
Complex Biomineralization Pathways of the Belemnite Rostrum Cause Biased Paleotemperature Estimates

 , , ,
, , ,  , , and
, , and 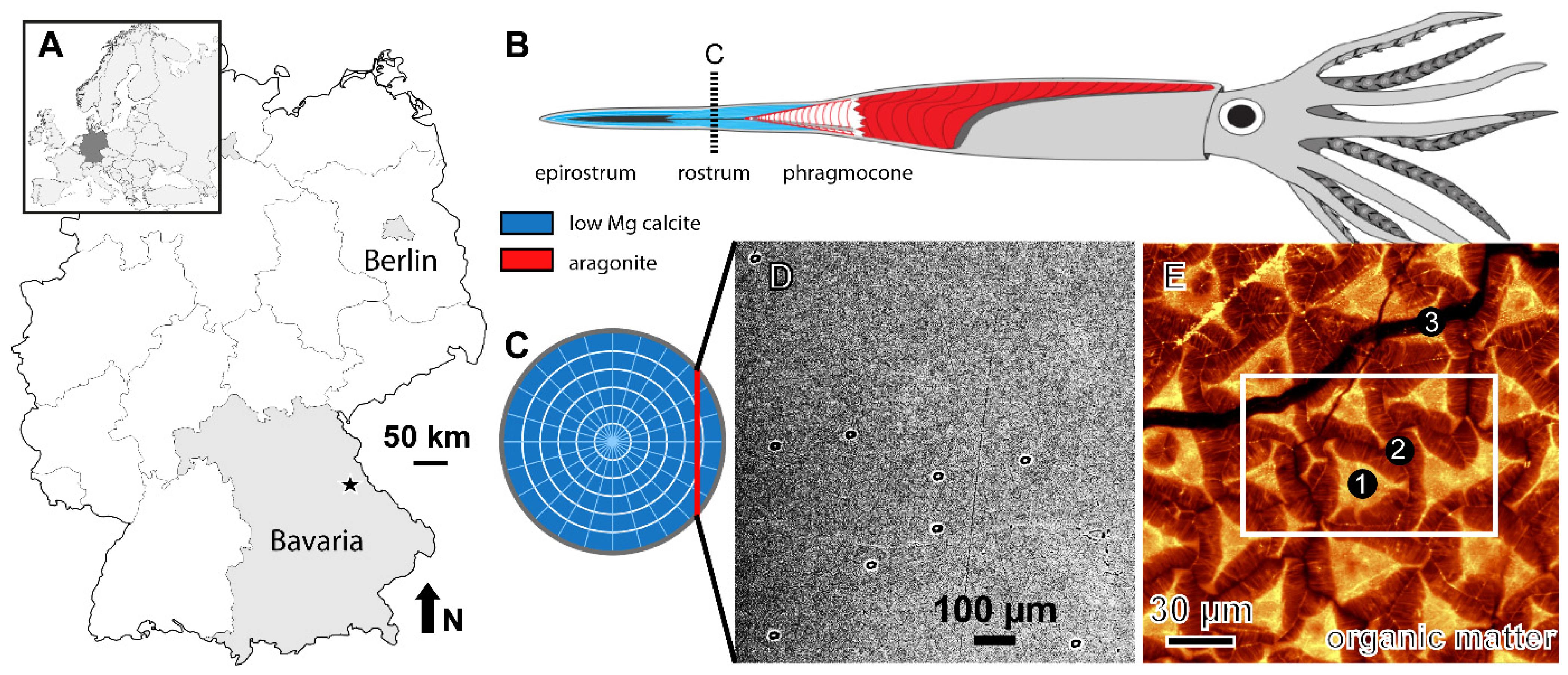{kind=link}
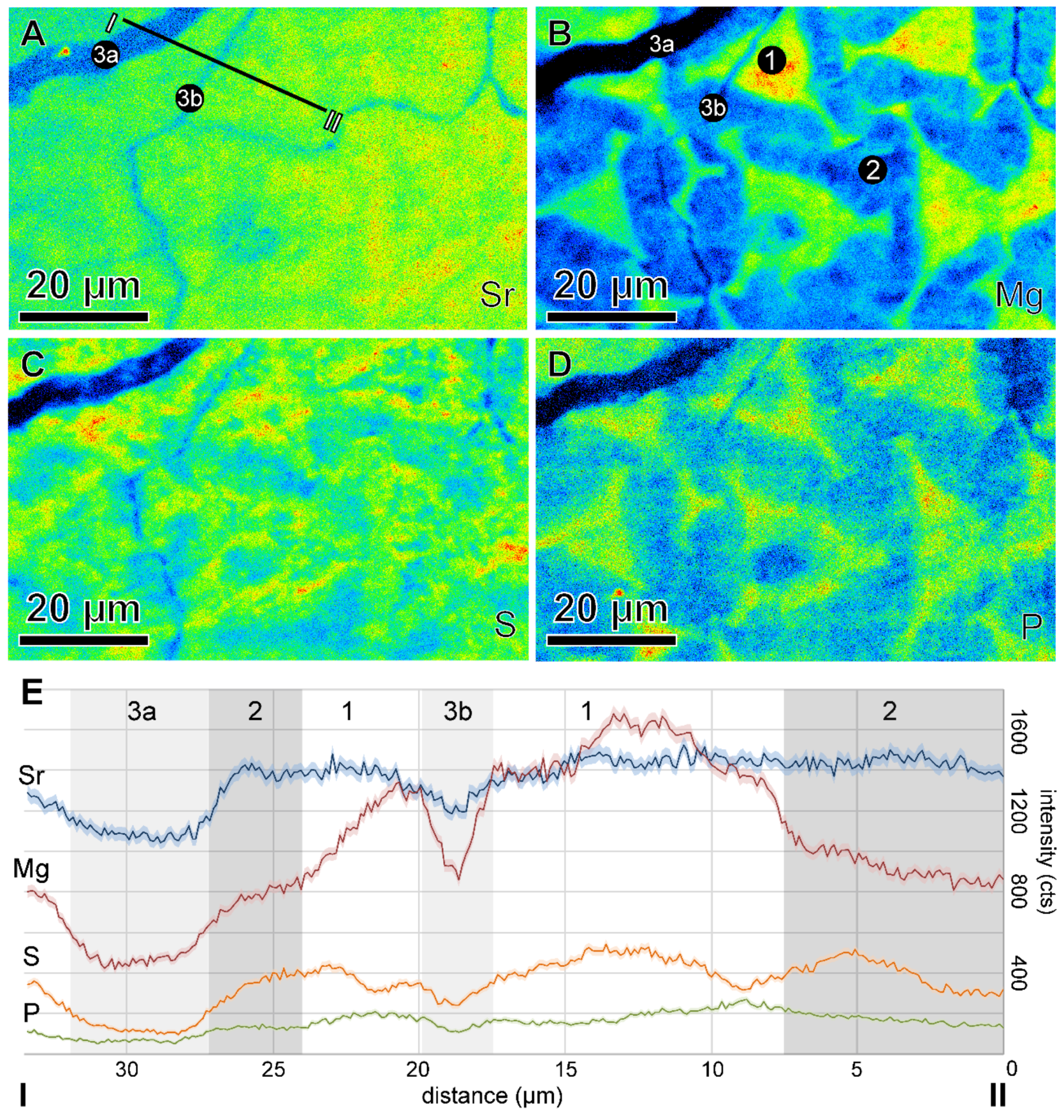{kind=link}
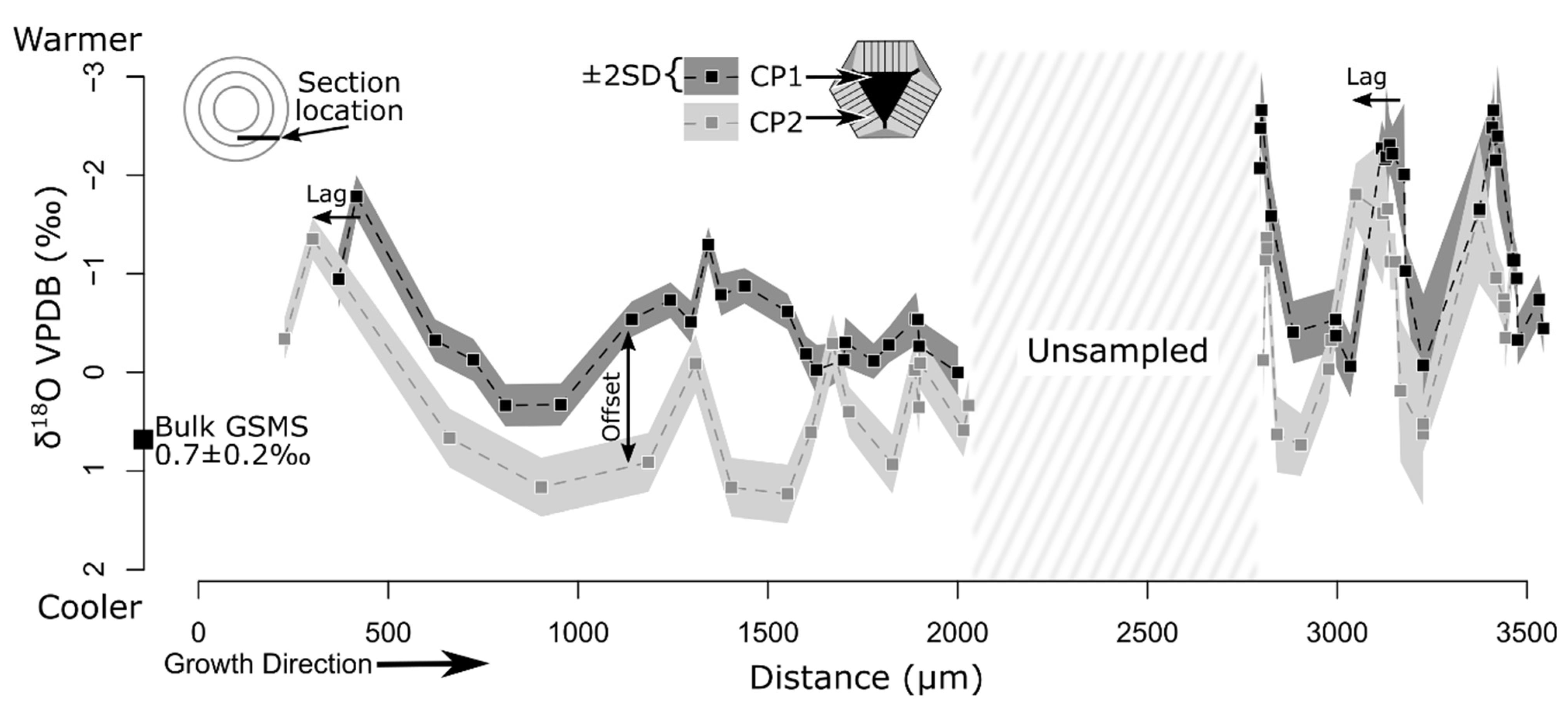{kind=link}
{kind=link}
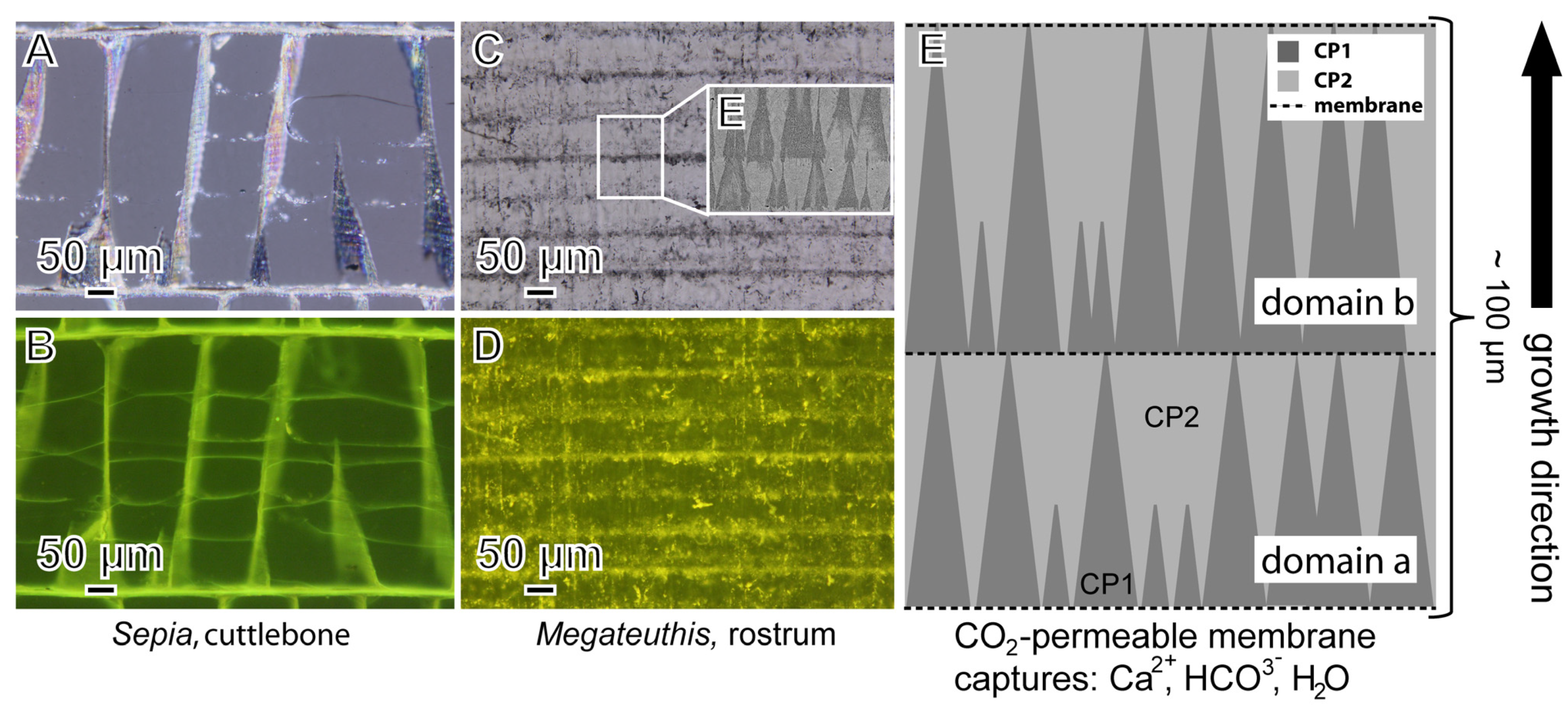{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
4.1. SIMS Accuracy
4.2. Oxygen Isotope Fractionation between Calcite and Water: Definitions and Range
4.3. Oxygen Isotope Differences in the Two Calcite Phases

4.4. Rostrum Biomineralization Model

4.5. Paleoceanographic Implications
4.6. Belemnite Paleobiology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Urey, H.C.; Lowenstam, H.A.; Epstein, S.; McKinney, C.R. Measurement of paleotemperatures and temperatures of the Upper Cretaceous of England, Denmark, and the Southeastern United States. GSA Bull. 1951, 62, 399–416. [Google Scholar] [CrossRef]
- McCrea, J.M. On the isotopic chemistry of carbonates and a paleotemperature scale. J. Chem. Phys. 1950, 18, 849–857. [Google Scholar] [CrossRef]
- O’Brien, C.L.; Robinson, S.A.; Pancost, R.D.; Sinninghe Damsté, J.S.; Schouten, S.; Lunt, D.J.; Alsenz, H.; Bornemann, A.; Bottini, C.; Brassell, S.C.; et al. Cretaceous sea-surface temperature evolution: Constraints from TEX86 and planktonic foraminiferal oxygen isotopes. Earth Sci. Rev. 2017, 172, 224–247. [Google Scholar] [CrossRef]
- Ivany, L.C. Reconstructing Paleoseasonality from Accretionary Skeletal Carbonates—Challenges and Opportunities. Paleontol. Soc. Pap. 2012, 18, 133–165. [Google Scholar] [CrossRef]
- Dietzel, M.; Tang, J.; Leis, A.; Köhler, S.J. Oxygen isotopic fractionation during inorganic calcite precipitation—Effects of temperature, precipitation rate and pH. Chem. Geol. 2009, 268, 107–115. [Google Scholar] [CrossRef]
- Mavromatis, V.; Schmidt, M.; Botz, R.; Comas-Bru, L.; Oelkers, E.H. Experimental quantification of the effect of Mg on calcite-aqueous fluid oxygen isotope fractionation. Chem. Geol. 2012, 310–311, 97–105. [Google Scholar] [CrossRef]
- Dietzel, M.; Purgstaller, B.; Kluge, T.; Leis, A.; Mavromatis, V. Oxygen and clumped isotope fractionation during the formation of Mg calcite via an amorphous precursor. Geochim. Cosmochim. Acta 2020, 276, 258–273. [Google Scholar] [CrossRef]
- Spero, H.J.; Lea, D.W. Experimental determination of stable isotope variability in Globigerina bulloides: Implications for paleoceanographic reconstructions. Mar. Micropaleontol. 1996, 28, 231–246. [Google Scholar] [CrossRef]
- Owen, R.; Kennedy, H.; Richardson, C. Isotopic partitioning between scallop shell calcite and seawater: Effect of shell growth rate. Geochim. Cosmochim. Acta 2002, 10, 1727–1737. [Google Scholar] [CrossRef]
- Stevens, K.; Griesshaber, E.; Schmahl, W.; Casella, L.A.; Iba, Y.; Mutterlose, J. Belemnite biomineralization, development, and geochemistry: The complex rostrum of Neohibolites minimus. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 468, 388–402. [Google Scholar] [CrossRef]
- Ullmann, C.V.; Korte, C. Diagenetic alteration in low-Mg calcite from macrofossils: A review. Geol. Q. 2015, 59, 3–20. [Google Scholar] [CrossRef]
- Immenhauser, A.; Schöne, B.R.; Hoffmann, R.; Niedermayr, A. Mollusc and brachiopod skeletal hard parts: Intricate archives of their marine environment. Sedimentology 2016, 63, 1–59. [Google Scholar] [CrossRef]
- Linzmeier, B.J. Refining the interpretation of oxygen isotope variability in free-swimming organisms. Swiss J. Palaeontol. 2019, 138, 109–121. [Google Scholar] [CrossRef]
- Sælen, G. Diagenesis and construction of the belemnite rostrum. Palaeontology 1989, 32, 765–798. [Google Scholar]
- Price, G.D.; Hart, M.B.; Wilby, P.R.; Page, K.N. Isotopic analysis of Jurassic (Callovian) mollusks from the Christian Malford Lagerstätte (UK): Implications for ocean water temperature estimates based on belemnoids. PALAIOS 2015, 30, 645–654. [Google Scholar] [CrossRef]
- Price, G.D.; Twitchett, R.J.; Wheeley, J.R.; Buono, G. Isotopic evidence for long term warmth in the Mesozoic. Sci. Rep. 2013, 3, 1438. [Google Scholar] [CrossRef]
- Veizer, J.; Prokoph, A. Temperatures and oxygen isotopic composition of Phanerozoic oceans. Earth Sci. Rev. 2015, 146, 92–104. [Google Scholar] [CrossRef]
- Benito, M.I.; Reolid, M.; Viedma, C. On the microstructure, growth pattern and original porosity of belemnite rostra: Insights from calcitic Jurassic belemnites. J. Iber. Geol. 2016, 42, 201–226. [Google Scholar]
- Hoffmann, R.; Richter, D.K.; Neuser, R.D.; Jöns, N.; Linzmeier, B.J.; Lemanis, R.E.; Fusseis, F.; Xiao, X.; Immenhauser, A. Evidence for a composite organic–inorganic fabric of belemnite rostra: Implications for palaeoceanography and palaeoecology. Sed. Geol. 2016, 341, 203–215. [Google Scholar] [CrossRef]
- Benito, M.I.; Reolid, M. Comparison of the Calcareous Shells of Belemnitida and Sepiida: Is the Cuttlebone Prong an Analogue of the Belemnite Rostrum Solidum? Minerals 2020, 10, 713. [Google Scholar] [CrossRef]
- Richter, D.K.; Neuser, R.D.; Schreuer, J.; Gies, H.; Immenhauser, A. Radial-fibrous calcites: A new look at an old problem. Sed. Geol. 2011, 239, 23–36. [Google Scholar] [CrossRef]
- Anagnōstu, C. Sedimentpetrographische Untersuchungen im Mittleren und Oberen Dogger Süddeutschlands. Bochumer Geol. Geotech. Arbeiten 1987, 25, 1–291. [Google Scholar]
- Linzmeier, B.J.; Kozdon, R.; Peters, S.E.; Valley, J.W. Oxygen isotope variability within Nautilus shell growth bands. PLoS ONE 2016, 11, e0153890. [Google Scholar]
- Śliwiński, M.G.; Kitajima, K.; Kozdon, R.; Spicuzza, M.J.; Fournelle, J.H.; Denny, A.; Valley, J.W. Secondary Ion Mass Spectrometry Bias on Isotope Ratios in Dolomite–Ankerite, Part I: δ18O Matrix Effects. Geostand. Geoanalytical Res. 2016, 40, 157–172. [Google Scholar] [CrossRef]
- Kozdon, R.; Ushikubo, T.; Kita, N.T.; Spicuzza, M.; Valley, J.W. Intratest oxygen isotope variability in the planktonic foraminifer N. pachyderma: Real vs. apparent vital effects by ion microprobe. Chem. Geol. 2009, 258, 327–337. [Google Scholar] [CrossRef]
- Pacton, M.; Breitenbach, S.F.M.; Lechleitner, F.A.; Vaks, A.; Rollion-Bard, C.; Gutareva, O.S.; Osintcev, A.V.; Vasconcelos, C. The role of microorganisms in the formation of a stalactite in Botovskaya Cave, Siberia paleoenvironmental implications. Biogeosciences 2013, 10, 6115. [Google Scholar] [CrossRef]
- Segar, D.A.; Collins, J.D.; Riley, J.P. The Distribution of the Major and Some Minor Elements in Marine Animals Part II. Molluscs. J. Mar. Biolog. Assoc. U. K. 1971, 51, 131–136. [Google Scholar] [CrossRef]
- Longinelli, A.; Wierzbowski, H.; Di Matteo, A. δ18O(PO43−) and δ18O(CO32−) from belemnite guards from Eastern Europe: Implications for palaeoceanographic reconstructions and for the preservation of pristine isotopic values. Earth Planet Sci. Lett. 2003, 209, 337–350. [Google Scholar] [CrossRef]
- Cuif, J.-P.; Dauphin, Y.; Berthet, P.; Jegoudez, J. Associated water and organic compounds in coral skeletons: Quantitative thermogravimetry coupled to infrared absorption spectrometry. Geochem. Geophys. Geosyst. 2004, 5, Q11011. [Google Scholar] [CrossRef]
- Orland, I.J.; Bar-Matthews, M.; Ayalon, A.; Matthews, A.; Kozdon, R.; Ushikubo, T.; Valley, J.W. Seasonal resolution of Eastern Mediterranean climate change since 34 ka from a Soreq Cave speleothem. Geochim. Cosmochim. Acta 2012, 89, 240–255. [Google Scholar] [CrossRef]
- Helser, T.E.; Kastelle, C.R.; McKay, J.L.; Orland, I.J.; Kozdon, R.; Valley, J.W. Evaluation of micromilling/conventional isotope ratio mass spectrometry and secondary ion mass spectrometry of δ18O values in fish otoliths for sclerochronology. Rapid Commun. Mass Spectrom. 2018, 32, 1781–1790. [Google Scholar] [CrossRef]
- Coplen, T.B. Calibration of the calcite-water oxygen-isotope geothermometer at Devils Hole, Nevada, a natural laboratory. Geochim. Cosmochim. Acta 2007, 71, 3948–3957. [Google Scholar] [CrossRef]
- Daëron, M.; Drysdale, R.N.; Peral, M.; Huyghe, D.; Blamart, D.; Coplen, T.B.; Lartaud, F.; Zanchetta, G. Most Earth-surface calcites precipitate out of isotopic equilibrium. Nat. Comm. 2019, 10, 429. [Google Scholar] [CrossRef]
- Geisler, T.; Perdikouri, C.; Kasioptas, A.; Dietzel, M. Real-time monitoring of the overall exchange of oxygen isotopes between aqueous and H2O by Raman spectroscopy. Geochim. Cosmochim. Acta 2012, 90, 1–11. [Google Scholar] [CrossRef]
- Tripati, A.K.; Hill, P.S.; Eagle, R.A.; Mosenfelder, J.L.; Tang, J.; Schauble, E.A.; Eiler, J.M.; Zeebe, R.E.; Uchikawa, J.; Coplen, T.B.; et al. Beyond temperature: Clumped isotope signatures in dissolved inorganic carbon species and the influence of solution chemistry on carbonate mineral composition. Geochim. Cosmochim. Acta 2015, 166, 344–371. [Google Scholar] [CrossRef]
- Watkins, J.M.; Hunt, J.D. A process-based model for non-equilibrium clumped isotope effects in carbonates. Earth Planet Sci. Lett. 2015, 432, 152–165. [Google Scholar] [CrossRef]
- Watkins, J.M.; Nielsen, L.C.; Ryerson, F.J.; DePaolo, D.J. The influence of kinetics on the oxygen isotope composition of calcium carbonate. Earth Planet Sci. Lett. 2013, 375, 349–360. [Google Scholar] [CrossRef]
- Kim, S.-T.; O’Neil, J.R. Equilibrium and nonequilibrium oxygen isotope effects in synthetic carbonates. Geochim. Cosmochim. Acta 1997, 61, 3461–3475. [Google Scholar] [CrossRef]
- Watkins, J.M.; Hunt, J.D.; Ryerson, F.J.; DePaolo, D.J. The influence of temperature, pH, and growth rate on the δ18O composition of inorganically precipitated calcite. Earth Planet Sci. Lett. 2014, 404, 332–343. [Google Scholar] [CrossRef]
- Chen, S.; Gagnon, A.C.; Adkins, J.F. Carbonic anhydrase, coral calcification and a new model of stable isotope vital effects. Geochim. Cosmochim. Acta 2018, 236, 179–197. [Google Scholar] [CrossRef]
- Miyamoto, H.; Miyashita, T.; Okushima, M.; Nakano, S.; Morita, T.; Matsushiro, A. A carbonic anhydrase from the nacreous layer in oyster pearls. Proc. Natl. Acad. Sci. USA 1996, 93, 9657–9660. [Google Scholar] [CrossRef]
- McConnaughey, T.A. Sub-equilibrium oxygen-18 and carbon-13 levels in biological carbonates: Carbonate and kinetic models. Coral Reefs 2003, 22, 316–327. [Google Scholar] [CrossRef]
- Uchikawa, J.; Zeebe, R.E. The effect of carbonic anhydrase on the kinetics and equilibrium of the oxygen isotope exchange in the CO2–H2O system: Implications for δ18O vital effects in biogenic carbonates. Geochim. Cosmochim. Acta 2012, 95, 15–34. [Google Scholar] [CrossRef]
- Löffler, N.; Fiebig, J.; Mulch, A.; Tütken, T.; Schmidt, B.C.; Bajnai, D.; Conrad, A.C.; Wacker, U.; Böttcher, M.E. Refining the temperature dependence of the oxygen and clumped isotopic compositions of structurally bound carbonate in apatite. Geochim. Cosmochim. Acta 2019, 253, 19–38. [Google Scholar] [CrossRef]
- McConnaughey, T.A. 13C and 18O isotopic disequilibrium in biological carbonates: I. Patterns. Geochim. Cosmochim. Acta 1989, 53, 151–162. [Google Scholar] [CrossRef]
- Zeebe, R.E. An explanation of the effect of seawater carbonate concentration on foraminiferal oxygen isotopes. Geochim. Cosmochim. Acta 1999, 63, 2001–2007. [Google Scholar] [CrossRef]
- Thébault, J.; Chauvaud, L.; Clavier, J.; Guarini, J.; Dunbar, R.B.; Fichez, R.; Mucciarone, D.A.; Morize, E. Reconstruction of seasonal temperature variability in the tropical Pacific Ocean from the shell of the scallop, Comptopallium radula. Geochim. Cosmochim. Acta 2007, 71, 918–928. [Google Scholar] [CrossRef][Green Version]
- Kennett, J.P.; Shackleton, N.J. Laurentide Ice Sheet Meltwater Recorded in Gulf of Mexico Deep-Sea Cores. Science 1975, 188, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Checa, A.G. Physical and biological determinants of the fabrication of molluscan shell microstructures. Front. Mar. Sci. 2018, 5, 353. [Google Scholar] [CrossRef]
- Seilacher, A.; Chinzei, K. Remote biomineralization 2: Fill skeletons controlling buoyancy in shelled cephalopods. Neues Jb. Geol. Paläontol. Abh. 1993, 190, 363–373. [Google Scholar]
- Seknazi, E.; Kozachkevich, S.; Polishchuk, I.; Bianco-Stein, N.; Villanova, J.; Suuronen, J.-P.; Dejoie, C.; Zaslansky, P.; Katsman, A.; Pokroy, B. From spinodal decomposition to alternating layered structure within single crystals of biogenic magnesium calcite. Nat. Commun. 2019, 10, 4559. [Google Scholar] [CrossRef]
- Bianco-Stein, N.; Polishchuk, I.; Lang, A.; Atiya, G.; Villanova, J.; Zaslansky, P.; Katsman, A.; Pokroy, B. Structural and chemical variations in Mg-calcite skeletal segments of coralline red algae lead to improved crack resistance. Acta Biomater. 2021, 130, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Gutowska, M.A.; Melzner, F.; Pörtner, H.O.; Meier, S. Cuttlebone calcification increases during exposure to elevated seawater pCO2 in the cephalopod Sepia officinalis. Mar. Biol. 2010, 157, 1653–1663. [Google Scholar] [CrossRef]
- Crick, R.E.; Ottensman, V.M. Sr, Mg, Ca and Mn chemistry of skeletal components of a Pennsylvanian and recent nautiloid. Chem. Geol. 1983, 39, 147–163. [Google Scholar] [CrossRef]
- Seuss, B.; Mapes, R.H.; Klug, C.; Nützel, A. Exceptional cameral deposits in a sublethally injured Carboniferous orthoconic nautiloid from the Buckhorn Asphalt Lagerstätte in Oklahoma, USA. Acta Palaeontol. Pol. 2011, 57, 375–390. [Google Scholar] [CrossRef]
- Checa, A.G.; Macías-Sánchez, E.; Harper, E.M.; Cartwright, J.H.E. Organic membranes determine the pattern of the columnar prismatic layer of mollusc shells. Proc. R. Soc. B 2016, 283, 20160032. [Google Scholar] [CrossRef] [PubMed]
- Bajnai, D.; Guo, W.; Spötl, C.; Coplen, T.B.; Methner, K.; Löffler, N.; Krsnik, E.; Gischler, E.; Hansen, M.; Henkel, D.; et al. Dual clumped isotope thermometry resolves kinetic biases in carbonate formation temperatures. Nat. Comm. 2020, 11, 4005. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.J.; Davis, S.; John, C.M. Evidence of taxonomic non-equilibrium effects in the clumped isotope composition of modern cephalopod carbonate. Chem. Geol. 2021, 578, 120317. [Google Scholar] [CrossRef]
- Hoffmann, R.; Stevens, K. The palaeobiology of belemnites—Foundation for the interpretation of rostrum geochemistry. Biol. Rev. 2020, 95, 94–123. [Google Scholar] [CrossRef] [PubMed]
- Jereb, P.; Roper, C.F.E. (Eds.) Cephalopods of the World: An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date Volume 1—Chambered Nautiluses and Sepioids (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae and Spirulidae); FAO Species Catalogue for Fishery Purposes; Food and Agriculture Organization of the United Nations—FAO: Rome, Italy, 2020; Volume 4, 262p. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, R.; Linzmeier, B.J.; Kitajima, K.; Nehrke, G.; Dietzel, M.; Jöns, N.; Stevens, K.; Immenhauser, A. Complex Biomineralization Pathways of the Belemnite Rostrum Cause Biased Paleotemperature Estimates. Minerals 2021, 11, 1406. https://doi.org/10.3390/min11121406
Hoffmann R, Linzmeier BJ, Kitajima K, Nehrke G, Dietzel M, Jöns N, Stevens K, Immenhauser A. Complex Biomineralization Pathways of the Belemnite Rostrum Cause Biased Paleotemperature Estimates. Minerals. 2021; 11(12):1406. https://doi.org/10.3390/min11121406
Chicago/Turabian StyleHoffmann, René, Benjamin J. Linzmeier, Kouki Kitajima, Gernot Nehrke, Martin Dietzel, Niels Jöns, Kevin Stevens, and Adrian Immenhauser. 2021. "Complex Biomineralization Pathways of the Belemnite Rostrum Cause Biased Paleotemperature Estimates" Minerals 11, no. 12: 1406. https://doi.org/10.3390/min11121406
APA StyleHoffmann, R., Linzmeier, B. J., Kitajima, K., Nehrke, G., Dietzel, M., Jöns, N., Stevens, K., & Immenhauser, A. (2021). Complex Biomineralization Pathways of the Belemnite Rostrum Cause Biased Paleotemperature Estimates. Minerals, 11(12), 1406. https://doi.org/10.3390/min11121406