The Molecular Effects of a High Fat Diet on Endometrial Tumour Biology

 , , ,
, , , 

Abstract
1. Introduction
2. Results
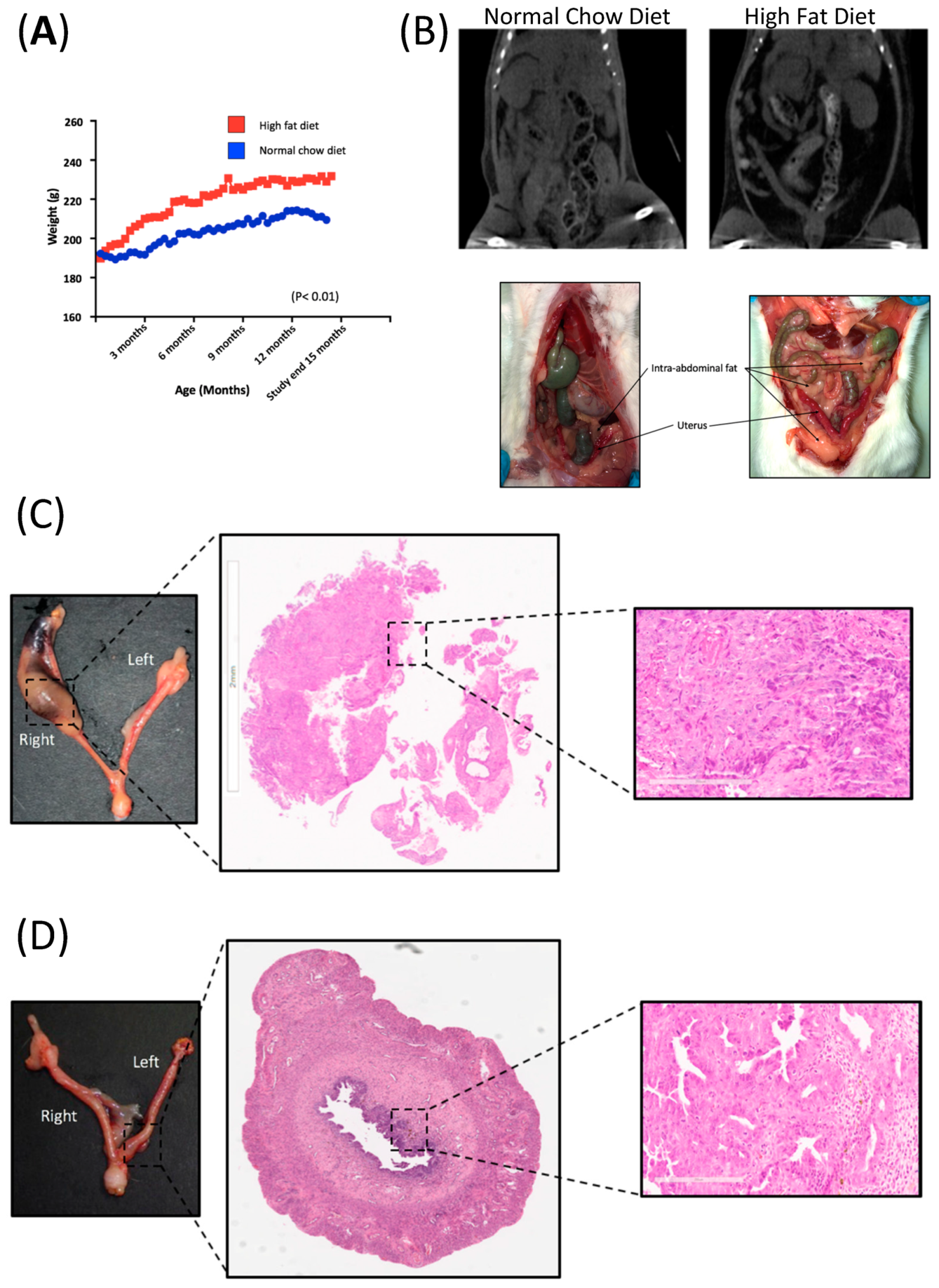
2.1. High Fat Diet-Induced Visceral Obesity in BD/II Han Rats
2.2. Diet-Induced Obesity Did Not Increase the Endometrial Cancer Burden in BDII/Han Rats
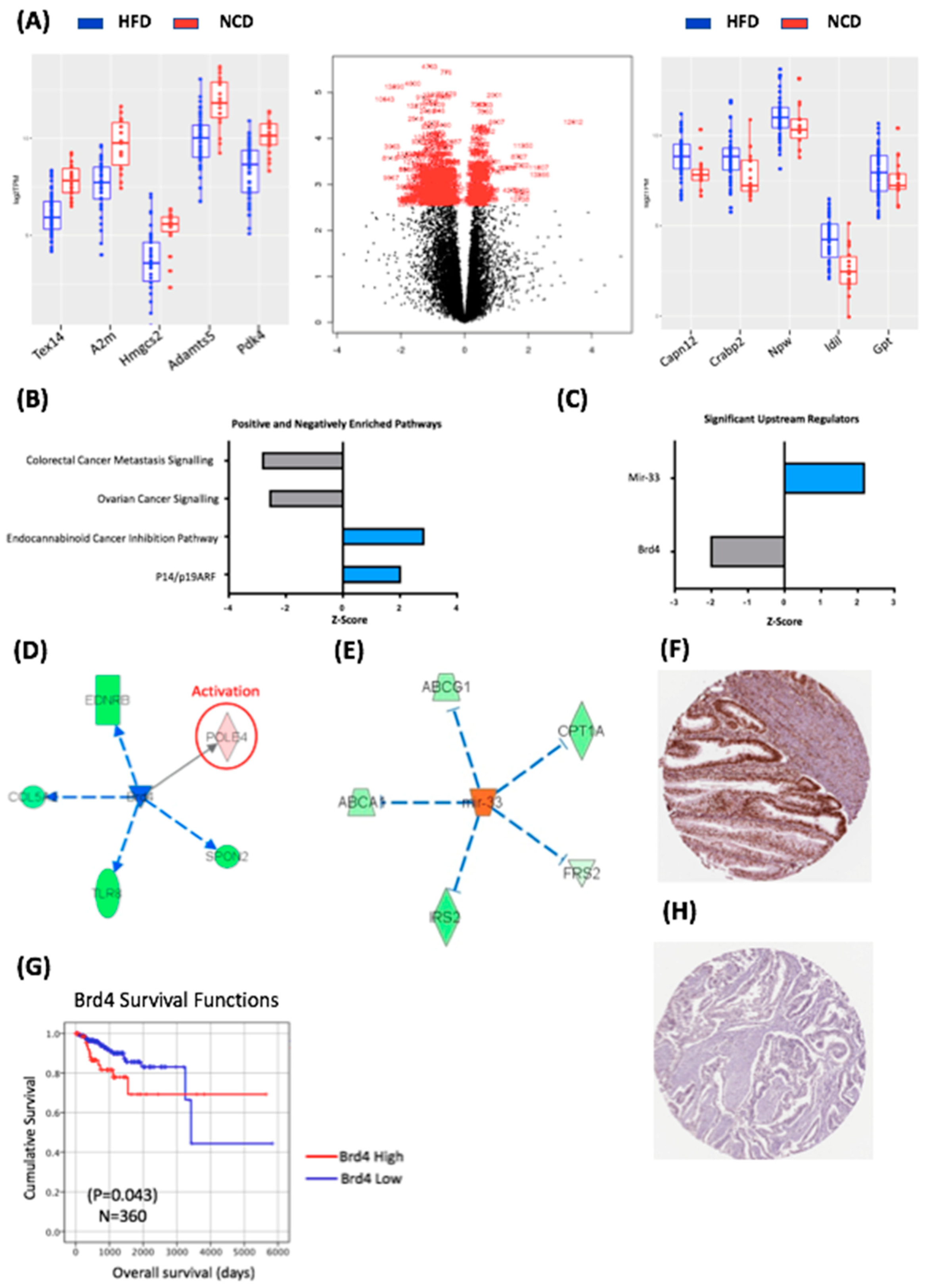
2.3. Diet-Induced Obesity Caused Specific Transcriptomic Changes in BDII/Han Endometrial Tumours
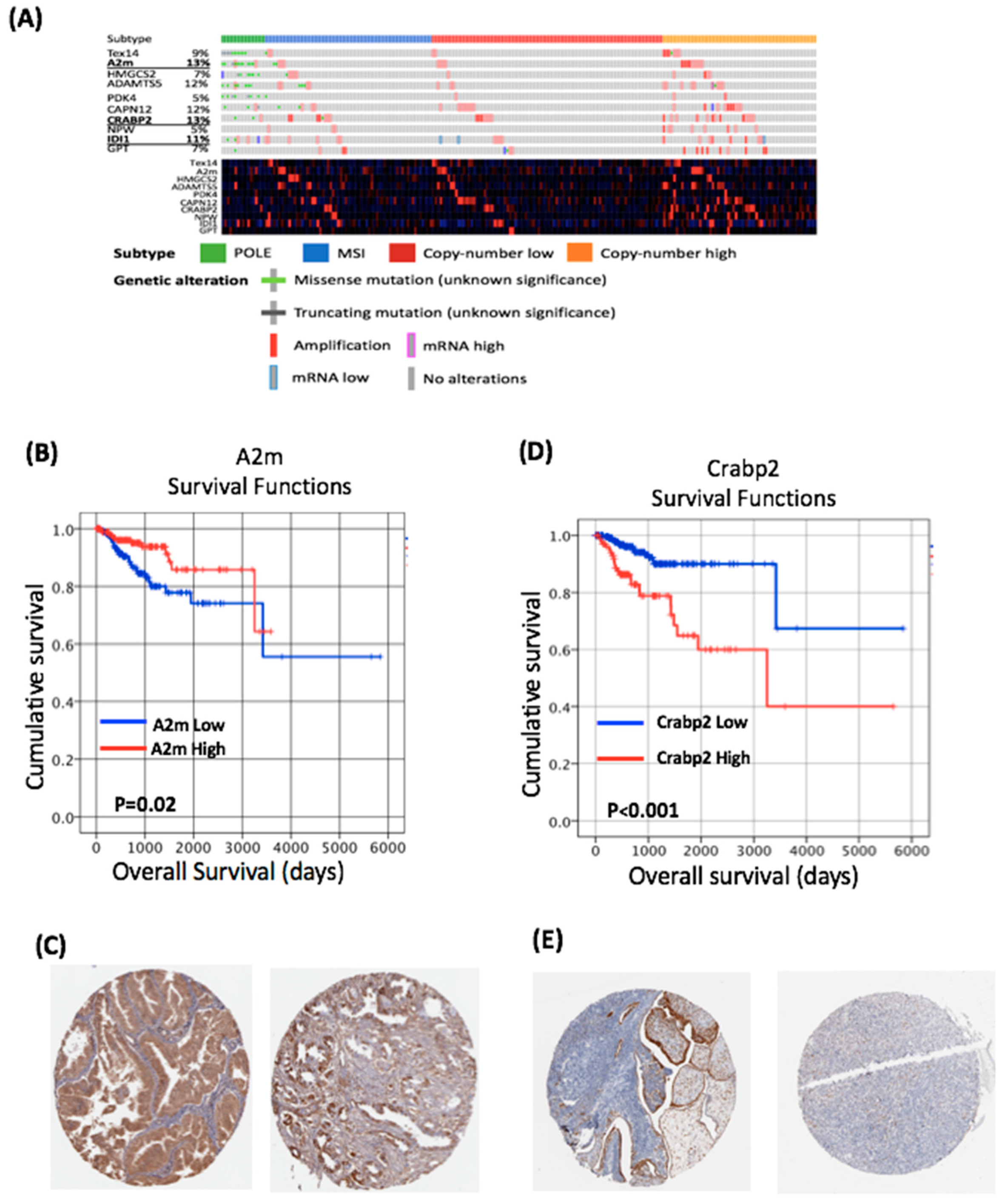
2.4. Obesity-Regulated Genes Can Be Used as Biomarkers in Human Endometrial Cancer
2.5. Mutation and Expression of Tp53, Molecular Characterisation
3. Discussion
4. Materials and Methods
4.1. CT Imaging
4.2. Assessment of Endometrial Cancer Burden
4.3. Retrieval of Endometrial Tissue for Histology and RNA Isolation
4.4. RNA Extraction, Library Prep and Sequencing
4.5. RNAseq Processing, Differential Expression Analysis and Variant Calling
4.6. Pathway and Functional Enrichment Analysis
4.7. TCGA Comparative Model
4.8. Ethics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A
RNA Extraction, Library Prep and Sequencing
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Colombo, N.; Creutzberg, C.; Amant, F.; Bosse, T.; Gonzalez-Martin, A.; Ledermann, J.; Marth, C.; Nout, R.; Querleu, D.; Mirza, M.R.; et al. ESMO-ESGO-ESTRO consensus conference on endometrial cancer: Diagnosis, treatment and follow-up. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2015, 117, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, V.W.; Yang, H.P.; Pike, M.C.; McCann, S.E.; Yu, H.; Xiang, Y.B.; Wolk, A.; Wentzensen, N.; Weiss, N.S.; Webb, P.M.; et al. Type I and II endometrial cancers: Have they different risk factors? J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2013, 31, 2607–2618. [Google Scholar] [CrossRef] [PubMed]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body Fatness and Cancer--Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef]
- Levine, R.L.; Cargile, C.B.; Blazes, M.S.; van Rees, B.; Kurman, R.J.; Ellenson, L.H. PTEN mutations and microsatellite instability in complex atypical hyperplasia, a precursor lesion to uterine endometrioid carcinoma. Cancer Res. 1998, 58, 3254–3258. [Google Scholar]
- Maxwell, G.L.; Risinger, J.I.; Gumbs, C.; Shaw, H.; Bentley, R.C.; Barrett, J.C.; Berchuck, A.; Futreal, P.A. Mutation of the PTEN tumor suppressor gene in endometrial hyperplasias. Cancer Res. 1998, 58, 2500–2503. [Google Scholar]
- Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; Benz, C.C.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef]
- Liu, Y.; Patel, L.; Mills, G.B.; Lu, K.H.; Sood, A.K.; Ding, L.; Kucherlapati, R.; Mardis, E.R.; Levine, D.A.; Shmulevich, I.; et al. Clinical significance of CTNNB1 mutation and Wnt pathway activation in endometrioid endometrial carcinoma. J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef]
- Deerberg, F.; Kaspareit, J. Endometrial carcinoma in BD II/Han rats: Model of a spontaneous hormone-dependent tumor. J. Natl. Cancer Inst. 1987, 78, 1245–1251. [Google Scholar]
- Vollmer, G. Endometrial cancer: Experimental models useful for studies on molecular aspects of endometrial cancer and carcinogenesis. Endocr. Relat. Cancer 2003, 10, 23–42. [Google Scholar] [CrossRef]
- Hogarty, M.D.; Norris, M.D.; Davis, K.; Liu, X.; Evageliou, N.F.; Hayes, C.S.; Pawel, B.; Guo, R.; Zhao, H.; Sekyere, E.; et al. ODC1 is a critical determinant of MYCN oncogenesis and a therapeutic target in neuroblastoma. Cancer Res. 2008, 68, 9735–9745. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, A.; Helou, K.; Walentinsson, A.; Hedrich, H.J.; Szpirer, C.; Levan, G. Amplification of Mycn, Ddx1, Rrm2, and Odc1 in rat uterine endometrial carcinomas. Genes Chromosomes Cancer 2001, 31, 345–356. [Google Scholar] [CrossRef]
- Deerberg, F.; Pohlmeyer, G.; Lorcher, K.; Petrow, V. Total suppression of spontaneous endometrial carcinoma in BDII/Han rats by melengestrol acetate. Oncology 1995, 52, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Adamovic, T.; Trosso, F.; Roshani, L.; Andersson, L.; Petersen, G.; Rajaei, S.; Helou, K.; Levan, G. Oncogene amplification in the proximal part of chromosome 6 in rat endometrial adenocarcinoma as revealed by combined BAC/PAC FISH, chromosome painting, zoo-FISH, and allelotyping. Genes Chromosomes Cancer 2005, 44, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Helou, K.; Walentinsson, A.; Beckmann, B.; Johansson, A.; Hedrich, H.J.; Szpirer, C.; Klinga-Levan, K.; Levan, G. Analysis of genetic changes in rat endometrial carcinomas by means of comparative genomic hybridization. Cancer Genet. Cytogenet. 2001, 127, 118–127. [Google Scholar] [CrossRef]
- Samuelson, E.; Hedberg, C.; Nilsson, S.; Behboudi, A. Molecular classification of spontaneous endometrial adenocarcinomas in BDII rats. Endocr. Relat. Cancer 2009, 16, 99–111. [Google Scholar] [CrossRef]
- Watanabe, J.; Nishizaki, R.; Jobo, T.; Kamata, Y.; Hata, H.; Nishimura, Y.; Fujisawa, T.; Okayasu, I.; Kuramoto, H. Expression of tumor suppressor gene product p14ARF in endometrioid adenocarcinoma of the uterine corpus. Int. J. Gynecol. Pathol. 2004, 23, 234–240. [Google Scholar] [CrossRef]
- Price, N.L.; Singh, A.K.; Rotllan, N.; Goedeke, L.; Wing, A.; Canfrán-Duque, A.; Diaz-Ruiz, A.; Araldi, E.; Baldán, Á.; Camporez, J.P.; et al. Genetic Ablation of miR-33 Increases Food Intake, Enhances Adipose Tissue Expansion, and Promotes Obesity and Insulin Resistance. Cell Rep. 2018, 22, 2133–2145. [Google Scholar] [CrossRef]
- Nagarajan, S.; Hossan, T.; Alawi, M.; Najafova, Z.; Indenbirken, D.; Bedi, U.; Taipaleenmaki, H.; Ben-Batalla, I.; Scheller, M.; Loges, S.; et al. Bromodomain protein BRD4 is required for estrogen receptor-dependent enhancer activation and gene transcription. Cell Rep. 2014, 8, 460–469. [Google Scholar] [CrossRef]
- Jones, D.H.; Lin, D.I. Amplification of the NSD3-BRD4-CHD8 pathway in pelvic high-grade serous carcinomas of tubo-ovarian and endometrial origin. Mol. Clin. Oncol. 2017, 7, 301–307. [Google Scholar] [CrossRef]
- Talhouk, A.; McConechy, M.K.; Leung, S.; Yang, W.; Lum, A.; Senz, J.; Boyd, N.; Pike, J.; Anglesio, M.; Kwon, J.S.; et al. Confirmation of ProMisE: A simple, genomics-based clinical classifier for endometrial cancer. Cancer 2017, 123, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Britton, H.; Huang, L.; Lum, A.; Leung, S.; Shum, K.; Kale, M.; Burleigh, A.; Senz, J.; Yang, W.; McConechy, M.; et al. Molecular classification defines outcomes and opportunities in young women with endometrial carcinoma. Gynecol. Oncol. 2019, 153, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Oncolnc. Available online: http://www.oncolnc.org (accessed on 1 July 2020).
- Atlas, T.H.P. The Human Protein Atlas. Available online: https://www.proteinatlas.org (accessed on 1 July 2020).
- cBioportal. Available online: https://www.cbioportal.org (accessed on 1 July 2020).
- Pimentel, H.; Bray, N.L.; Puente, S.; Melsted, P.; Pachter, L. Differential analysis of RNA-seq incorporating quantification uncertainty. Nat. Methods 2017, 14, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Tayyari, F.; Pinnaduwage, D.; Bayani, J.; Bartlett, J.M.S.; Mulligan, A.M.; Bull, S.B.; Andrulis, I.L. Tumoral BRD4 expression in lymph node-negative breast cancer: Association with T-bet+ tumor-infiltrating lymphocytes and disease-free survival. BMC Cancer 2018, 18, 750. [Google Scholar] [CrossRef]
- Kurz, S.; Thieme, R.; Amberg, R.; Groth, M.; Jahnke, H.G.; Pieroh, P.; Horn, L.C.; Kolb, M.; Huse, K.; Platzer, M.; et al. The anti-tumorigenic activity of A2M-A lesson from the naked mole-rat. PLoS ONE 2017, 12, e0189514. [Google Scholar] [CrossRef] [PubMed]
- Kostova, M.B.; Brennen, W.N.; Lopez, D.; Anthony, L.; Wang, H.; Platz, E.; Denmeade, S.R. PSA-alpha-2-macroglobulin complex is enzymatically active in the serum of patients with advanced prostate cancer and can degrade circulating peptide hormones. Prostate 2018, 78, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Birkenmeier, G.; Muller, R.; Huse, K.; Forberg, J.; Glaser, C.; Hedrich, H.; Nicklisch, S.; Reichenbach, A. Human alpha2-macroglobulin: Genotype-phenotype relation. Exp. Neurol. 2003, 184, 153–161. [Google Scholar] [CrossRef]
- Ochiai, A.; Kuroda, K.; Ozaki, R.; Ikemoto, Y.; Murakami, K.; Muter, J.; Matsumoto, A.; Itakura, A.; Brosens, J.J.; Takeda, S. Resveratrol inhibits decidualization by accelerating downregulation of the CRABP2-RAR pathway in differentiating human endometrial stromal cells. Cell Death Dis. 2019, 10, 276. [Google Scholar] [CrossRef]
- Delva, L.; Bastie, J.N.; Rochette-Egly, C.; Kraïba, R.; Balitrand, N.; Despouy, G.; Chambon, P.; Chomienne, C. Physical and functional interactions between cellular retinoic acid binding protein II and the retinoic acid-dependent nuclear complex. Mol. Cell. Biol. 1999, 19, 7158–7167. [Google Scholar] [CrossRef]
- Vreeland, A.C.; Yu, S.; Levi, L.; de Barros Rossetto, D.; Noy, N. Transcript stabilization by the RNA-binding protein HuR is regulated by cellular retinoic acid-binding protein 2. Mol. Cell. Biol. 2014, 34, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-I.; Lin, Y.-P.; Tseng, C.-W.; Chen, H.-J.; Wang, L.-H. Crabp2 Promotes Metastasis of Lung Cancer Cells via HuR and Integrin β1/FAK/ERK Signaling. Sci. Rep. 2019, 9, 845. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Parameswaran, N.; Li, M.; Wang, Y.; Jackson, M.W.; Liu, H.; Xin, W.; Zhou, L. CRABP-II enhances pancreatic cancer cell migration and invasion by stabilizing interleukin 8 expression. Oncotarget 2017, 8, 52432–52444. [Google Scholar] [CrossRef] [PubMed]
- Toyama, A.; Suzuki, A.; Shimada, T.; Aoki, C.; Aoki, Y.; Umino, Y.; Nakamura, Y.; Aoki, D.; Sato, T.A. Proteomic characterization of ovarian cancers identifying annexin-A4, phosphoserine aminotransferase, cellular retinoic acid-binding protein 2, and serpin B5 as histology-specific biomarkers. Cancer Sci. 2012, 103, 747–755. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, M.; Wang, B.; Zhou, C.; Mu, Y.; Li, J.; Liu, X.; Wang, Y.; Song, Z.; Liu, P. CRABP2 regulates invasion and metastasis of breast cancer through hippo pathway dependent on ER status. J. Exp. Clin. Cancer Res. 2019, 38, 361. [Google Scholar] [CrossRef]
- Baxter, E.; Windloch, K.; Kelly, G.; Lee, J.S.; Gannon, F.; Brennan, D.J. Molecular basis of distinct oestrogen responses in endometrial and breast cancer. Endocr. Relat. Cancer 2019, 26, 31–46. [Google Scholar] [CrossRef]
- Ruehl-Fehlert, C.; Kittel, B.; Morawietz, G.; Deslex, P.; Keenan, C.; Mahrt, C.R.; Nolte, T.; Robinson, M.; Stuart, B.P.; Deschl, U. Revised guides for organ sampling and trimming in rats and mice—Part 1. Exp. Toxicol. Pathol. 2003, 55, 91–106. [Google Scholar] [CrossRef]
- Morawietz, G.; Ruehl-Fehlert, C.; Kittel, B.; Bube, A.; Keane, K.; Halm, S.; Heuser, A.; Hellmann, J. Revised guides for organ sampling and trimming in rats and mice—Part 3. A joint publication of the RITA and NACAD groups. Exp. Toxicol. Pathol. 2004, 55, 433–449. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Li, L.; Feng, Q.; Wang, X. PreMSIm: An R package for predicting microsatellite instability from the expression profiling of a gene panel in cancer. Comput. Struct. Biotechnol. J. 2020, 18, 668–675. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| A2M Low (n = 191) | A2M High (n = 180) | p-Value | Crabp2 Low (n = 265) | Crabp2 High (n = 106) | p-Value | |
|---|---|---|---|---|---|---|
| Age Median (range) | 63 (35–90) | 63 (31–90) | 0.665 | 63 (33–90) | 65 (31–90) | 0.093 * |
| BMI Median (range) | 33.1 (19–82) | 32.3 (17–61) | <0.001 * | 33 (17–82) | 31 (19–68) | 0.485 * |
| Histology | ||||||
| Endometrioid | 147 (77) | 158 (87) | 0.003 | 238(90) | 67 (63) | <0.001 |
| Serous | 32 (16.7) | 21 (12) | 20 (7) | 33 (31) | ||
| Mixed | 12 (6.3) | 1 (1) | 7 (3) | 6 (6) | ||
| Stage | ||||||
| 1 | 120 (63.1) | 132(74.2) | 0.15 | 194 (74) | 59 (55) | 0.005 |
| 2 | 16 (8.4) | 10 (5) | 16 (6) | 10 (9) | ||
| 3 | 44 (23.1) | 30 (16.8) | 47 (18) | 28 (26) | ||
| 4 | 11 (5.4) | 8 (4) | 8 (2) | 9 (9) | ||
| Grade | ||||||
| 1 | 31 (16.2) | 58 (32.2) | <0.001 | 74 (28) | 15 (14) | 0.001 |
| 2 | 50 (26.1) | 55 (30.5) | 77 (29) | 28 (26) | ||
| 3 | 110 (57.7) | 67 (37.3) | 114 (43) | 63 (59) | ||
| TCGA molecular subtype | ||||||
| POLE | 11 (10.2) | 6 (4.8) | 0.28 | 16 (10) | 1 (2) | <0.001 |
| MSI | 30 (28) | 35 (28) | 58 (35) | 7 (10) | ||
| CN Low | 32 (30) | 58 (46.4) | 65 (39) | 25 (37) | ||
| CN High | 34 (31.8) | 26 (20.8) | 26 (16) | 34 (51) |
| Univariate | Multivariate | |||||||
|---|---|---|---|---|---|---|---|---|
| HR | 95.0% CI | p-Value | HR | 95.0% CI | p-Value | |||
| Overall Survival | Lower | Upper | Lower | Upper | ||||
| A2M (low vs. high) | 0.455 | 0.23 | 0.9 | 0.024 | 0.612 | 0.305 | 1.229 | 0.167 |
| Age (continuous) | 1.041 | 1.009 | 1.075 | 0.013 | 1.053 | 1.015 | 1.092 | 0.005 |
| Histology (endometrioid vs. non-endometrioid | 2.298 | 1.18 | 4.475 | 0.014 | 0.705 | 0.311 | 1.595 | 0.401 |
| Stage (1 and 2 vs. 3 and 4) | 4.654 | 2.448 | 8.849 | <0.001 | 3.912 | 1.903 | 8.042 | <0.001 |
| Grade (1 and 2 vs. 3) | 3.393 | 1.652 | 6.969 | 0.001 | 1.848 | 0.796 | 4.289 | 0.153 |
| HR | 95.0% CI | p-Value | HR | 95.0% CI | p-Value | |||
| Overall Survival | Lower | Upper | Lower | Upper | ||||
| Crabp2 (low vs. high) | 3.554 | 1.875 | 6.735 | <0.001 | 3.134 | 1.524 | 6.443 | 0.002 |
| Age (continuous) | 1.041 | 1.009 | 1.075 | 0.013 | 1.046 | 1.011 | 1.083 | 0.01 |
| Histology (endometrioid vs. non-endometrioid) | 2.298 | 1.18 | 4.475 | 0.014 | 0.451 | 0.193 | 1.052 | 0.065 |
| Stage (1 and 2 vs. 3 and 4) | 4.654 | 2.448 | 8.849 | <0.001 | 3.875 | 1.868 | 8.036 | <0.001 |
| Grade (1 and 2 vs. 3) | 3.393 | 1.652 | 6.969 | 0.001 | 1.802 | 0.775 | 4.19 | 0.171 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilkinson, M.; Sinclair, P.; Dellatorre-Teixeira, L.; Swan, P.; Brennan, E.; Moran, B.; Wedekind, D.; Downey, P.; Sheahan, K.; Conroy, E.; et al. The Molecular Effects of a High Fat Diet on Endometrial Tumour Biology. Life 2020, 10, 188. https://doi.org/10.3390/life10090188
Wilkinson M, Sinclair P, Dellatorre-Teixeira L, Swan P, Brennan E, Moran B, Wedekind D, Downey P, Sheahan K, Conroy E, et al. The Molecular Effects of a High Fat Diet on Endometrial Tumour Biology. Life. 2020; 10(9):188. https://doi.org/10.3390/life10090188
Chicago/Turabian StyleWilkinson, Michael, Piriyah Sinclair, Ludmilla Dellatorre-Teixeira, Patrick Swan, Eoin Brennan, Bruce Moran, Dirk Wedekind, Paul Downey, Kieran Sheahan, Emer Conroy, and et al. 2020. "The Molecular Effects of a High Fat Diet on Endometrial Tumour Biology" Life 10, no. 9: 188. https://doi.org/10.3390/life10090188
APA StyleWilkinson, M., Sinclair, P., Dellatorre-Teixeira, L., Swan, P., Brennan, E., Moran, B., Wedekind, D., Downey, P., Sheahan, K., Conroy, E., Gallagher, W. M., Docherty, N., Roux, C. l., & Brennan, D. J. (2020). The Molecular Effects of a High Fat Diet on Endometrial Tumour Biology. Life, 10(9), 188. https://doi.org/10.3390/life10090188