The Role of Long Non-Coding RNAs in Intracranial Aneurysms and Subarachnoid Hemorrhage

 ,
,  ,
,
Abstract
1. Introduction
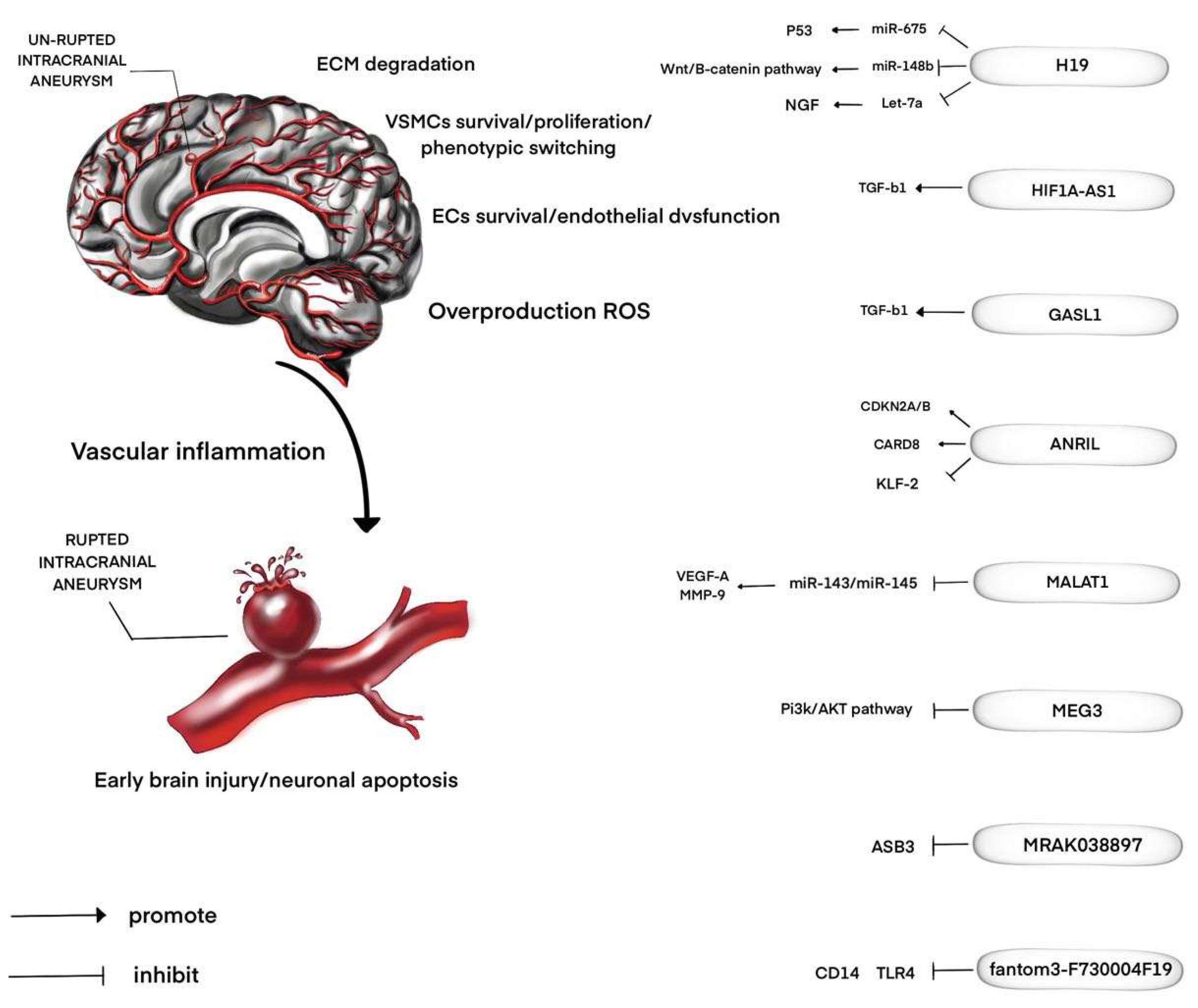
2. LncRNAs and IAs
3. LncRNAs and SAH
4. LncRNAs as Non-Invasive Biomarkers
5. LncRNA-Based Therapeutics
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jin, D.; Song, C.; Leng, X.; Han, P. A systematic review and meta-analysis of risk factors for unruptured intracranial aneurysm growth. Int J. Surg. 2019, 69, 68–76. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S. Aneurysmal Subarachnoid Hemorrhage. J. Neurosurg. Anesth. 2015, 27, 222–240. [Google Scholar] [CrossRef] [PubMed]
- Lepoivre, C.; Belhocine, M.; Bergon, A.; Griffon, A.; Yammine, M.; Vanhille, L.; Zacarias-Cabeza, J.; Garibal, M.A.; Koch, F.; Maqbool, M.A.; et al. Divergent transcription is associated with promoters of transcriptional regulators. BMC Genom. 2013, 14, 914. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef]
- Zhang, X.; Hong, R.; Chen, W.; Xu, M.; Wang, L. The role of long noncoding RNA in major human disease. Bioorg. Chem. 2019, 92, 103214. [Google Scholar] [CrossRef]
- Huang, B.; Lu, S.; Lai, H.; Li, J.; Sun, Y.; Wang, C. LncRNA LOXL1-AS is up-regulated in thoracic aortic aneurysm and regulated proliferation and apoptosis of aortic smooth muscle cells. Biosci. Rep. 2019, 39, BSR20191649. [Google Scholar] [CrossRef]
- Guo, X.; Chang, Q.; Pei, H.; Sun, X.; Qian, X.; Tian, C.; Lin, H. Long Non-coding RNA-mRNA Correlation Analysis Reveals the Potential Role of HOTAIR in Pathogenesis of Sporadic Thoracic Aortic Aneurysm. Eur. J. Vasc. Endovasc. Surg. 2017, 54, 303–314. [Google Scholar] [CrossRef]
- Yu, B.; Liu, L.; Sun, H.; Chen, Y. Long noncoding RNA AK056155 involved in the development of Loeys-Dietz syndrome through AKT/PI3K signaling pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 10768. [Google Scholar]
- Li, Y.; Liu, Y.; Liu, S.; Wu, F.; Li, S.; Yang, F.; Gu, Y.; Xu, Z.; Wang, G. Differential expression profile of long non-coding RNAs in human thoracic aortic aneurysm. J. Cell Biochem. 2018, 119, 7991–7997. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, H.; Yu, C.; Lu, R.; Song, T.; Wang, X.; Tang, W.; Gao, Y. Long noncoding RNA myocardial infarction associated transcript promotes the development of thoracic aortic by targeting microRNA-145 via the PI3K/Akt signaling pathway. J. Cell Biochem. 2019, 120, 14405–14413. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, W.; Yao, K.; Wang, Z.; He, H. Interleukin 6 induces expression of NADPH oxidase 2 in human aortic endothelial cells via long noncoding RNA MALAT1. Pharmazie 2016, 71, 592–597. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Tan, J.; Yu, B.; Shi, W.; Liang, K. Long noncoding RNA HIF1A-AS1A reduces apoptosis of vascular smooth muscle cells: Implications for the pathogenesis of thoracoabdominal aorta aneurysm. Pharmazie 2015, 70, 310–315. [Google Scholar] [PubMed]
- Zhao, Y.; Feng, G.; Wang, Y.; Yue, Y.; Zhao, W. Regulation of apoptosis by long non-coding RNA HIF1A-AS1 in VSMCs: Implications for TAA pathogenesis. Int. J. Clin. Exp. Pathol. 2014, 7, 7643–7652. [Google Scholar] [PubMed]
- Lino Cardenas, C.L.; Kessinger, C.W.; Cheng, Y.; MacDonald, C.; MacGillivray, T.; Ghoshhajra, B.; Huleihel, L.; Nuri, S.; Yeri, A.S.; Jaffer, F.A.; et al. An HDAC9-MALAT1-BRG1 complex mediates smooth muscle dysfunction in thoracic aortic aneurysm. Nat. Commun. 2018, 9, 1009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zou, G.; Chen, X.; Lu, W.; Liu, J.; Zhai, S.; Qiao, G. Knockdown of lncRNA PVT1 Inhibits Vascular Smooth Muscle Cell Apoptosis and Extracellular Matrix Disruption in a Murine Abdominal Aortic Aneurysm Model. Mol. Cells 2019, 42, 218–227. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, X.; Yuan, Y.; Tan, M.; Zhang, L.; Xue, X.; Yan, Y.; Han, L.; Xu, Z. BRG1 expression is increased in thoracic aortic aneurysms and regulates proliferation and apoptosis of vascular smooth muscle cells through the long non-coding RNA HIF1A-AS1 in vitro. Eur. J. Cardiothorac. Surg. 2015, 47, 439–446. [Google Scholar] [CrossRef]
- He, X.; Wang, S.; Li, M.; Zhong, L.; Zheng, H.; Sun, Y.; Lai, Y.; Chen, X.; Wei, G.; Si, X.; et al. Long noncoding RNA GAS5 induces abdominal aortic aneurysm formation by promoting smooth muscle apoptosis. Theranostics 2019, 9, 5558–5576. [Google Scholar] [CrossRef]
- Li, D.Y.; Busch, A.; Jin, H.; Chernogubova, E.; Pelisek, J.; Karlsson, J.; Sennblad, B.; Liu, S.; Lao, S.; Hofmann, P.; et al. H19 induces abdominal aortic aneurysm development and progression. Circulation 2018, 138, 1551–1568. [Google Scholar] [CrossRef]
- Sun, Y.; Zhong, L.; He, X.; Wang, S.; Lai, Y.; Wu, W.; Song, H.; Chen, Y.; Yang, Y.; Liao, W.; et al. LncRNA H19 promotes vascular inflammation and abdominal aortic aneurysm formation by functioning as a competing endogenous RNA. J. Mol. Cell Cardiol. 2019, 131, 66–81. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, C.; Qin, Q.; Liu, Z.; Li, P. LncRNA LEF1-AS1 regulates the migration and proliferation of vascular smooth muscle cells by targeting miR-544a/PTEN axis. J. Cell Biochem. 2019, 120, 14670–14678. [Google Scholar] [CrossRef]
- Cui, C.; Wang, X.; Shang, X.M.; Li, L.; Ma, Y.; Zhao, G.Y.; Song, Y.X.; Geng, X.B.; Zhao, B.Q.; Tian, M.R.; et al. lncRNA 430945 promotes the proliferation and migration of vascular smooth muscle cells via the ROR2/RhoA signaling pathway in atherosclerosis. Mol. Med. Rep. 2019, 19, 4663–4672. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ma, S.; Sun, L.; Gao, J.; Zhao, C. TGF-β1 upregulates the expression of lncRNA-ATB to promote atherosclerosis. Mol. Med. Rep. 2019, 19, 4222–4228. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Meng, Q.; Qi, M.; Li, F.; Liu, B. Shear-Sensitive lncRNA AF131217.1 Inhibits Inflammation in HUVECs via Regulation of KLF4. Hypertension 2019, 73, e25–e34. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, X.; Wu, Y.; Yang, Y.; Gao, L.; Qiao, X.; Chen, T. LncRNA AK094457 promotes AngII-mediated hypertension and endothelial dysfunction through suppressing of activation of PPARγ. Life Sci. 2019, 233, 116745. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.Z.; Li, Z.J.; Liu, W.T.; Shan, J.J.; Wang, L.; Su, Q. Down-regulation of lncRNA MALAT1 alleviates vascular lesion and vascular remodeling of rats with hypertension. Aging (Albany NY) 2019, 11, 5192–5205. [Google Scholar] [CrossRef]
- Fang, G.; Qi, J.; Huang, L.; Zhao, X. LncRNA MRAK048635_P1 is critical for vascular smooth muscle cell function and phenotypic switching in essential hypertension. Biosci. Rep. 2019, 39, BSR20182229. [Google Scholar] [CrossRef]
- Wang, W.; Li, H.; Yu, L.; Zhao, Z.; Wang, H.; Zhang, D.; Zhang, Y.; Lan, Q.; Wang, J.; Zhao, J. Aberrant expression of lncRNAs and mRNAs in patients with intracranial aneurysm. Oncotarget 2017, 8, 2477–2484. [Google Scholar] [CrossRef]
- Hung, J.; Miscianinov, V.; Sluimer, J.C.; Newby, D.E.; Baker, A.H. Targeting Non-coding RNA in Vascular Biology and Disease. Front. Physiol. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Chen, Y.; Li, G.; Fan, H.; Guo, S.; Li, R.; Yin, J.; Zhang, X.; Li, X.; He, X.; Duan, C. CDKN2BAS gene polymorphisms and the risk of intracranial aneurysm in the Chinese population. BMC Neurol. 2017, 17, 214. [Google Scholar] [CrossRef]
- Helgadottir, A.; Thorleifsson, G.; Magnusson, K.P.; Grétarsdottir, S.; Steinthorsdottir, V.; Manolescu, A. The same sequence variant on 9p21 associates with myocardial infarction, abdominal aortic aneurysm and intracranial aneurysm. Nat. Genet. 2008, 40, 217–224. [Google Scholar] [CrossRef]
- Low, S.K.; Takahashi, A.; Cha, P.C.; Zembutsu, H.; Kamatani, N.; Kubo, M.; Nakamura, Y. Genome-wide association study for intracranial aneurysm in the Japanese population identifies three candidate susceptible loci and a functional genetic variant at EDNRA. Hum. Mol. Genet. 2012, 21, 2102–2110. [Google Scholar] [CrossRef] [PubMed]
- Alg, V.S.; Sofat, R.; Houlden, H.; Werring, D.J. Genetic risk factors for intracranial aneurysms: A meta-analysis in more than 116,000 individuals. Neurology 2013, 80, 2154–2165. [Google Scholar] [CrossRef] [PubMed]
- Mongelli, A.; Martelli, F.; Farsetti, A.; Gaetano, C. The Dark That Matters: Long Non-coding RNAs as Master Regulators of Cellular Metabolism in Non-communicable Diseases. Front. Physiol. 2019, 10, 369. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiutian, S. Replication of GWAS Loci Revealed an Increased Risk of BET1L and H19 Polymorphisms with Intracranial Aneurysm. Dis. Markers 2019, 2019, 1–8. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Chu, L.; Chen, W.; Du, Y.; Gu, J. Long non-coding RNA HIF1A-AS1 is upregulated in intracranial aneurysms and participates in the regulation of proliferation of vascular smooth muscle cells by upregulating TGF-β1. Exp. Ther. Med. 2019, 17, 1797–1801. [Google Scholar] [CrossRef]
- Man, H.; Bi, W. Expression of a Novel Long Noncoding RNA (lncRNA), GASL1, is Downregulated in Patients with Intracranial Aneurysms and Regulates the Proliferation of Vascular Smooth Muscle Cells In Vitro. Med. Sci. Monit. 2019, 25, 1133–1139. [Google Scholar] [CrossRef]
- Li, H.; Yue, H.; Hao, Y.; Li, H.; Wang, S.; Yu, L.; Zhang, D.; Cao, Y.; Zhao, J. Expression profile of long noncoding RNAs in human cerebral aneurysms: A microarray analysis. J. Neurosurg. 2016, 127, 1055–1062. [Google Scholar] [CrossRef]
- Rass, V.; Helbok, R. Early Brain Injury after Poor-Grade Subarachnoid Hemorrhage. Curr. Neurol. Neurosci. Rep. 2019, 19, 78. [Google Scholar] [CrossRef]
- Li, Y.; Wu, P.; Bihl, J.C.; Shi, H. Underlying Mechanisms and Potential Therapeutic Molecular Targets in Blood-Brain Barrier Disruption After Subarachnoid Hemorrhage. Curr. Neuropharmacol. 2020. [CrossRef]
- Qian, C.; Jin, J.; Chen, J.; Li, J.; Yu, X.; Mo, H.; Chen, G. SIRT1 activation by resveratrol reduces brain edema and neuronal apoptosis in an experimental rat subarachnoid hemorrhage model. Mol. Med. Rep. 2017, 16, 9627–9635. [Google Scholar] [CrossRef]
- Zhang, N.; Jiang, T.; Wang, Y.; Wang, S.; Hu, L.; Bu, Y. BTG4 is A Novel p53 Target Gene That Inhibits Cell Growth and Induces Apoptosis. Genes (Basel) 2020, 11, 217. [Google Scholar] [CrossRef]
- Yang, S.; Tang, W.; He, Y.; Wen, L.; Sun, B.; Li, S. Long non-coding RNA and microRNA-675/let-7a mediates the protective effect of melatonin against early brain injury after subarachnoid hemorrhage via targeting TP53 and neural growth factor. Cell Death Dis. 2018, 9, 99. [Google Scholar] [CrossRef]
- Liang, Z.; Chi, Y.J.; Lin, G.Q.; Xiao, L.F.; Su, G.L.; Yang, L.M. LncRNA MEG3 participates in neuronal cell injury induced by subarachnoid hemorrhage via inhibiting the Pi3k/Akt pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2824–2831. [Google Scholar] [CrossRef]
- Liu, J.; Li, Q.; Zhang, K.S.; Hu, B.; Niu, X.; Zhou, S.M.; Li, S.G.; Luo, Y.P.; Wang, Y.; Deng, Z.F. Downregulation of the Long Non-Coding RNA Meg3 Promotes Angiogenesis After Ischemic Brain Injury by Activating Notch Signaling. Mol. Neurobiol. 2017, 54, 8179–8190. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Liu, H.; Wang, R.; Xu, S.; Liu, Y.; Wang, K.; Hou, X.; Shen, C.; Wu, J.; Chen, X.; et al. Expression signatures of long non-coding RNAs in early brain injury following experimental subarachnoid hemorrhage. Mol. Med. Rep. 2015, 12, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.H.; Feske, S.K.; Atherton, J.; Konigsberg, R.G.; De Jager, P.L.; Du, R.; Ogilvy, C.S.; Lo, E.H.; Ning, M. Early elevation of serum tumor necrosis factor-alpha is associated with poor outcome in subarachnoid hemorrhage. J. Investig. Med. 2012, 60, 1054–1058. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Turon, R.; Mendes, A.; Melo, N.; Lacerda, P.; Brasil, P.; Bozza, F.A.; Kurtz, P.; Righy, C. Effect of Early Brain Infarction After Subarachnoid Hemorrhage: A Systematic Review and Meta-Analysis. World Neurosurg. 2018, 115, e292–e298. [Google Scholar] [CrossRef]
- Peng, J.; Wu, Y.; Tian, X.; Pang, J.; Kuai, L.; Cao, F.; Qin, X.; Zhong, J.; Li, X.; Li, Y.; et al. High-Throughput Sequencing and Co-Expression Network Analysis of lncRNAs and mRNAs in Early Brain Injury Following Experimental Subarachnoid Haemorrhage. Sci. Rep. 2017, 7, 46577. [Google Scholar] [CrossRef]
- Ahmed, S.U.; Mocco, J.; Zhang, X.; Kelly, M.; Doshi, A.; Nael, K.; De Leacy, R. MRA versus DSA for the follow-up imaging of intracranial aneurysms treated using endovascular techniques: A meta-analysis. J. Neurointerv. Surg. 2019, 11, 1009–1014. [Google Scholar] [CrossRef]
- Sohel, M.M.H. Circulating microRNAs as biomarkers in cancer diagnosis. Life Sci. 2020, 248, 117473. [Google Scholar] [CrossRef]
- Li, P.; Duan, S.; Fu, A. Long noncoding RNA NEAT1 correlates with higher disease risk, worse disease condition, decreased miR-124 and miR-125a and predicts poor recurrence-free survival of acute ischemic stroke. J. Clin. Lab. Anal. 2019, e23056. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Guo, J.; Ai, F. Circulating long noncoding RNA ANRIL downregulation correlates with increased risk, higher disease severity and elevated pro-inflammatory cytokines in patients with acute ischemic stroke. J. Clin. Lab. Anal. 2019, 33, e22629. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Li, X.M.; Song, N.; Zhai, H.; Gao, X.M.; Yang, Y.N. Long non-coding RNAs H19, MALAT1 and MIAT as potential novel biomarkers for diagnosis of acute myocardial infarction. Biomed. Pharmacother. 2019, 118, 109208. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Y.F.; Yang, X.C.; Xu, L.; Li, W.M.; Xia, K.; Zhang, D.P.; Wu, R.N.; Gan, T. Circulating Long Noncoding RNA LIPCAR Acts as a Novel Biomarker in Patients with ST-Segment Elevation Myocardial Infarction. Med. Sci. Monit. 2018, 24, 5064–5070. [Google Scholar] [CrossRef]
- Yin, Q.; Wu, A.; Liu, M. Plasma Long Non-Coding RNA (lncRNA) GAS5 is a New Biomarker for Coronary Artery Disease. 2017;23:6042-6048. Med. Sci. Monit. 2017, 23, 6042–6048. [Google Scholar] [CrossRef]
- Kumarswamy, R.; Bauters, C.; Volkmann, I.; Maury, F.; Fetisch, J.; Holzmann, A.; Lemesle, G.; de Groote, P.; Pinet, F.; Thum, T. Circulating long noncoding RNA, LIPCAR, predicts survival in patients with heart failure. Circ. Res. 2014, 114, 1569–1575. [Google Scholar] [CrossRef]
- Zhang, X.; Nie, X.; Yuan, S.; Li, H.; Fan, J.; Li, C.; Sun, Y.; Zhao, Y.; Hou, H.; Wang, D.W.; et al. Circulating Long Non-coding RNA ENST00000507296 Is a Prognostic Indicator in Patients with Dilated Cardiomyopathy. Mol. Ther. Nucleic. Acids. 2019, 16, 82–90. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, J.; Xu, J.; Wang, X.; Zhang, X.; Wang, W.; Chen, L.; Yuan, T. Circulating exosomes and exosomal lncRNA HIF1A-AS1 in atherosclerosis. Int. J. Clin. Exp. Pathol. 2017, 10, 8383–8388. [Google Scholar]
- Ouyang, Y.; Jiang, Y.; Yu, M.; Zhang, M.; Tan, Z. Upregulation of MALAT1 expression predicts a poor prognosis in the development of intracranial aneurysm (IA). Int. J. Clin. Exp. Pathol. 2017, 10, 5907–5912. [Google Scholar]
- Wu, C.; Song, H.; Wang, Y.; Gao, L.; Cai, Y.; Cheng, Q.; Chen, Y.; Zheng, Z.; Liao, Y.; Lin, J.; et al. Long non-coding RNA TCONS_00000200 as a non-invasive biomarker in patients with intracranial aneurysm. Biosci. Rep. 2019, 39, BSR20182224. [Google Scholar] [CrossRef]
- Ma, J.; Han, L.N.; Song, J.R.; Bai, X.M.; Wang, J.Z.; Meng, L.F.; Li, J.; Zhou, W.; Feng, Y.; Feng, W.R.; et al. Long noncoding RNA LINC01234 silencing exerts an anti-oncogenic effect in esophageal cancer cells through microRNA-193a-5p-mediated CCNE1 downregulation. Cell. Oncol. (Dordr) 2020. [Google Scholar] [CrossRef]
- Zuo, X.; Chen, Z.; Gao, W.; Zhang, Y.; Wang, J.; Wang, J.; Cao, M.; Cai, J.; Wu, J.; Wang, X. M6A-mediated upregulation of LINC00958 increases lipogenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. J. Hematol. Oncol. 2020, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pinera, P.; Jones, M.F.; Lal, A.; Lu, T.K. Putting Non-coding RNA on Display with CRISPR. Mol. Cell 2015, 59, 146–148. [Google Scholar] [CrossRef][Green Version]
- Fontanella, M.; Gallone, S.; Panciani, P.P.; Garbossa, D.; Stefini, R.; Latronico, N.; Rubino, E.; Marengo, N.; Ducati, A.; Pinessi, L.; et al. Vascular endothelial growth factor gene polymorphisms and intracranial aneurysms. Acta Neurochir. (Wien) 2013, 155, 1511–1515. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yan, S.; Tan, H.; Ma, L.; Feng, H.; Han, H.; Pan, M.; Yu, L.; Fang, C. The miR-143/145 cluster reverses the regulation effect of KLF5 in smooth muscle cells with proliferation and contractility in intracranial aneurysm. Gene 2018, 679, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Zhang, Y.; Yu, J.; Chen, Y.; Gu, D.; Niu, C.; Fu, X.; Wei, J. Long Non-coding RNA MALAT1/microRNA-143/VEGFA Signal Axis Modulates Vascular Endothelial Injury-Induced Intracranial Aneurysm. Nanoscale Res. Lett. 2020, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Wei, C.; Li, K.; Lu, Z. LncRNA MALAT1 up-regulates VEGF-A and ANGPT2 to promote angiogenesis in brain microvascular endothelial cells against oxygen-glucose deprivation via targetting miR-145. Biosci. Rep. 2019, 39, BSR20180226. [Google Scholar] [CrossRef]
- Ma, S.C.; Li, Q.; Peng, J.Y.; Zhouwen, J.L.; Zhang, D.N.; Zhang, C.B.; Jiang, W.G.; Jia, W. CLDN5 affects lncRNAs acting as ceRNA dynamics contributing to regulating blood-brain barrier permeability in tumor brain metastasis. Oncol. Rep. 2018, 39, 1441–1453. [Google Scholar] [CrossRef]
- Uthaya Kumar, D.B.; Williams, A. Long non-coding RNAs in immune regulation and their potential as therapeutic targets. Int. Immunopharmacol. 2020, 81, 106279. [Google Scholar] [CrossRef]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef]
- Shen, J.; Zhao, Z.; Shang, W.; Liu, C.; Zhang, B.; Xu, Z.; Cai, H. Fabrication of a nano polymer wrapping Meg3 ShRNA plasmid for the treatment of cerebral infarction. Artif. Cells Nanomed. Biotechnol. 2018, 46, 894–903. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| LncRNA | Type of Aneurysm | Gene-Target | Related Functions | Sample Studied | Cellular Origin | Regulation | References |
|---|---|---|---|---|---|---|---|
| LOXL1-AS | TAA | Giver | Promotes apoptosis and inhibit VSMCs proliferation | HAoSMCs, human aorta tissue | VSMCs | Up | [6] |
| HOTAIR | TAA | ALOX15B, AZU1, PIP, TBX1, VIL1, KRT20, PRAMEF22 MMP-8, KERA, CCBE1 | Promotes apoptosis and inhibit VSMCs proliferation ECM remodeling (downregulates collagen type I and III expressions) | HAoSMCs, human aorta tissue, CABG tissue | VSMCs | Down | [7] |
| AK056155 | AAA | TGF-β, PI3K/Akt | Development of LDS and aortic aneurysm | HUVECs, human serum | ECs | Up | [8] |
| HLTF-5 | TAA | MMP9 | ECM remodeling, VSMC phenotype switching | Human aorta tissue | VSMCs | Up | [9] |
| MIAT | TAA | MiR-145, PI3K/Akt/Bcl-2 | Improves the viability and inhibits VSMCs apoptosis | Primary hVSMCs, human aorta tissue | VSMCs | Up | [10] |
| MALAT1 | AAA | NADPH NOX2 and IL-6 | Promotes AAA development | HAoSMCs, human aorta tissue | ECs | Up | [11] |
| HIF1A-AS1A | TAAA | Caspase-3, caspase-8, and Bcl2 | Reduces apoptosis of VSMCs | HAoSMCs, human serum | VSMCs | Up | [12,13] |
| MALAT1 | TAA | HDAC9 and BRG1 | VSMCs dysfunction and ECM disruption (increase the activity of MMP2 and MMP9) | Primary hVSMCs, human aorta tissue | VSMCs | Up | [14] |
| PVT1 | AAA | MMP2, MMP9, TNF-α, IL-1β, and IL-6 | Suppresses VSMCs apoptosis, ECM disruption (decrease the activity of MMP2 and MMP9), and serum pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) | Primary mVSMCs, human and mouse aorta tissue | VSMCs | Down | [15] |
| HIF1A-AS1 | TAA | BRG1 | Promotes apoptosis and inhibit VSMCs proliferation | Primary hVSMCs, human aorta tissue | VSMCs | Up | [16] |
| GAS5 | AAA | MiR-21 /PTEN/Akt, YBX1 | Promotes apoptosis and inhibit VSMCs proliferation | HAoSMCs, human and mouse aorta tissue | VSMCs | Up | [17] |
| H19 | AAA | HIF1α | Promotes apoptosis and inhibit VSMCs proliferation | HAoSMCs, human and mouse aorta tissue | VSMCs | Up | [18] |
| H19 | AAA | Let-7a, MCR-1, IL-6, | Promotes aneurysm formation by enhancing vascular pro-inflammatory cytokines and enhancing macrophage infiltration. | Primary mVSMCs and mouse macrophages, human and mouse aorta tissue | VSMCs | Up | [19] |
| LncRNA | Gene-Target | Disease | Regulation | Regulatory Effect of lncRNA | References |
|---|---|---|---|---|---|
| LEF1-AS1 | miR-544a/PTEN axis | Atherosclerosis | Up | Regulates VSMCs proliferation and migration | [20] |
| lncRNA 430945 | ROR2/RhoA | Atherosclerosis | Up | Promotes the proliferation and migration of VSMCs | [21] |
| lncRNA-ATB | TGF-β1 and caspase-3 | Atherosclerosis | Up | Promotes apoptosis and inhibits proliferation of ECs | [22] |
| lncRNA AF131217.1 | miR-128-3p/KLF4 axis | Atherosclerosis | Down | Reduces endothelial inflammation | [23] |
| lncRNA AK094457 | PPARγ | Hypertension | Up | Increase Ang II-induced hypertension and endothelial dysfunction | [24] |
| MALAT1 | Notch-1 | Hypertension | Down | Alleviates the vascular lesion and remodeling: reduction relative expression of inflammation-related, endothelial function-related and oxidative stress-related factors; inhibit ECs apoptosis | [25] |
| MRAK048635_P1 | Cyclin D1/E, CDK2/4, p-Rb, caspase3, PARP, α-SMA and calponin | Hypertension | Down | Induces VSMCs phenotypic switching from a contractile to a secretory phenotype. Promotes proliferation, migration and inhibits apoptosis of VSMCs | [26] |
| Disease | LncRNA | Sample | Regulation | Diagnostic | Prognostic | Sensitivity % | Specificity % | AUC | References |
|---|---|---|---|---|---|---|---|---|---|
| AIS | NEAT1 | Plasma | Up | Yes | Yes | 64.3 | 82.9 | 0.80 | [51] |
| AIS | ANRIL | Plasma | Down | Yes | No | 72.2 | 71.2 | 0.75 | [52] |
| AMI | H19, MALAT1 and MIAT | PBMC | Up | Yes | No | Comb. 0.68 | Comb. 0.76 | Comb. 0.76 | [53] |
| STEMI | LIPCAR | Plasma | Up | Yes | No | 0.82 | 0.75 | 0.78 | [54] |
| CAD | GAS5 | Plasma | Down | Yes | No | 0.78 | 0.92 | 0.97 | [55] |
| HF | LIPCAR | Plasma | Up | No | Yes | / | / | / | [56] |
| DCM | LncRNA ENST00000507296 | Plasma | Up | Yes | Yes | / | / | 0.78 | [57] |
| Athero-sclerosis | Exosomal HIF1A-AS1 | Plasma | Up | Yes | No | 0.86 | 0.82 | 0.75 | [58] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gareev, I.; Beylerli, O.; Aliev, G.; Pavlov, V.; Izmailov, A.; Zhang, Y.; Liang, Y.; Yang, G. The Role of Long Non-Coding RNAs in Intracranial Aneurysms and Subarachnoid Hemorrhage. Life 2020, 10, 155. https://doi.org/10.3390/life10090155
Gareev I, Beylerli O, Aliev G, Pavlov V, Izmailov A, Zhang Y, Liang Y, Yang G. The Role of Long Non-Coding RNAs in Intracranial Aneurysms and Subarachnoid Hemorrhage. Life. 2020; 10(9):155. https://doi.org/10.3390/life10090155
Chicago/Turabian StyleGareev, Ilgiz, Ozal Beylerli, Gjumrakch Aliev, Valentin Pavlov, Adel Izmailov, Yiwei Zhang, Yanchao Liang, and Guang Yang. 2020. "The Role of Long Non-Coding RNAs in Intracranial Aneurysms and Subarachnoid Hemorrhage" Life 10, no. 9: 155. https://doi.org/10.3390/life10090155
APA StyleGareev, I., Beylerli, O., Aliev, G., Pavlov, V., Izmailov, A., Zhang, Y., Liang, Y., & Yang, G. (2020). The Role of Long Non-Coding RNAs in Intracranial Aneurysms and Subarachnoid Hemorrhage. Life, 10(9), 155. https://doi.org/10.3390/life10090155