Contribution of Benthic Processes to the Growth of Ooids on a Low-Energy Shore in Cat Island, The Bahamas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Measurements and Petrographic Analyses
2.3. Grain Erosion Experiments
2.4. Hydro-Morphodynamic Model
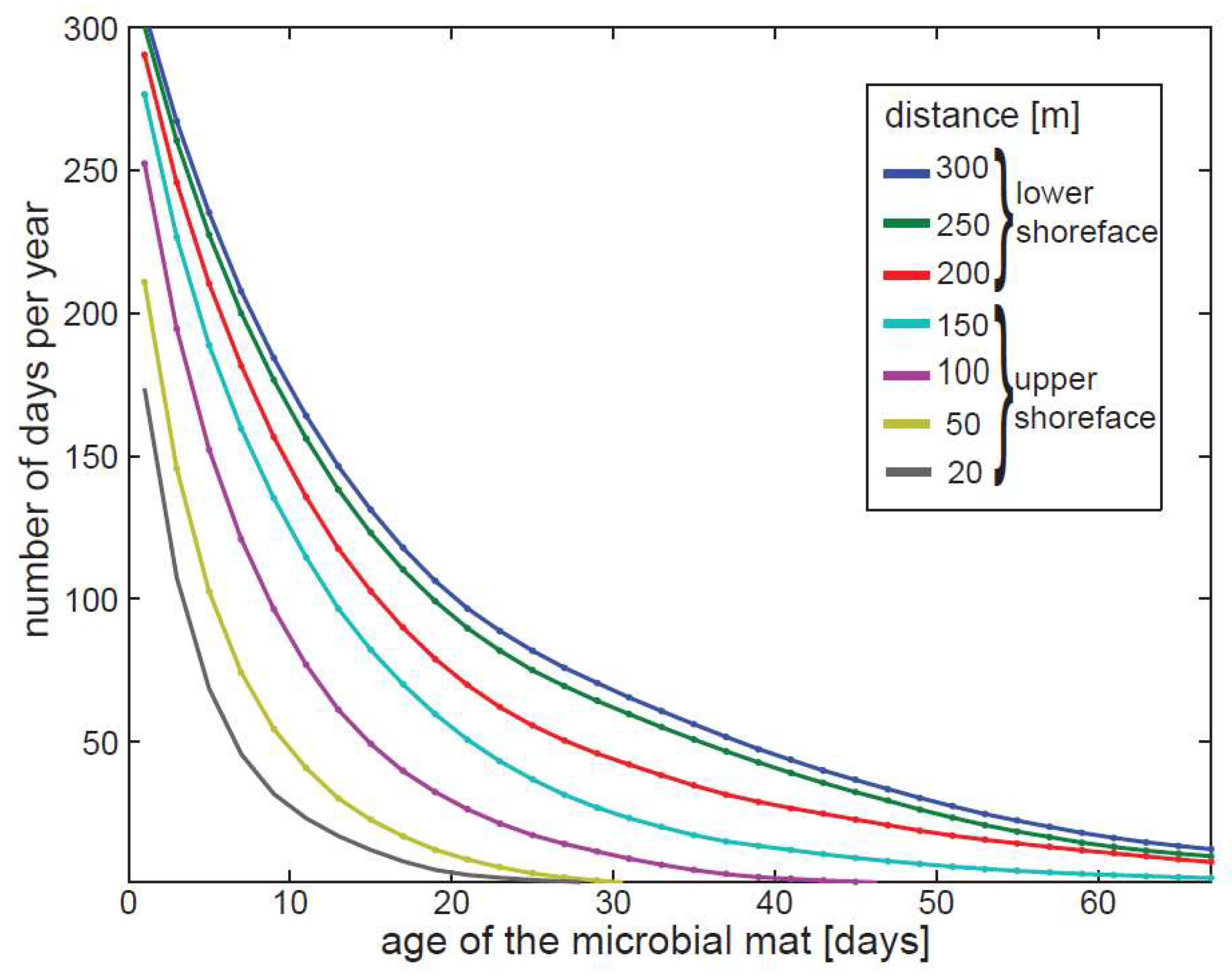
2.5. Modeling Microbial Colonization of Sediments
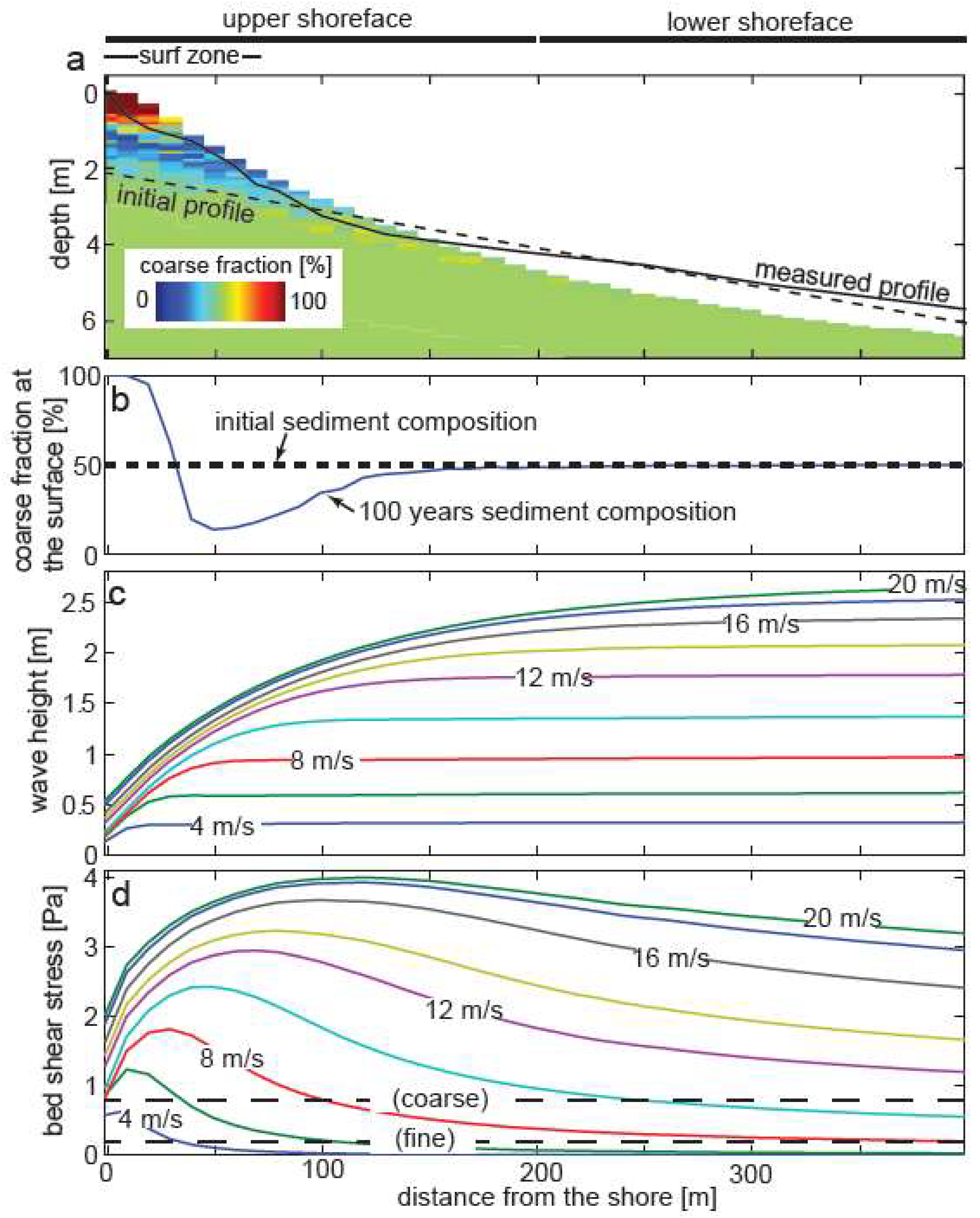
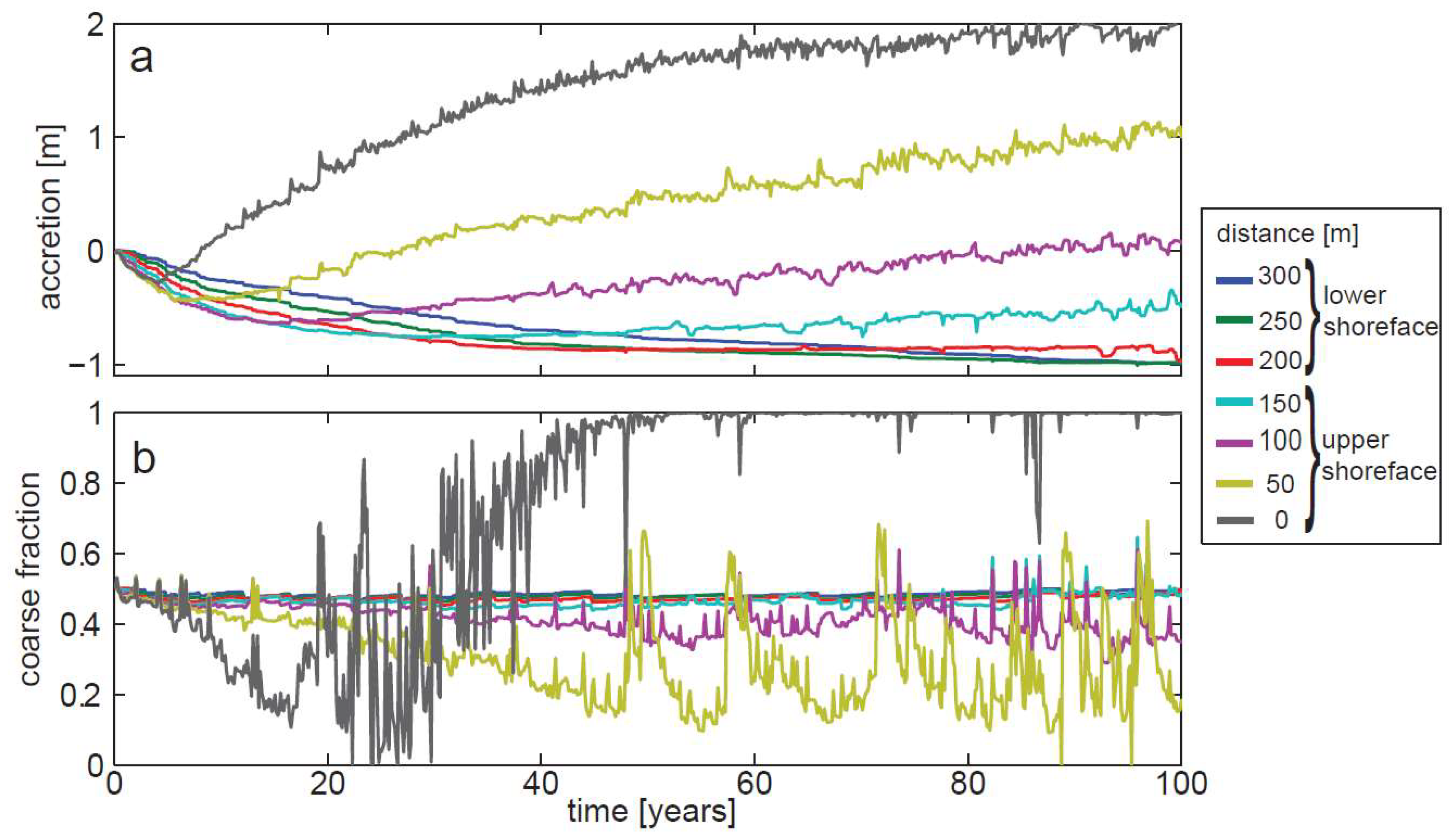
3. Results
3.1. Field Data and Petrographic Analyses
3.2. Model of Sediment Transport and Sorting in Pigeon Cay
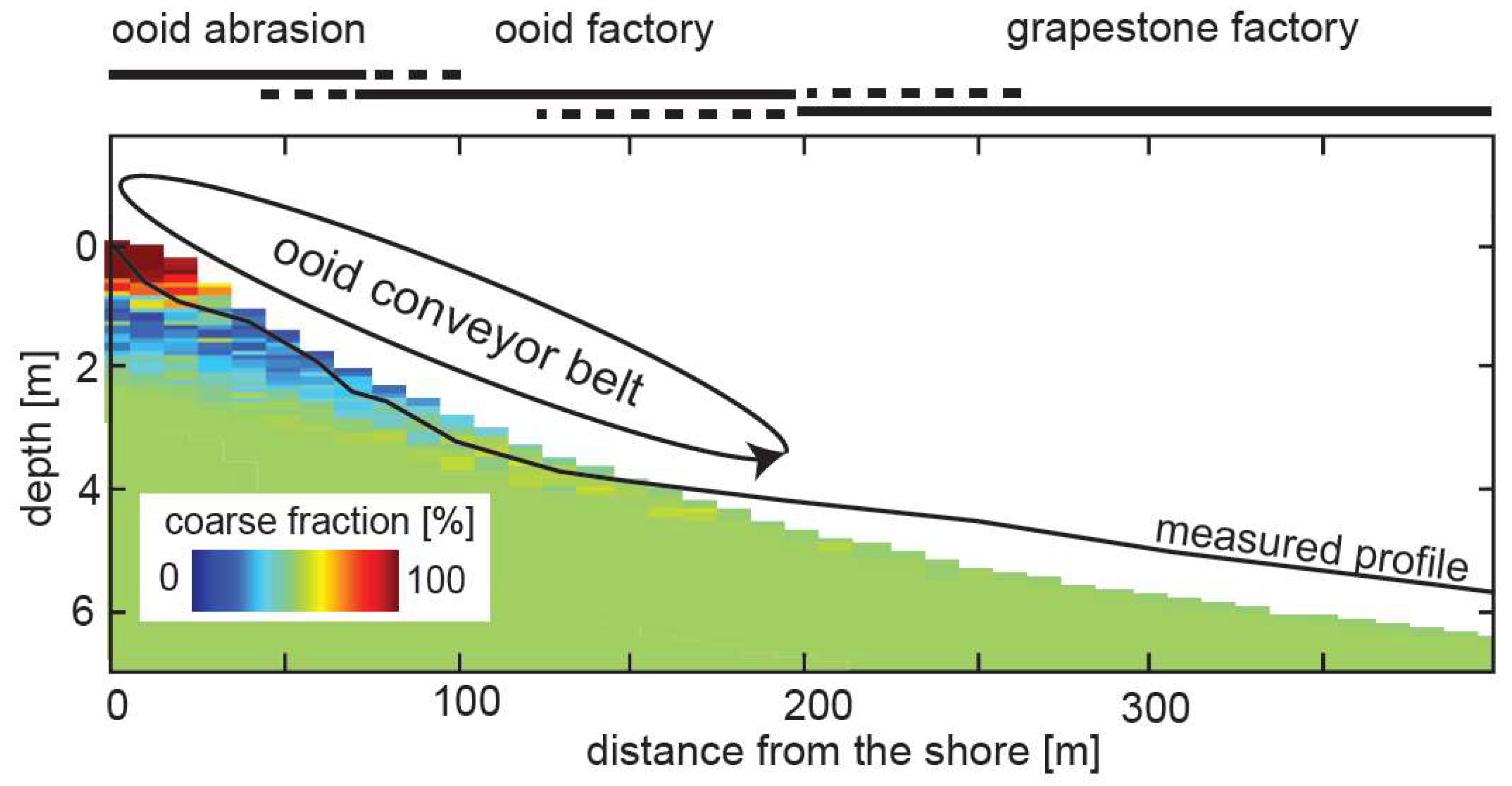
4. Discussion
4.1. Sedimentary Processes at Pigeon Cay
4.2. A Conceptual Model for Ooid Growth: The Conveyor-Belt
4.3. Biosignatures in Ooids
4.4. Petrographic Implications of Models for Suspended and Benthic Ooid Growth
4.5. Conditions for Benthic Ooid Growth at Other Sites
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Newell, N.D.; Purdy, E.G.; Imbrie, J. Bahamian oölitic sand. J. Geol. 1960, 68, 481–497. [Google Scholar] [CrossRef]
- Heller, P.L.; Komar, P.D.; Pevear, D.R. Transport processes in ooid genesis. J. Sediment. Res. 1980, 50, 943–951. [Google Scholar] [CrossRef]
- Given, R.K.; Wilkinson, B.H. Kinetic control of morphology, composition, and mineralogy of abiotic sedimentary carbonates. J. Sediment. Res. 1985, 55, 109–119. [Google Scholar] [CrossRef]
- Strasser, A. Ooids in Purbeck limestones (lowermost Cretaceous) of the Swiss and French Jura. Sedimentology 1986, 33, 711–727. [Google Scholar] [CrossRef]
- Davies, P.J.; Bubela, B.; Ferguson, J. The formation of ooids. Sedimentology 1978, 25, 703–730. [Google Scholar] [CrossRef]
- Ferguson, J.; Bubela, B.; Davies, P.J. Synthesis and possible mechanism of formation of radial carbonate ooids. Chem. Geol. 1978, 22, 285–308. [Google Scholar] [CrossRef]
- Duguid, S.M.A.; Kyser, T.K.; James, N.P.; Rankey, E.C. Microbes and ooids. J. Sediment. Res. 2010, 80, 236–251. [Google Scholar] [CrossRef]
- Trower, E.J.; Lamb, M.P.; Fischer, W.W. Experimental evidence that ooid size reflects a dynamic equilibrium between rapid precipitation and abrasion rates. Earth Planet. Sci. Lett. 2017, 468, 112–118. [Google Scholar] [CrossRef]
- Sumner, D.Y.; Grotzinger, J.P. Numerical modeling of ooid size and the problem of Neoproterozoic giant ooids. J. Sediment. Res. 1993, 63, 974–982. [Google Scholar]
- Diaz, M.R.; Eberli, G.P.; Blackwelder, P.; Phillips, B.; Swart, P.K. Microbially mediated organomineralization in the formation of ooids. Geology 2017, 45, 771–774. [Google Scholar] [CrossRef]
- Batchelor, M.T.; Burne, R.V.; Henry, B.I.; Li, F.; Paul, J. A biofilm and organomineralisation model for the growth and limiting size of ooids. Sci. Rep. 2018, 8, 559. [Google Scholar] [CrossRef] [PubMed]
- Mitterer, R.M. Aminoacid composition of organic matrix in calcareous oolites. Science 1968, 162, 1498–1499. [Google Scholar] [CrossRef] [PubMed]
- Mitterer, R.M. Biogeochemistry of aragonite mud and oolites. Geochim. Cosmochim. Acta 1972, 36, 1407–1422. [Google Scholar] [CrossRef]
- Land, L.S.; Behrens, E.W.; Frishman, S.A. The ooids of Baffin Bay, Texas. J. Sediment. Res. 1979, 49, 1269–1277. [Google Scholar] [CrossRef]
- Reitner, J.; Arp, G.; Thiel, V.; Gautret, P.; Galling, U.; Michaelis, W. Organic matter in Great Salt Lake ooids (Utah, USA)—First approach to a formation via organic matrices. Facies 1997, 36, 210–219. [Google Scholar]
- Folk, R.L.; Leo Lynch, F. Organic matter, putative nannobacteria and the formation of ooids and hardgrounds. Sedimentology 2001, 48, 215–229. [Google Scholar] [CrossRef]
- Summons, R.E.; Bird, L.R.; Gillespie, A.L.; Pruss, S.B.; Roberts, M.; Sessions, A.L. Lipid biomarkers in ooids from different locations and ages: Evidence for a common bacterial flora. Geobiology 2013, 11, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Bathurst, R.G.C. Carbonate Sediments and Their Diagenesis. In Developments in Sedimentology; Elsevier: Amsterdam, The Netherlands, 1971; Volume 12. [Google Scholar]
- Wanless, H.R.; Tedesco, L.P. Comparison of oolitic sand bodies generated by tidal vs. wind-wave agitation: Chapter 15. AAPG Spec. Vol. 1993, 48, 199–225. [Google Scholar]
- Rankey, E.C.; Riegl, B.; Steffen, K. Form, function and feedbacks in a tidally dominated ooid shoal, Bahamas. Sedimentology 2006, 53, 1191–1210. [Google Scholar] [CrossRef]
- Gebelein, C.D. Distribution, morphology, and accretion rate of recent subtidal algal stromatolites, Bermuda. J. Sediment. Res. 1969, 39, 49–69. [Google Scholar] [CrossRef]
- Neumann, A.C.; Gebelein, C.D.; Scoffin, T.P. The composition, structure and erodability of subtidal mats, Abaco, Bahamas. J. Sediment. Res. 1970, 40, 274–297. [Google Scholar] [CrossRef]
- Mariotti, G.; Perron, J.T.; Bosak, T. Feedbacks between flow, sediment motion and microbial growth on sand bars initiate and shape elongated stromatolite mounds. Earth Planet. Sci. Lett. 2014, 397, 93–100. [Google Scholar] [CrossRef]
- Visscher, P.T.; Reid, R.P.; Bebout, B.M. Microscale observations of sulfate reduction: Correlation of microbial activity with lithified micritic laminae in modern marine stromatolites. Geology 2000, 28, 919–922. [Google Scholar] [CrossRef]
- Camoin, G.F.; Montaggioni, L.F. High energy coralgal-stromatolite frameworks from Holocene reefs (Tahiti, French Polynesia). Sedimentology 1994, 41, 655–676. [Google Scholar] [CrossRef]
- Birgel, D.; Meister, P.; Lundberg, R.; Horath, T.D.; Bontognali, T.R.R.; Bahniuk, A.M.; de Rezende, C.E.; Vasconcelos, C.; McKenzie, J.A. Methanogenesis produces strong 13C enrichment in stromatolites of Lagoa Salgada, Brazil: A modern analogue for Palaeo-/Neoproterozoic stromatolites? Geobiology 2015, 13, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.S.; Sumner, D.Y.; Reid, R.P.; Swart, P.K. Isotopic fingerprints of microbial respiration in aragonite from Bahamian stromatolites. Geology 2006, 34, 973–976. [Google Scholar] [CrossRef]
- Swart, P.K.; Reijmer, J.J.G.; Otto, R. A re-evaluation of facies on great Bahama Bank II: Variations in the δ13C, δ18O and mineralogy of surface sediments. In Perspectives in Carbonate Geology; Swart, P.K., Eberli, G.P., McKenzie, J.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2009; pp. 47–59. ISBN 978-1-4443-1206-5. [Google Scholar]
- Purdy, E. Recent calcium carbonate facies of the Great Bahama Bank. 2. Sedimentary facies. J. Geol. 1963, 71, 472–497. [Google Scholar] [CrossRef]
- Winland, H.D.; Matthews, R.K. Origin and significance of grapestone, Bahama Islands. J. Sediment. Res. 1974, 44, 921–927. [Google Scholar]
- Fabricius, F.H. Origin of Marine Ooids and Grapestones. In Contribution to Sedimentology; Schweizerbart: Stuttgart, Germany, 1977; ISBN 978-3-510-57007-2. [Google Scholar]
- Gaffey, S.J. Formation and infilling of pits in marine ooid surfaces. J. Sediment. Res. 1983, 53, 193–208. [Google Scholar]
- Tucker, M.E.; Wright, P. Carbonate Sedimentology; Wiley: Chichester, UK, 1990; ISBN 978-0-632-01472-9. [Google Scholar]
- Haq, B.U.; Boersma, A. Introduction to Marine Micropaleontology; Elsevier Science: Amsterdam, The Netherlands, 1998; ISBN 978-0-444-82672-5. [Google Scholar]
- Bathurst, R.G.C. Oölitic films on low energy carbonate sand grains, Bimini Lagoon, Bahamas. Mar. Geol. 1967, 5, 89–109. [Google Scholar] [CrossRef]
- Dravis, J. Rapid and widespread generation of recent oolitic hardgrounds on a high energy Bahamian platform, Eleuthera Bank, Bahamas. J. Sediment. Res. 1979, 49, 195–207. [Google Scholar] [CrossRef]
- Husinec, A.; Read, J.F. Transgressive oversized radial ooid facies in the Late Jurassic Adriatic Platform interior: Low-energy precipitates from highly supersaturated hypersaline waters. GSA Bull. 2006, 118, 550–556. [Google Scholar] [CrossRef]
- O’Reilly, S.S.; Mariotti, G.; Winter, A.R.; Newman, S.A.; Matys, E.D.; McDermott, F.; Pruss, S.B.; Bosak, T.; Summons, R.E.; Klepac-Ceraj, V. Molecular biosignatures reveal common benthic microbial sources of organic matter in ooids and grapestones from Pigeon Cay, The Bahamas. Geobiology 2016, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Beaupré, S.R.; Roberts, M.L.; Burton, J.R.; Summons, R.E. Rapid, high-resolution 14C chronology of ooids. Geochim. Cosmochim. Acta 2015, 159, 126–138. [Google Scholar] [CrossRef]
- Dominguez, L.L.; Mullins, H.T.; Hine, A.C. Cat Island platform, Bahamas: An incipiently drowned Holocene carbonate shelf. Sedimentology 1988, 35, 805–819. [Google Scholar] [CrossRef]
- Rankey, E.C. Contrasts between wave- and tide-dominated oolitic systems: Holocene of Crooked–Acklins Platform, southern Bahamas. Facies 2013, 60, 405–428. [Google Scholar] [CrossRef]
- Blott, S.J.; Pye, K. Gradistat: A grain size distribution and statistics package for the analysis of unconsolidated sediments. Earth Surf. Process. Landf. 2001, 26, 1237–1248. [Google Scholar] [CrossRef]
- Francus, P. An image-analysis technique to measure grain-size variation in thin sections of soft clastic sediments. Sediment. Geol. 1998, 121, 289–298. [Google Scholar] [CrossRef]
- Wadell, H. Volume, shape, and roundness of rock particles. J. Geol. 1932, 40, 443–451. [Google Scholar] [CrossRef]
- Lesser, G.R.; Roelvink, J.A.; van Kester, J.A.T.M.; Stelling, G.S. Development and validation of a three-dimensional morphological model. Coast. Eng. 2004, 51, 883–915. [Google Scholar] [CrossRef]
- Soulsby, R.L. Dynamics of Marine Sands; Thomas Telford Publishing: London, UK, 1997. [Google Scholar]
- Herterich, K.; Hasselmann, K. The horizontal diffusion of tracers by surface waves. J. Phys. Oceanogr. 1982, 12, 704–711. [Google Scholar] [CrossRef]
- Booij, N.; Ris, R.C.; Holthuijsen, L.H. A third-generation wave model for coastal regions—1. Model description and validation. J. Geophys. Res. Oceans 1999, 104, 7649–7666. [Google Scholar] [CrossRef]
- Cavaleri, L.; Rizzoli, P.M. Wind wave prediction in shallow water: Theory and applications. J. Geophys. Res. Oceans 1981, 86, 10961–10973. [Google Scholar] [CrossRef]
- Komen, G.J.; Hasselmann, K.; Hasselmann, K. On the existence of a fully developed wind-sea spectrum. J. Phys. Oceanogr. 1984, 14, 1271–1285. [Google Scholar] [CrossRef]
- Collins, J.I. Prediction of shallow-water spectra. J. Geophys. Res. 1972, 77, 2693–2707. [Google Scholar] [CrossRef]
- Battjes, J.A.; Janssen, J.P.F.M. Energy loss and set-up due to breaking of random waves. Coast. Eng. Proc. 2011, 1, 569–587. [Google Scholar] [CrossRef]
- Hasselmann, S.; Hasselmann, K. Computations and parameterizations of the nonlinear energy transfer in a gravity-wave spectrum. Part I: A new method for efficient computations of the exact nonlinear transfer integral. J. Phys. Oceanogr. 1985, 15, 1369–1377. [Google Scholar] [CrossRef]
- Bijker, E.W. Some Considerations about Scales for Coastal Models with Movable Bed; Delft Hydraulics Laboratory: Delft, The Netherlands, 1967. [Google Scholar]
- Bailard, J.A. An energetics total load sediment transport model for a plane sloping beach. J. Geophys. Res. Oceans 1981, 86, 10938–10954. [Google Scholar] [CrossRef]
- Roelvink, J.A. Coastal morphodynamic evolution techniques. Coast. Eng. 2006, 53, 277–287. [Google Scholar] [CrossRef]
- Mariotti, G.; Fagherazzi, S. Modeling the effect of tides and waves on benthic biofilms. J. Geophys. Res. Biogeosci. 2012, 117, 1–14. [Google Scholar] [CrossRef]
- Cowell, P.J.; Stive, M.J.F.; Niedoroda, A.W.; de Vriend, H.J.; Swift, D.J.P.; Kaminsky, G.M.; Capobianco, M. The coastal-tract (Part 1): A conceptual approach to aggregated modeling of low-order coastal change. J. Coast. Res. 2003, 19, 812–827. [Google Scholar]
- Stive, M.J.F.; de Vriend, H.J. Modelling shoreface profile evolution. Mar. Geol. 1995, 126, 235–248. [Google Scholar] [CrossRef]
- Paterson, D.M.; Aspden, R.J.; Visscher, P.T.; Consalvey, M.; Andres, M.S.; Decho, A.W.; Stolz, J.; Reid, R.P. Light-dependant biostabilisation of sediments by stromatolite assemblages. PLoS ONE 2008, 3, e3176. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.C.; Ashton, A.D. Exploring shoreface dynamics and a mechanistic explanation for a morphodynamic depth of closure. J. Geophys. Res. Earth Surf. 2016, 121, 442–464. [Google Scholar] [CrossRef]
- Larson, M.; Kraus, N.C. Prediction of cross-shore sediment transport at different spatial and temporal scales. Mar. Geol. 1995, 126, 111–127. [Google Scholar] [CrossRef]
- Reid, D.R.P.; Macintyre, D.I.G.; Browne, D.K.M.; Steneck, D.R.S.; Miller, D.T. Modern marine stromatolites in the Exuma Cays, Bahamas: Uncommonly common. Facies 1995, 33, 1–17. [Google Scholar] [CrossRef]
- Reid, R.P.; Visscher, P.T.; Decho, A.W.; Stolz, J.F.; Bebout, B.M.; Dupraz, C.; Macintyre, I.G.; Paerl, H.W.; Pinckney, J.L.; Prufert-Bebout, L.; et al. The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 2000, 406, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Van Ee, N.J.; Wanless, H.R. Ooids and grapestone—A significant source of mud on Caicos platform. In Developing Models and Analogs for Isolated Carbonate Platforms-Holocene and Pleistocene Carbonate s of Caicos Platform, British West Indies; Morgan, W.A., Harris, P.M., Eds.; SEPM Society for Sedimentary Geology: Broken Arrow, OK, USA, 2008; pp. 121–125. [Google Scholar]
- Freeman, T. Quiet water oolites from Laguna Madre, Texas. J. Sediment. Res. 1962, 32, 475–483. [Google Scholar] [CrossRef]
- Komar, P.D. Beach Processes and Sedimentation; Prentice-Hall: Upper Saddle River, NJ, USA, 1976; ISBN 978-0-13-072595-0. [Google Scholar]
- Woodroffe, C.D. Coasts: Form, Process and Evolution; Cambridge University Press: Cambridge, UK, 2002; ISBN 978-0-521-01183-9. [Google Scholar]
- Harris, P.M.; Halley, R.B.; Lukas, K.J. Endolith microborings and their preservation in Holocene-Pleistocene (Bahama-Florida) ooids. Geology 1979, 7, 216–220. [Google Scholar] [CrossRef]
- Diaz, M.R.; Norstrand, J.D.; Eberli, G.P.; Piggot, A.M.; Zhou, J.; Klaus, J.S. Functional gene diversity of oolitic sands from Great Bahama Bank. Geobiology 2014, 12, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Medwedeff, D.A.; Wilkinson, B.H. Cortical fabrics in calcite and aragonite ooids. In Coated Grains; Peryt, D.T.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 109–115. ISBN 978-3-642-68871-3. [Google Scholar]
- Wilkinson, B.H.; Buczynski, C.; Owen, R.M. Chemical control of carbonate phases; implications from Upper Pennsylvanian calcite-aragonite ooids of southeastern Kansas. J. Sediment. Res. 1984, 54, 932–947. [Google Scholar] [CrossRef]
- Flügel, E. Microfacies of Carbonate Rocks; Springer: Berlin/Heidelberg, Germany, 2004; ISBN 978-3-662-08728-2. [Google Scholar]
- Simone, L. Ooids: A review. Earth-Sci. Rev. 1980, 16, 319–355. [Google Scholar] [CrossRef]
- Gerdes, G.; Dunajtschik-Piewak, K.; Riege, H.; Taher, A.G.; Krumbein, W.E.; Reineck, H.-E. Structural diversity of biogenic carbonate particles in microbial mats. Sedimentology 1994, 41, 1273–1294. [Google Scholar] [CrossRef]
- Cayeux, L. Les Roches Sédimentaires de France. Roches Carbonatées; Impr. Nationale: Paris, France, 1929. [Google Scholar]
- Li, S.; Wang, Z.J.; Chang, T.-T. Temperature oscillation modulated self-assembly of periodic concentric layered magnesium carbonate microparticles. PLoS ONE 2014, 9, e88648. [Google Scholar] [CrossRef] [PubMed]
- Beukes, N.J. Ooids and Oolites of the Proterophytic Boomplaas Formation, Transvaal Supergroup, Griqualand West, South Africa. In Coated Grains; Peryt, D.T.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 199–214. ISBN 978-3-642-68871-3. [Google Scholar]
- Swett, K.; Knoll, A.H. Marine pisolites from Upper Proterozoic carbonates of East Greenland and Spitsbergen. Sedimentology 1989, 36, 75–93. [Google Scholar] [CrossRef] [PubMed]
- Swirydczuk, K.; Wilkinson, B.H.; Smith, G.R. The Pliocene Glenns Ferry Oolite: Lake-margin carbonate deposition in the Southwestern Snake River Plain. J. Sediment. Petrol. 1979, 49, 995–1004. [Google Scholar]
- Davaud, E.; Girardclos, S. Recent freshwater ooids and oncoids from Western Lake Geneva (Switzerland): Indications of a common organically mediated origin. J. Sediment. Res. 2001, 71, 423–429. [Google Scholar] [CrossRef]
- Talbot, M.R. Foraminiferid-encrusted ooids: Evidence for ancient subtidal algal mats? Geol. Mag. 1974, 111, 297–302. [Google Scholar] [CrossRef]
- Wilson, R.C.L. Carbonate facies variation within the Osmington Oolite series in Southern England. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1968, 4, 89–123. [Google Scholar] [CrossRef]
- Siahi, M.; Hoffman, A.; Master, S.; Mueller, C.W.; Gerdes, A. Carbonate ooids of the mesoarchean pongola group, South Africa. Geobiology 2017, 15, 750–766. [Google Scholar] [CrossRef] [PubMed]
- Sipos, A.A.; Domokos, G.; Jerolmack, D.J. Shape evolution of ooids: A geometric model. Sci. Rep. 2018, 8, 1758. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.R.; Piggot, A.M.; Eberli, G.P.; Klaus, J.S. Bacterial community of oolitic carbonate sediments of the Bahamas Archipelago. Mar. Ecol. Prog. Ser. 2013, 485, 9–24. [Google Scholar] [CrossRef]
- Hine, A.C. Lily Bank, Bahamas; history of an active oolite sand shoal. J. Sediment. Res. 1977, 47, 1554–1581. [Google Scholar] [CrossRef]
- Wilkinson, B.H.; Owen, R.M.; Carroll, A.R. Submarine hydrothermal weathering, global eustasy, and carbonate polymorphism in Phanerozoic marine oolites. J. Sediment. Res. 1985, 55, 171–183. [Google Scholar] [CrossRef]
- Lehrmann, D.J.; Minzoni, M.; Li, X.; Yu, M.; Payne, J.L.; Kelley, B.M.; Schaal, E.K.; Enos, P. Lower triassic oolites of the Nanpanjiang Basin, South China: Facies architecture, giant ooids, and diagenesis-implications for hydrocarbon reservoirs. Aapg Bull. 2012, 96, 1389–1414. [Google Scholar] [CrossRef]
- Woods, A.D. Microbial ooids and cortoids from the Lower Triassic (Spathian) Virgin Limestone, Nevada, USA: Evidence for an Early Triassic microbial bloom in shallow depositional environments. Glob. Planet. Chang. 2013, 105, 91–101. [Google Scholar] [CrossRef]











© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariotti, G.; Pruss, S.B.; Summons, R.E.; Newman, S.A.; Bosak, T. Contribution of Benthic Processes to the Growth of Ooids on a Low-Energy Shore in Cat Island, The Bahamas. Minerals 2018, 8, 252. https://doi.org/10.3390/min8060252
Mariotti G, Pruss SB, Summons RE, Newman SA, Bosak T. Contribution of Benthic Processes to the Growth of Ooids on a Low-Energy Shore in Cat Island, The Bahamas. Minerals. 2018; 8(6):252. https://doi.org/10.3390/min8060252
Chicago/Turabian StyleMariotti, Giulio, Sara B. Pruss, Roger E. Summons, Sharon A. Newman, and Tanja Bosak. 2018. "Contribution of Benthic Processes to the Growth of Ooids on a Low-Energy Shore in Cat Island, The Bahamas" Minerals 8, no. 6: 252. https://doi.org/10.3390/min8060252
APA StyleMariotti, G., Pruss, S. B., Summons, R. E., Newman, S. A., & Bosak, T. (2018). Contribution of Benthic Processes to the Growth of Ooids on a Low-Energy Shore in Cat Island, The Bahamas. Minerals, 8(6), 252. https://doi.org/10.3390/min8060252