and 100-seed weight were recorded.
4.5. Isolation, Purification, and Structural Analysis of Scopoletin
The Snef1650 fermentation broth was separated and purified through organic solvent extraction, silica gel column, and semi-preparative HPLC, according to the method described by Xing zhifu [
58]. Snef1650 was cultured on PDA at 25 °C for 7 days, and three pieces of Snef1650 were punched with a 5-mm diameter punch and inoculated with 50 mL of sterile Chagas liquid fermentation medium. The culture was kept in a shaker incubator (120 rpm) at 25 °C. Day-8 culture was used as a seed fermentation broth and was added to a fully automatic mechanically stirred stainless steel fermentation tank for large-scale preparation of 25-L Chac liquid fermentation broth. The Snef1650 fermentation broth was centrifuged in a high-speed centrifuge, evaporated and concentrated, and then mixed with ethanol (1:1,
v/v). The concentrates were separated using a silica gel column through gradient elution with dichloromethane:methanol. A total of 82 tubes were collected. Thin-layer chromatography was performed to combine similar fractions of the concentrates, methane/methanol 90:10 (
v/v), and then they were freeze-dried. Using hydroxamic acid iron reagent, we determined the fourth component as coumarin and further isolated different components and purified them through semi-preparative HPLC under the following chromatographic conditions: semi-preparative chromatography column AQ-C18; mobile phases consisting of 0.1% formic acid aqueous solution (A) and methanol (B) under gradient elution (0–10 min, 85%–66% A; 10–20 min, 66% A); 1.5 mL/min flow rate; 25 °C temperature; 345 nm detection wavelength; and 100 μL injection volume. The corresponding absorption peaks were evaluated and interpreted to obtain compound 1.
To analyse the structure of compound 1 in the purified metabolites of Snef1650, HPLC–MS, and NMR spectroscopy were performed. The chromatographic column for HPLC–MS detection was the Agilent Extend C18 column; other conditions were similar to those for semi-preparative HPLC. The mass spectrometric analysis was performed using AB Sciex QTRAPTM 3200 series quadrupole linear ion trap mass spectrometer (Applied Biosystems, USA). Mass spectrometric analysis conditions were maintained as described by Xing Zhifu [
58]. The structure of compound 1 was detected using NMR spectroscopy (
1H NMR and
13C NMR) (Brook 600 MHz, Karlsruhe, Germany). NMR conditions included a deuterated reagent to dissolve the sample, and the scanned ranges were −2–16 ppm and 0–220 ppm for
1H NMR and
13C NMR, respectively, at 25 °C.
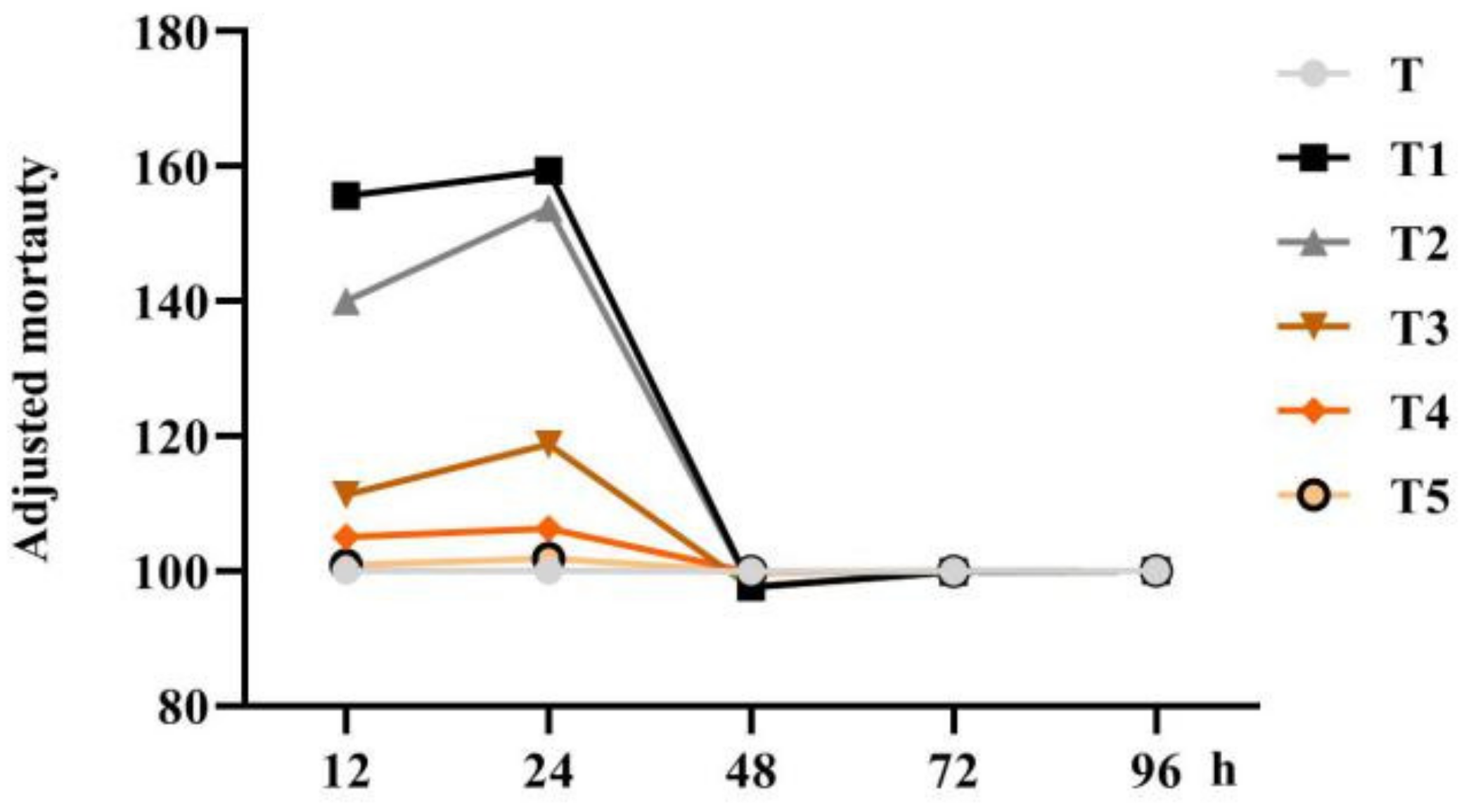
4.6. Effect of Scopoletin on Juveline Mortality In Vitro
An experiment was performed to test scopoletin’s nematicidal properties by adding the second instar larval suspension of the SCN (200 J2/50 L) to the 1-mL sample solution in a 35-mm Petri dish. Nematodes were treated with seven solutions (S, T, T1, T2, T3, T4, and T5). After cultivating the Petri dish containing nematodes in a constant-temperature incubator (25 ± 2 °C) for 12, 24, 48, 72, and 96 h, the second-stage juveniles of live and dead SCNs were counted under an optical microscope (n = 6). The adjusted mortality was calculated according to the following formula: adjusted mortality = (treatment mortality − control mortality)/control mortality × 100 + 100.
Scopoletin was dissolved in 1% methanol to prepare T1 (81.00 μg/mL), T2 (40.50 μg/mL), T3 (20.25 μg/mL), T4 (4.05 μg/mL), and T5 (0.81 μg/mL). In addition, 1% methanol (T) in water (S) served as a control (CK). Fine needles were used to detect deformed, immobile, or stationary juveniles, which were considered dead.
4.7. Biocontrol of H. glycines by Scopoletin in Pot Tests
Seeds were coated with scopoletin in a proportion of approximately 70:1 (w:v). At 3, 6, and 9 dpi, the amount of juveniles inside the roots, root length, fresh root weight, and fresh shoot weight were recorded. The test design reference was 4.3. The optimal concentration of scopoletin was used for field trials.
4.8. Biocontrol of H. glycines by Scopoletin in Field Tests
The optimal concentration of scopoletin was used in pot tests for the field test. Soybean seeds were coated with scopoletin in May 2017 and 2018 at NINC of Shenyang Agricultural University. Scoploletin was applied to soybean seeds in a 70:1 (g/mL) ratio. The test design reference was 4.4. Fresh weight, root/shoot lengths of the above seedlings, juvenile quantity within roots and cyst quantity on roots, cysts per 100 mL, dry weight of the seedlings, pod/seed quantities in each plant, yield per plot, and 100-seed weight were recorded.
4.9. Histological Observation
For observing histological alterations in TJ2 and CKJ2 at 9 dpi, root samples were cut in 1-cm cubes prior to fixation with Carnoy’s fix solution for measuring the syncytia, according to the method described by Lei, P [
55].
4.10. Data Analysis
SPSS17.0 (SPSS, Inc., Chicago, IL, USA) and Microsoft Office Excel 2010 were used for statistical analyses. Analysis of variance (ANOVA), t-test, and Duncan’s multirange test were used to analyse differences among various treatments. A p value of <0.05 was considered to denote statistical significance.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}