
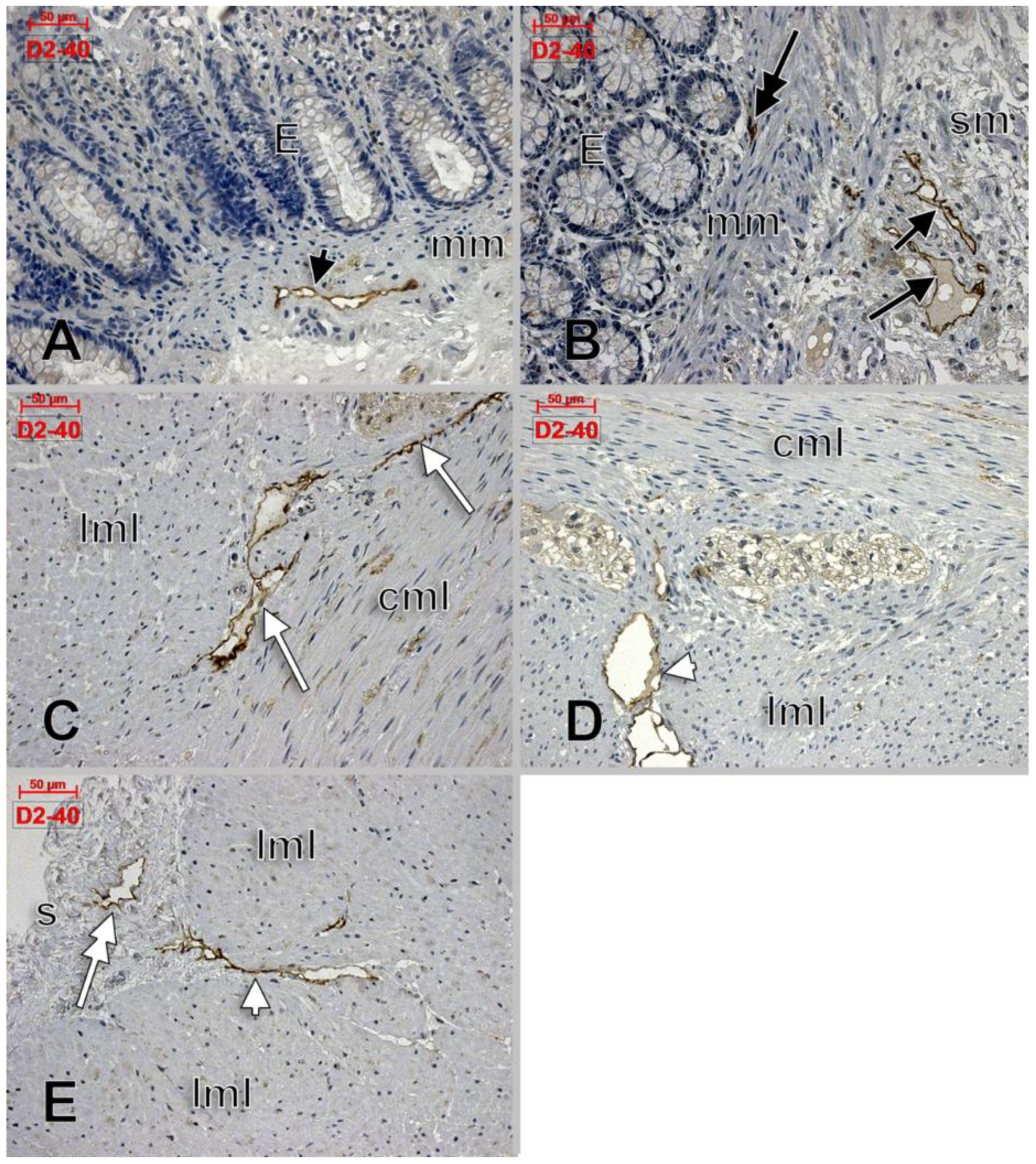
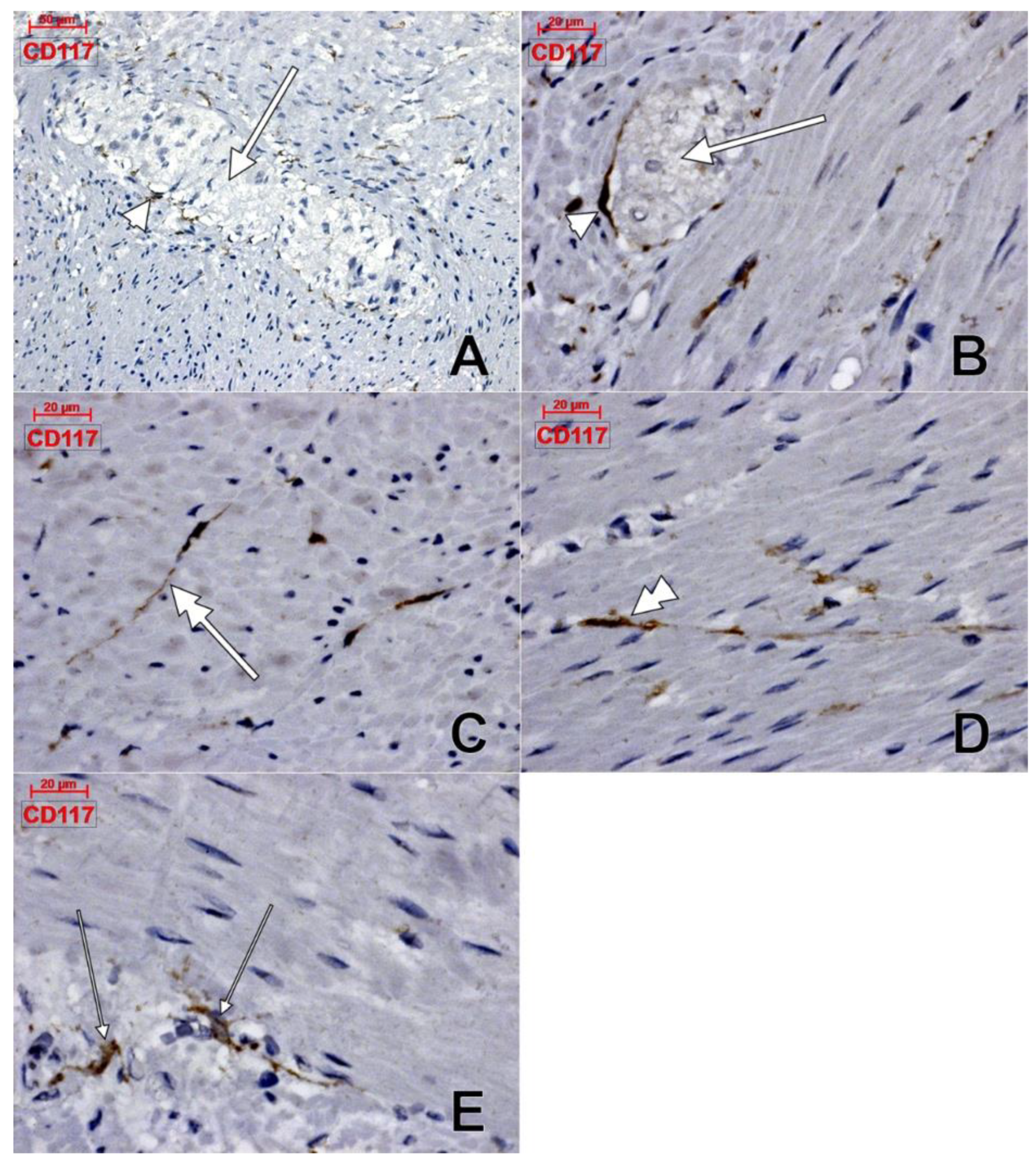
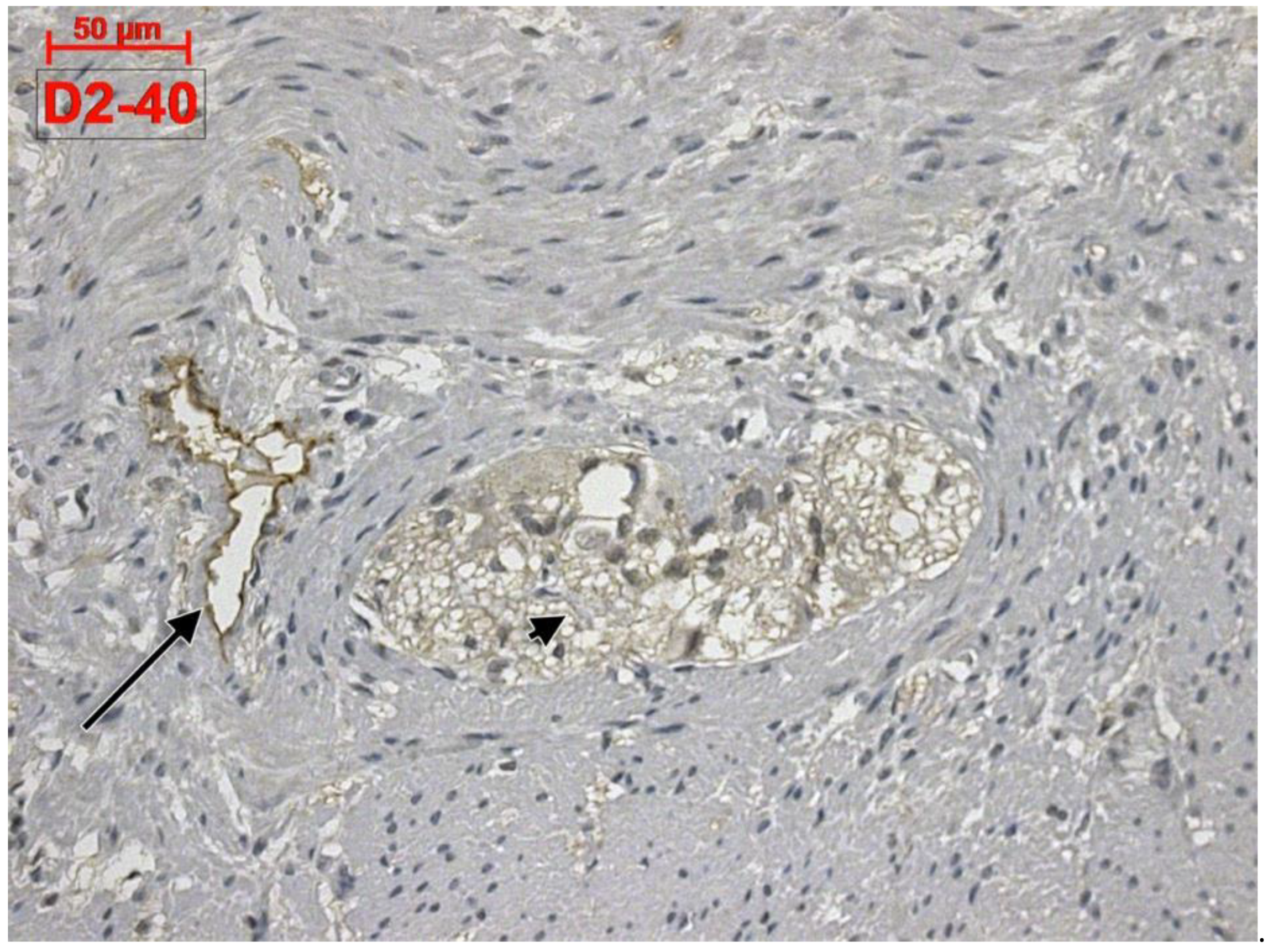
Telocytes and Lymphatics of the Human Colon

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
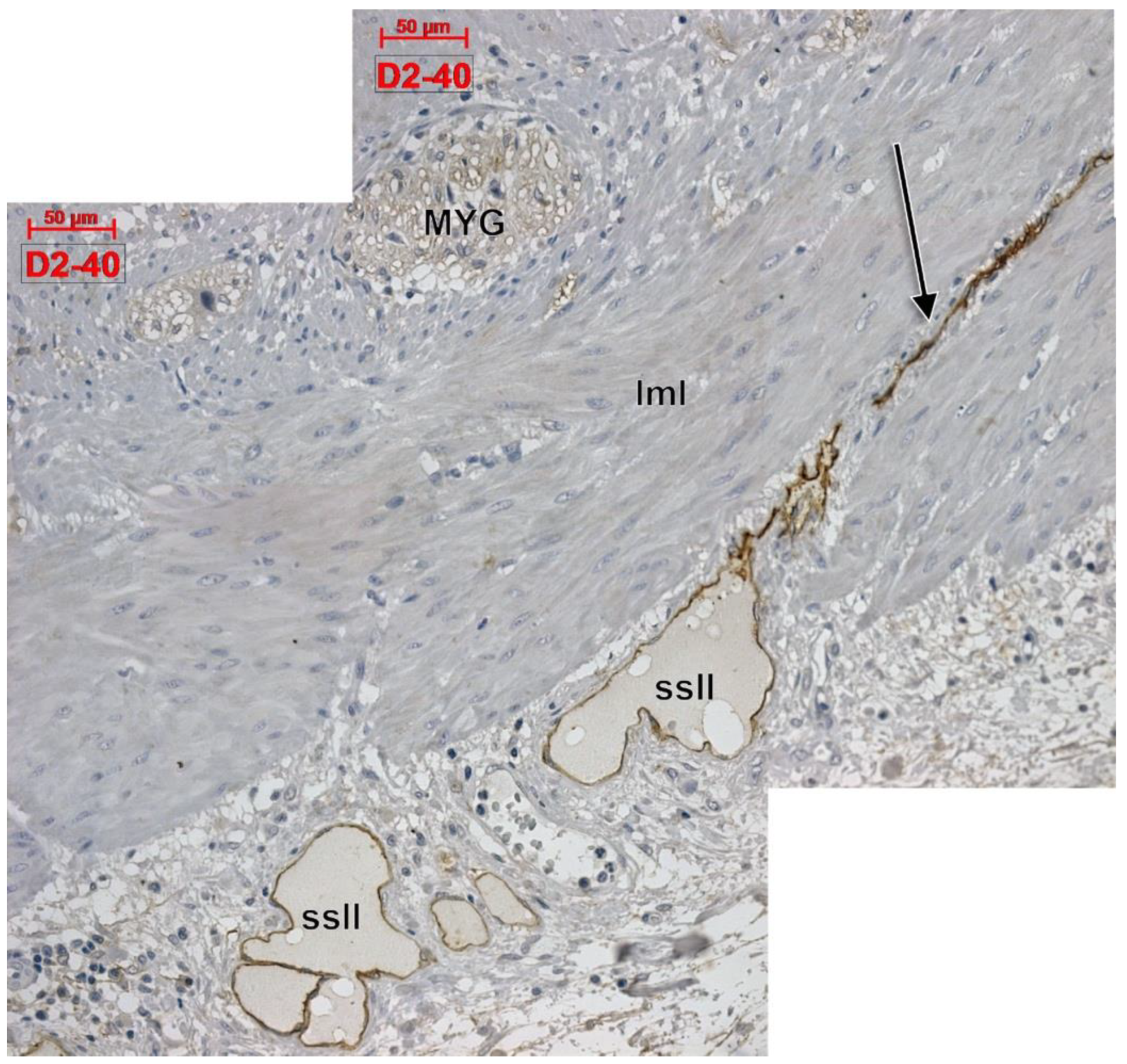
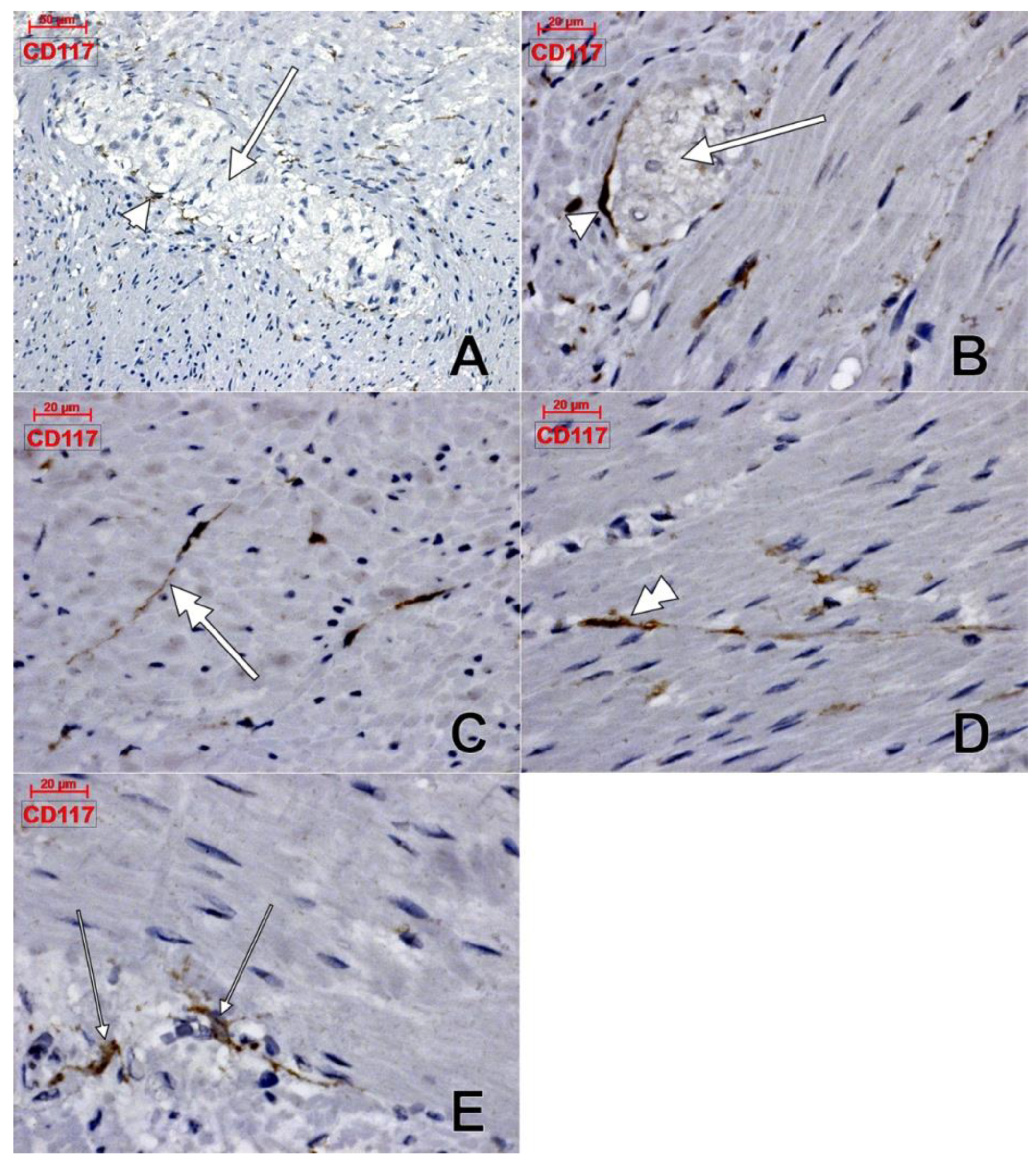
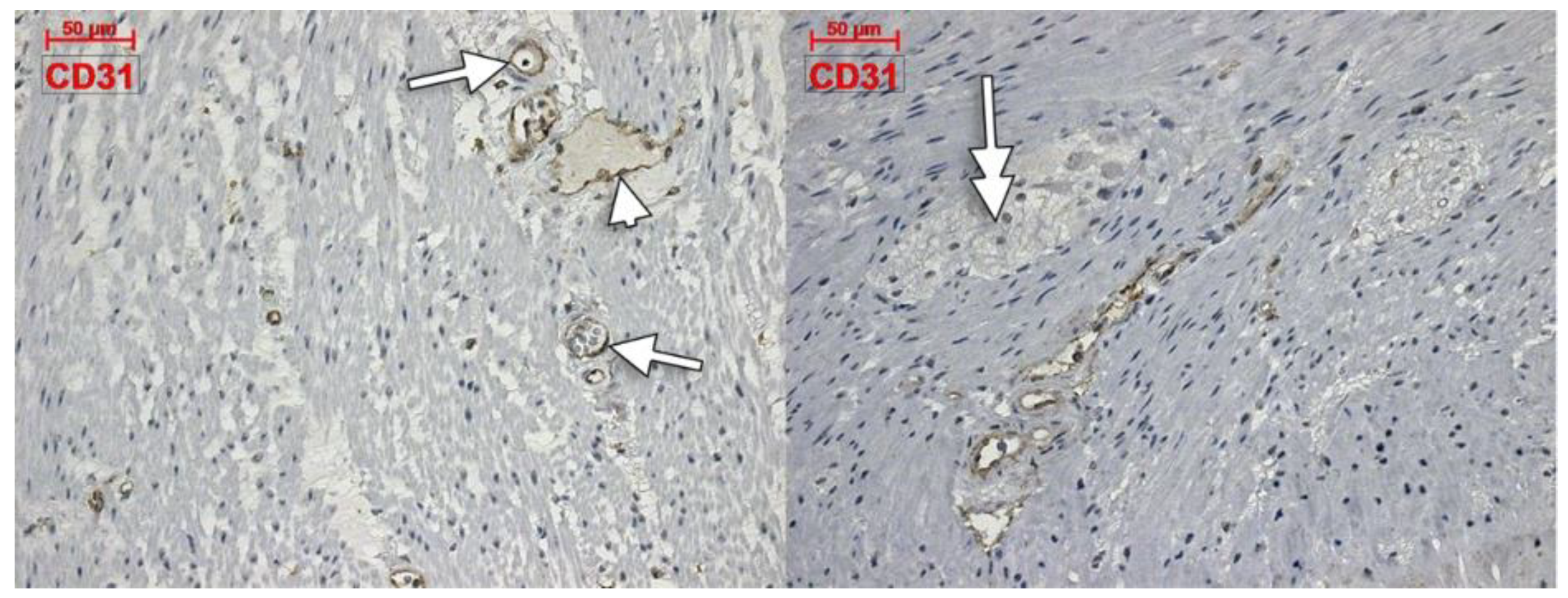
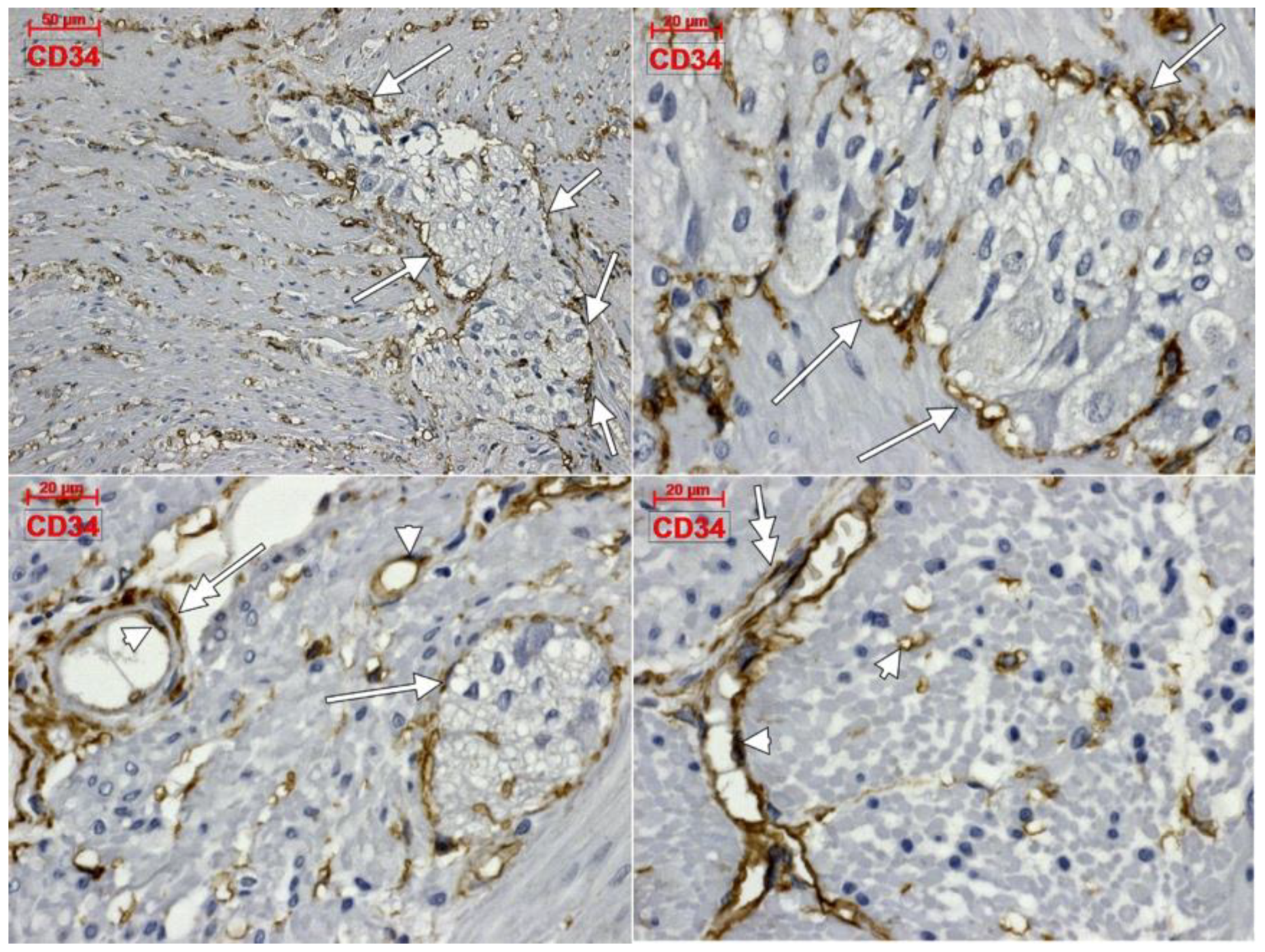
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pieri, L.; Vannucchi, M.G.; Faussone-Pellegrini, M.S. Histochemical and ultrastructural characteristics of an interstitial cell type different from ICC and resident in the muscle coat of human gut. J. Cell Mol. Med. 2008, 12, 1944–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, I.; Marini, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Telocytes and lymphatic endothelial cells: Two immunophenotypically distinct and spatially close cell entities. Acta Histochem. 2020, 122, 151530. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Faussone-Pellegrini, M.S. Telocytes—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobra, M.A.; Vrapciu, A.D.; Pop, F.; Petre, N.; Rusu, M.C. The molecular phenotypes of ureteral telocytes are layer-specific. Acta Histochem. 2018, 120, 41–45. [Google Scholar] [CrossRef]
- Grigoriu, F.; Hostiuc, S.; Vrapciu, A.D.; Rusu, M.C. Subsets of telocytes: The progenitor cells in the human endocardial niche. Rom. J. Morphol. Embryol. 2016, 57 (Suppl. S2), 767–774. [Google Scholar] [PubMed]
- Iancu, C.B.; Rusu, M.C.; Mogoantă, L.; Hostiuc, S.; Toader, O.D. The telocytes in the subepicardial niche. Appl. Sci. 2019, 9, 1615. [Google Scholar] [CrossRef] [Green Version]
- Nicolescu, M.I.; Bucur, A.; Dinca, O.; Rusu, M.C.; Popescu, L.M. Telocytes in parotid glands. Anat. Rec. (Hoboken) 2012, 295, 378–385. [Google Scholar] [CrossRef]
- Petrea, C.E.; Craitoiu, S.; Vrapciu, A.D.; Manoiu, V.S.; Rusu, M.C. The telopode- and filopode-projecting heterogeneous stromal cells of the human sclera niche. Ann. Anat. 2018, 218, 129–140. [Google Scholar] [CrossRef]
- Petrea, C.E.; Rusu, M.C.; Manoiu, V.S.; Vrapciu, A.D. Telocyte-Like Cells Containing Weibel-Palade Bodies in Rat Lamina Fusca. Ann. Anat. 2018, 218, 88–94. [Google Scholar] [CrossRef]
- Rusu, M.C.; Nicolescu, M.I.; Jianu, A.M.; Lighezan, R.; Manoiu, V.S.; Paduraru, D. Esophageal telocytes and hybrid morphologies. Cell Biol. Int. 2012, 36, 1079–1088. [Google Scholar] [CrossRef]
- Rusu, M.C.; Hostiuc, S. Critical review: Cardiac telocytes vs. cardiac lymphatic endothelial cells. Ann. Anat 2018, 222, 40–54. [Google Scholar] [CrossRef]
- Rusu, M.C.; Hostiuc, S.; Fildan, A.P.; Tofolean, D.E. Critical Review: What Cell Types Are the Lung Telocytes? Anat. Rec. (Hoboken) 2019, 303, 1280–1292. [Google Scholar] [CrossRef]
- Toader, O.D.; Rusu, M.C.; Mogoanta, L.; Hostiuc, S.; Jianu, A.M.; Ilie, A.C. An Immunohistochemical Study of Gastric Mucosa and Critical Review Indicate that the Subepithelial Telocytes are Prelymphatic Endothelial Cells. Medicina (Kaunas) 2019, 55, 316. [Google Scholar] [CrossRef] [Green Version]
- Manta, L.; Rusu, M.C.; Pop, F. What podoplanin tells us about cells with telopodes. Ann. Anat. 2018, 218, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, M.I.; Rusu, M.C.; Voinea, L.M.; Vrapciu, A.D.; Bara, R.I. Lymphatic lacunae of the human eye conjunctiva embedded within a stroma containing CD34+ telocytes. J. Cell Mol. Med. 2020, 24, 8871–8875. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.C.; Hostiuc, S.; Vrapciu, A.D.; Mogoanta, L.; Manoiu, V.S.; Grigoriu, F. Subsets of telocytes: Myocardial telocytes. Ann. Anat. 2017, 209, 37–44. [Google Scholar] [CrossRef]
- Alexander, J.S.; Ganta, V.C.; Jordan, P.A.; Witte, M.H. Gastrointestinal lymphatics in health and disease. Pathophysiology 2010, 17, 315–335. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.S.; Chaitanya, G.V.; Grisham, M.B.; Boktor, M. Emerging roles of lymphatics in inflammatory bowel disease. Ann. N. Y. Acad. Sci. 2010, 1207 (Suppl. S1), E75–E85. [Google Scholar] [CrossRef]
- Rahier, J.F.; De Beauce, S.; Dubuquoy, L.; Erdual, E.; Colombel, J.F.; Jouret-Mourin, A.; Geboes, K.; Desreumaux, P. Increased lymphatic vessel density and lymphangiogenesis in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2011, 34, 533–543. [Google Scholar] [CrossRef]
- Sui, G.; Wang, Y.; Yang, C. The intraorgan lymphatics of the colon. Chin. J. Anat. 1991, 4, 1. [Google Scholar]
- Weiland, S.A.; Chen, Y.K. Colonic Lymphatics. In Colonic Diseases; Kioch, T.R., Ed.; Humana Press Inc.: Totiwa, NJ, USA, 2003; pp. 123–140. [Google Scholar]
- Hirai, T.; Nimura, Y.; Sakai, H. The three-dimensional microstructure of intramural lymphatics in the canine large intestine. Gastroenterol. Jpn. 1990, 25, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Kachlik, D.; Ziskova, M.; Miko, M. Lymphatic lacunae of the mucosal folds of human uterine tubes—A rediscovery of forgotten structures and their possible role in reproduction. Ann. Anat. 2018, 219, 121–128. [Google Scholar] [CrossRef]
- Thomsen, L.; Robinson, T.L.; Lee, J.C.; Farraway, L.A.; Hughes, M.J.; Andrews, D.W.; Huizinga, J.D. Interstitial cells of Cajal generate a rhythmic pacemaker current. Nat. Med. 1998, 4, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Manea, C.M.; Rusu, M.C.; Constantin, D.; Manoiu, V.M.; Moldovan, L.; Jianu, A.M. Ultrastructural features of human adipose-derived multipotent mesenchymal stromal cells. Rom. J. Morphol. Embryol. 2014, 55, 1363–1369. [Google Scholar] [PubMed]
- Robinson, T.L.; Sircar, K.; Hewlett, B.R.; Chorneyko, K.; Riddell, R.H.; Huizinga, J.D. Gastrointestinal stromal tumors may originate from a subset of CD34-positive interstitial cells of Cajal. Am. J. Pathol. 2000, 156, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Kindblom, L.G.; Remotti, H.E.; Aldenborg, F.; Meis-Kindblom, J.M. Gastrointestinal pacemaker cell tumor (GIPACT): Gastrointestinal stromal tumors show phenotypic characteristics of the interstitial cells of Cajal. Am. J. Pathol. 1998, 152, 1259–1269. [Google Scholar]
- Hirota, S.; Isozaki, K.; Moriyama, Y.; Hashimoto, K.; Nishida, T.; Ishiguro, S.; Kawano, K.; Hanada, M.; Kurata, A.; Takeda, M.; et al. Gain-of-function mutations of c-kit in human gastrointestinal stromal tumors. Science 1998, 279, 577–580. [Google Scholar] [CrossRef]
- Vanderwinden, J.M.; Rumessen, J.J.; De Laet, M.H.; Vanderhaeghen, J.J.; Schiffmann, S.N. CD34 immunoreactivity and interstitial cells of Cajal in the human and mouse gastrointestinal tract. Cell Tissue Res. 2000, 302, 145–153. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.S. Telocytes express PDGFRalpha in the human gastrointestinal tract. J. Cell Mol. Med. 2013, 17, 1099–1108. [Google Scholar] [CrossRef]
- Manetti, M.; Rosa, I.; Messerini, L.; Ibba-Manneschi, L. Telocytes are reduced during fibrotic remodelling of the colonic wall in ulcerative colitis. J. Cell Mol. Med. 2015, 19, 62–73. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zurzu, M.; Nicolescu, M.I.; Mogoantă, L.; Pantea, S.; Rusu, M.C. Telocytes and Lymphatics of the Human Colon. Life 2021, 11, 1001. https://doi.org/10.3390/life11101001
Zurzu M, Nicolescu MI, Mogoantă L, Pantea S, Rusu MC. Telocytes and Lymphatics of the Human Colon. Life. 2021; 11(10):1001. https://doi.org/10.3390/life11101001
Chicago/Turabian StyleZurzu, Mihai, Mihnea Ioan Nicolescu, Laurențiu Mogoantă, Stelian Pantea, and Mugurel Constantin Rusu. 2021. "Telocytes and Lymphatics of the Human Colon" Life 11, no. 10: 1001. https://doi.org/10.3390/life11101001
APA StyleZurzu, M., Nicolescu, M. I., Mogoantă, L., Pantea, S., & Rusu, M. C. (2021). Telocytes and Lymphatics of the Human Colon. Life, 11(10), 1001. https://doi.org/10.3390/life11101001