3.1. Systematic Description
Uncinifera Engeser, 1990 (parataxon (form taxon) of ordinal level)
form-genus Onychites Quenstedt, 1857
Type species: Onychites ornatus Quenstedt, 1857, SD Quenstedt, 1866.
form-species O. aff. barbatus (Fraas, 1855)
pars 1855 Onychoteuthis barbata; Fraas 1855, 89
pars 1857 Onychites barbatus; Quenstedt 1857, 804, pl. 99 Figure 14 (non Figure 15)
1987 Onychites barbatus Quenstedt 1857; Engeser 1987, 12, pl. 2 Figure 3
pars 1999 Onychites barbatus (Fraas 1855); Schweigert 1999, 11, pl. 5 Figure 1, 3; pl. 6 Figure 5
2010 Onychites barbatus; Stevens 2010, 400, Figure 4h
2013 Onychites barbatus (Fraas 1855); Hammer et al., 2013, 833
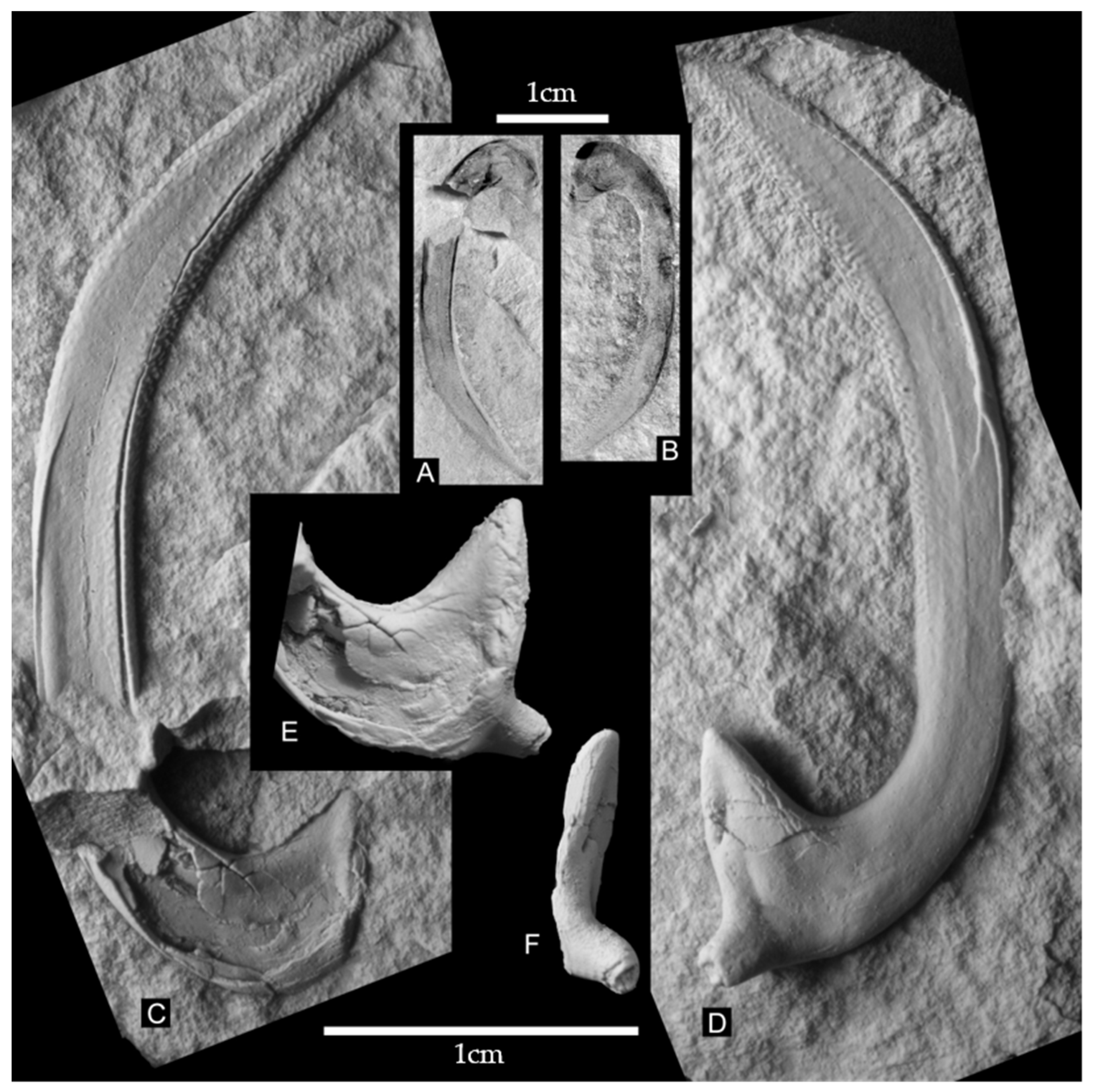
The specimen is 31 mm long and 10 mm high (i.e., largest total dimension perpendicular to length).
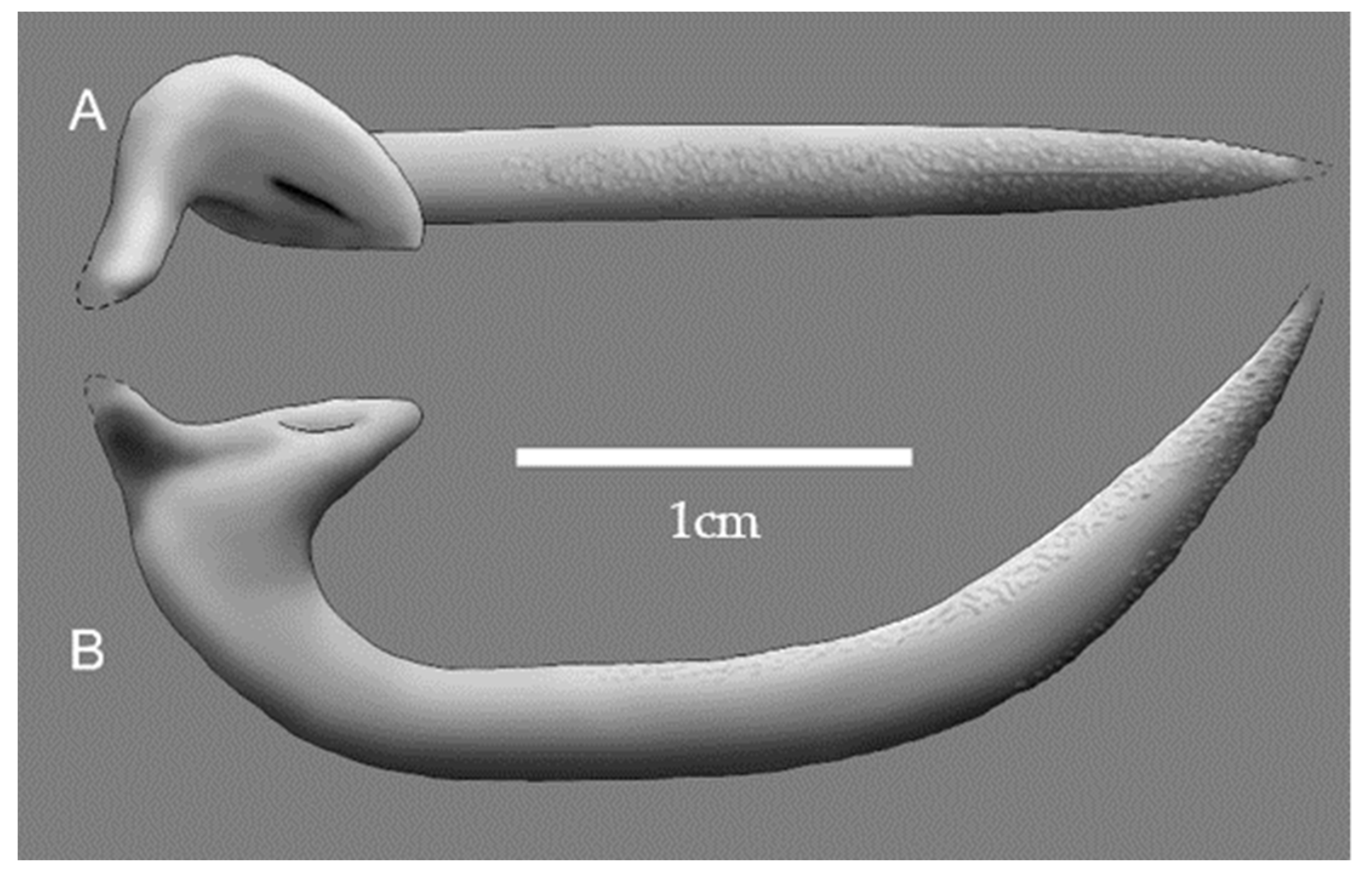
The base is 8.5 mm long, measured parallel to total length. The inner (anterior) process is ~3 mm long and has a subtriangular outline. The outer (posterior) process tapers slightly and is broken at the tip. The posterior process projects slightly (>1 mm) toward the posterior and conspicuously (1 mm) downward (i.e., toward the axis of the animal body), which gives the base a concave profile. The base is positioned at a considerable angle relative to the curvature plane of the shaft and uncinus (in vivo―ventrally or dorsally; in rock―probably downward), protruding ~1.3 mm toward this latter direction. Considering sediment compaction, the preserved part of the process originally protruded ~2–4 mm from the base, which is also strongly flattened. Thus, together with the process, the breadth of the base could have exceeded its length. The inner process also pointed in the same direction, albeit to a lesser extent. The position of the processes serves to differentiate onychite sides, i.e., the processes-bearing side and the side lacking processes (one of these was ventral, the other dorsal). Reconstructions of the onychite appearance before crushing and compaction are shown in
Figure 4.
On the processes-bearing side, at the base of the inner process, there is an inward-directed (i.e., toward the body axis) slit that most likely is a collapsed opening or a depression. Despite relatively good preservation, the orbicular scar could not be located. On the opposite side, however, despite considerable crushing, there seems to be a fringed ridge along its edge on the inner process. This ridge could possibly represent the orbicular scar.
Along its entire length, the shaft is 3.5 mm wide (originally, before flattening, it was probably ~3 mm wide). The original cross-section was probably oval, slightly flattened laterally. The shaft is strongly bent near the base. The curvature is gradually reduced, and the shaft continues as a 5 mm straight section, approximately parallel to the basal line. Following this straight section, there is a uniform arch that passes into the uncinus. Over the terminal ~10 mm, the uncinus tapers. The specimen has a blunt termination, but the tip (1–2 mm the length) was probably broken off. The tip protruded beyond the basal line.
The ornamentation is the most pronounced at the tip. Ornaments are in the form of roundish knobs, usually linked in groups of few, in the form of irregular wrinkles. On the processes-bearing side, some 3 mm from the preserved tip of a knob, in the middle section of the side, the knobs become separated, less pronounced, slightly elongated, and finally disappear altogether some 8 mm from the tip. The remainder of the side is smooth. On the other side, the ornamentation disappears closer to the tip, and the smooth belt is broader. On the external side, the ornament disappears at the end of the uncinus arch. On the internal side, the ornament reaches almost to the end of the straight section of the shaft. At the uncinus deflection, the dominant direction of the wrinkles is skewed, approximately perpendicular to the longest dimension. Along the internal edge of the uncinus there appears to be a kind of a smooth edge, visible on both imprints. It is impossible to say whether this was indeed a smooth edge; it could possibly be a fracture within the plane of maximum width, along which both sides aligned.
The entire onychite was hollow inside, and the walls were ~0.1–0.15 mm thick.
3.2. Taxonomic Discussion
The specimens of
O. barbatus illustrated in the literature, including the specimen of Fraas [
1] that was chosen as a paratype by Schweigert [
11] (pl. 5, Figure 1,3), the next two specimens illustrated by the latter author (pl. 6, Figure 5), and the specimens of Quenstedt [
2] (pl. 99, Figure 14) and Engeser [
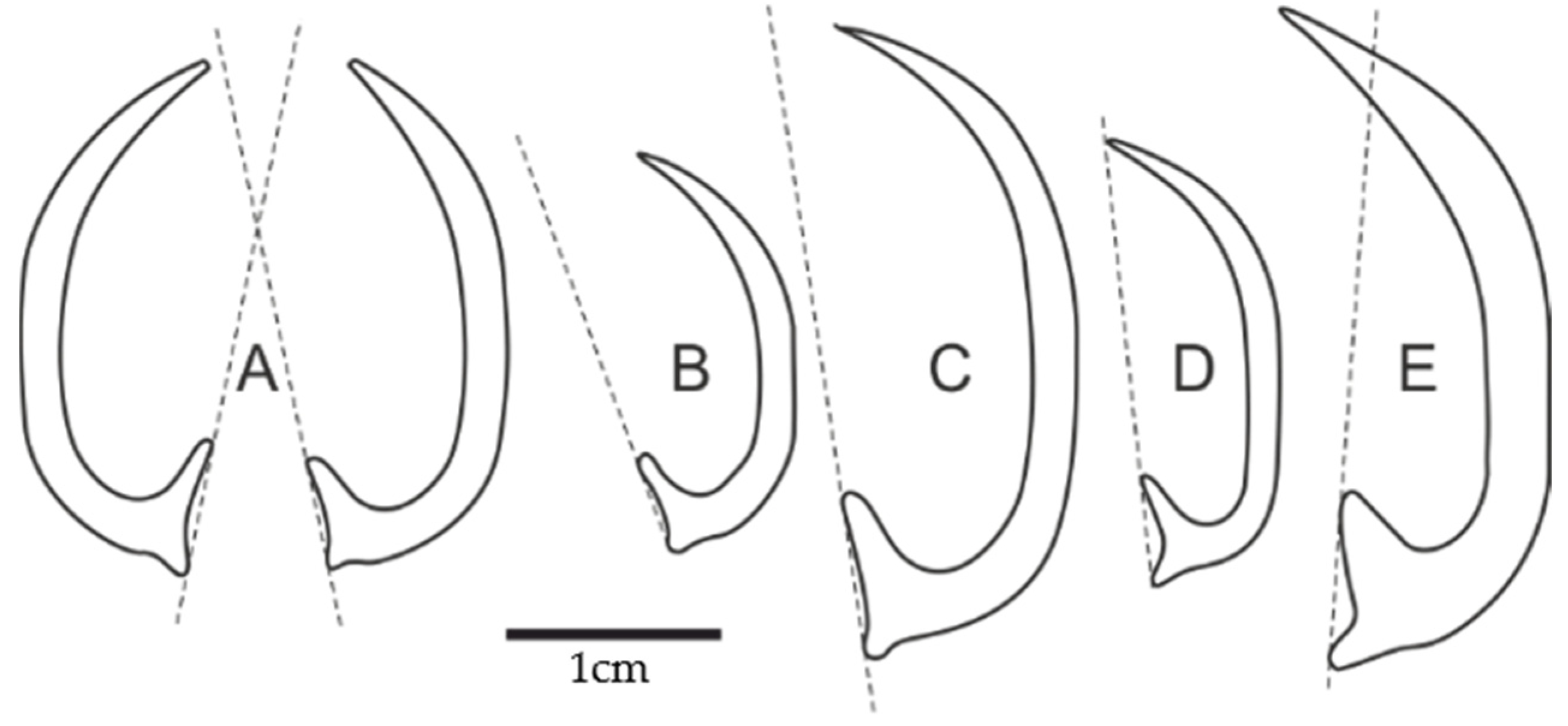
3] (pl. 2, Figure 3), do not display significant differences among themselves. They are, respectively, 24, 18, 29, and 21 mm long and have basically the same shape and proportions (see comparison in
Figure 5). Note, however, that Engeser [
3] and Stevens [
5] reported different lengths for the Quenstedt specimen. The specimen presented here has almost precisely the same outline and ornamentation as the above specimens, but it is slightly larger (31 mm) and considerably broader (0.11 of total length, against 0.08 for the remaining specimens). My specimen also has a shorter (0.1 vs. 0.11–0.17) and broader inner process. The larger breadth could have resulted from crushing, but the well-illustrated specimen of Engeser shows identical fracturing and, thus, likely a similar degree of broadening. Furthermore, the illustrated specimens of previous workers are less curved; their tips do not project beyond the basal line. The ornament visible in Engeser’s illustration terminates more proximally on the shaft, but the smooth surface in the middle of the side is shorter. The previous authors also did not mention the lateral position of the outer process, although it should be visible on the specimens preserved to some degree three-dimensionally, since the ornament is visible. Even though the specimen presented here falls outside the range of variation of the material from Germany, the differences may not be sufficient to establish a separate form, especially given the variation known from a larger number of specimens such as
O. macnaughti Marwick, 1956 [
5] and
O. quenstedti Engeser, 1987 [
4].
To date, the only specimens of rostra preserved together with onychites belong to
Passaloteuthis paxillosa (Schlotheim, 1820). Recognized in association with these are
O. amalthei Quenstedt, 1856 and
O. uncus Quenstedt, 1856 [
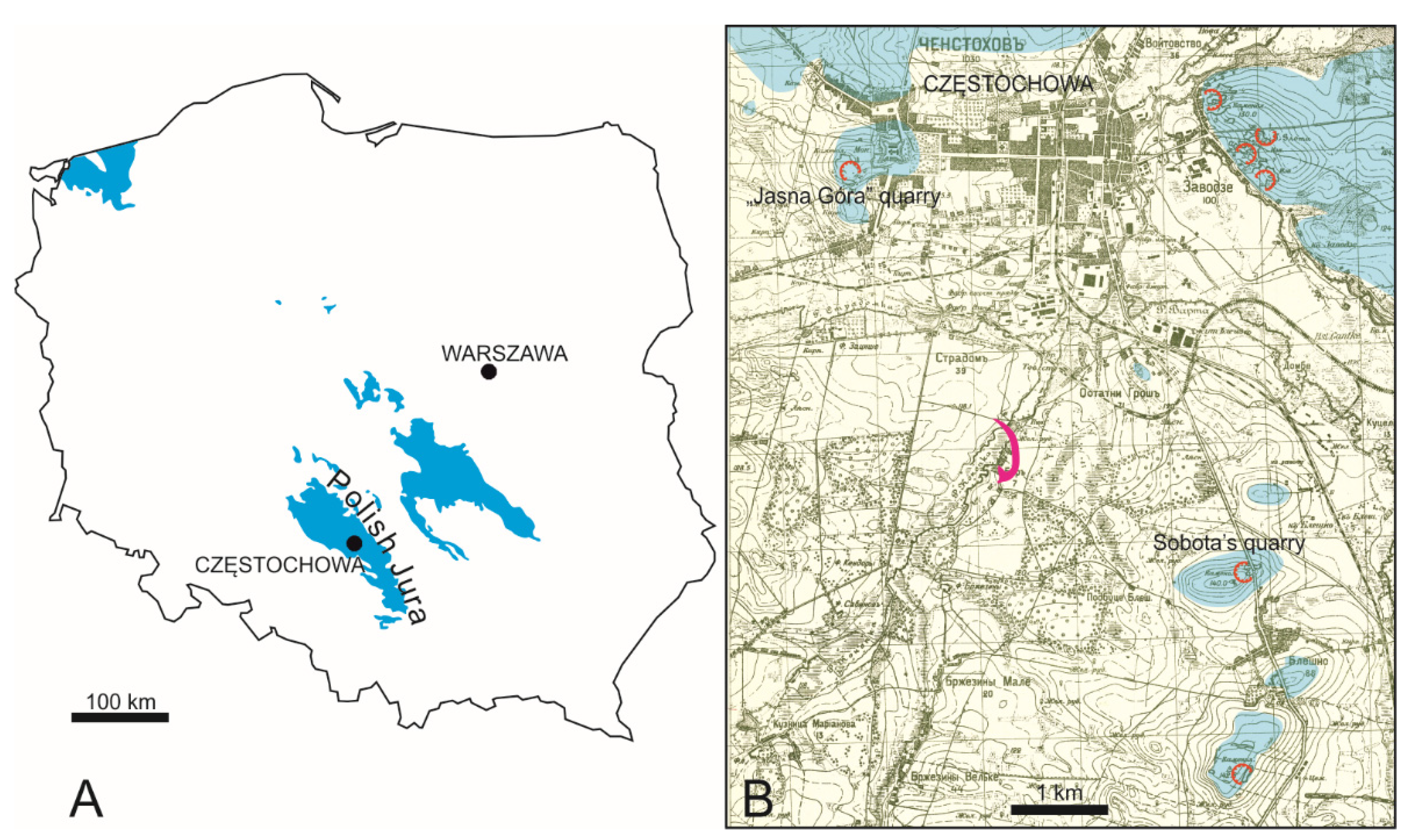
3]. Thus, it can only be stated with certainty that representatives of the family Belemnitidae had onychites. In the Oxfordian of the Polish Jura, however, they are extremely rare. I am familiar with two specimens only, from the lower Oxfordian of Wrzosowa (apical part only, MGUS/Sz.1943) and from Zalas (not numbered). In any case, the rare occurrence of both Belemnitidae and onychites (besides, in not exactly age-equivalent strata) is no evidence for their association.
The type material of
O. barbatus comes from Nusplingen in Germany. Apart from numerous onychites (including
O. fraasi Quenstedt, 1857 and
O. quenstedti), the site yielded representatives of only one belemnite species,
Hibolites semisulcatus (Münster, 1830) [
11], which makes a strong case for the association between these onychite forms with that particular belemnite species. Moreover, a rich assemblage of
O. macnaughti from New Zealand co-occurs with representatives of the Belemnopseidae [
5]. The only belemnite species that I found in the material from which the onychite reported here originates and in age-equivalent platy limestones of the Polish Jura are
Hibolites hastatus Montfort, 1808 and
H. girardoti (Loriol, 1902). It is, therefore, likely that the onychite comes from one of these species.
The morphological variation among onychites associated with
Passaloteuthis paxillosa and
H. semisulcatus may be interpreted as evidence for their higher specificity [
3] or intraspecific variability [
11]. The first option seems considerably more likely. Contrary to rostra (which are part of the internal skeleton and invisible to females), as a manifestation of sexual dimorphism, onychites were subject to sexual selection, which is known to be a strong driver of speciation and species diversification. It seems to be different for
O. macnaughti, which co-occurs with several species of the Belemnopseidae.
O. macnaughti, however, is so variable (as evidenced by illustrations in [
5]) that, with a larger number of complete specimens, it would likely be possible to distinguish further species. It also seems unlikely that onychites have been found of all species of belemnites, identified on the base of rostra, which are many times more numerous. Thus, I suspect that belemnite species described using rostra include numerous cryptospecies. Unfortunately, this cannot be rectified on the basis of onychites, as they are too rare and almost invariably disarticulated from rostra.
3.3. Remarks on Functional Morphology
As only a minor fraction of the few complete belemnite finds include onychites, most authors since Engeser [
3] have assumed they played a part in mating. On the basis of the shape of a particular paraspecies, Hammer et al. [
4] performed a functional analysis, suggesting that the studied form followed a logarithmic spiral, typical of vertebrate claws. Hammer et al. [
4] found that the spiral center in
O. quenstedti occurred near the base, unlike in vertebrates, where it occurs adjacent to the top. On the basis of this difference, Hammer et al. [
4] concluded that the function of the onychite must have been associated with pushing rather than pulling. Lastly, Hammer et al. [
4] cited the opinions of other workers, who suggested that onychites served for holding the female during copulation [
3], and that onychites may have served for fights among males [
12,
13], regarding the function of special arm hooks in present-day coleoid males.
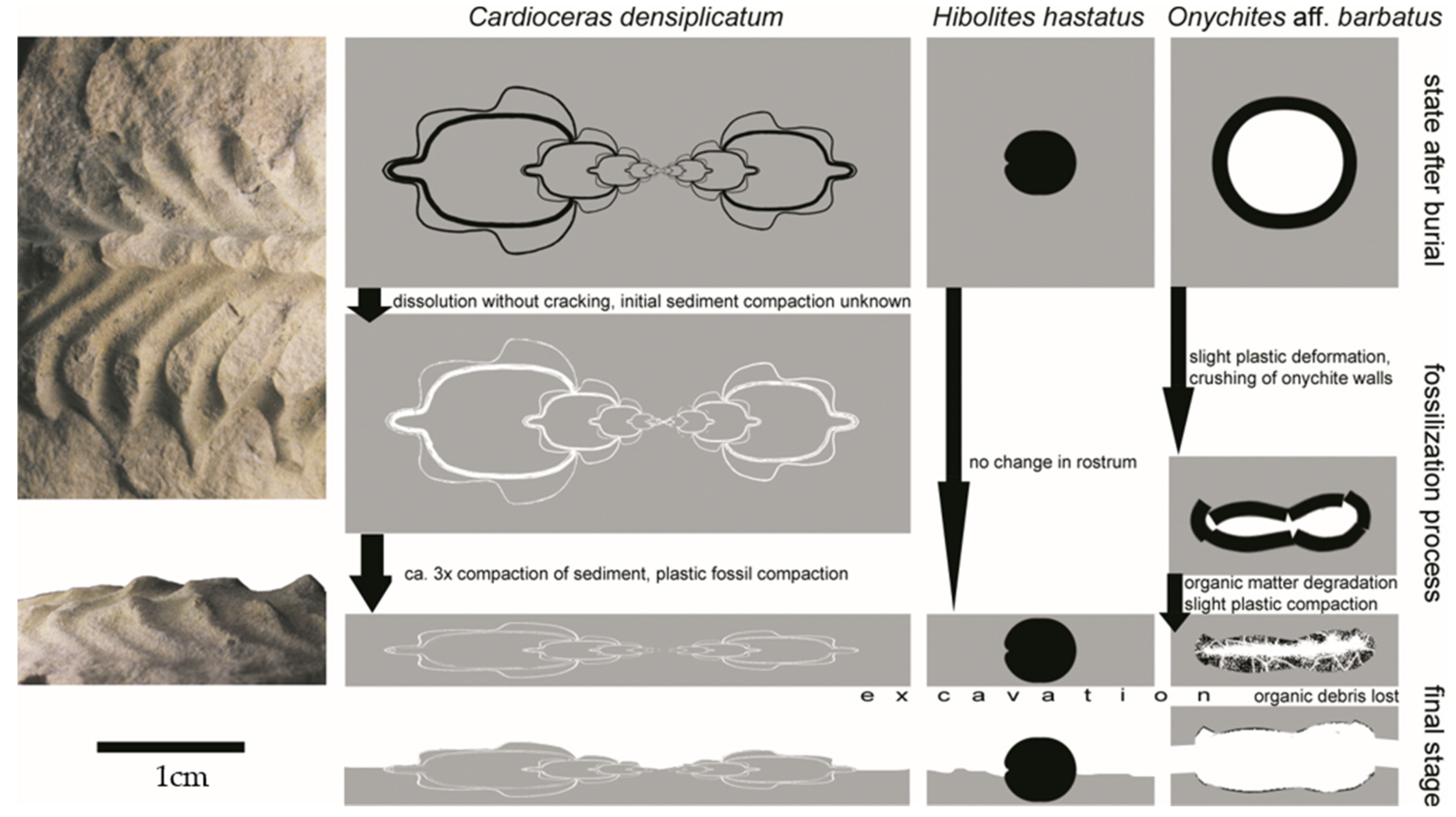
The logarithmic spiral is a remarkable shape. Vertebrate claws (and mollusk shells) attain this shape naturally, owing to constant growth of the claw at one end, in a simultaneously growing individual. In this way, the claws have the same properties along the whole section, on a gradually larger scale. Mechanical properties of the logarithmic spiral are a benefit to vertebrates, but they are an added value to the laws of growth. Chitinous hooks of the cephalopods are not used before fully formed, and their logarithmic spiral shape (or whatever other shape) must have been attained by natural selection or accidentally. Not all paraspecies, including the studied one, have such shape. Thus, the logarithmic spiral shape was not always optimum, shape imperfections (giving a constant and uniform strain distribution during specific work) were equally well compensated for by structural variation (structure, wall thickness, cross-section shape), or the properties of the logarithmic spiral were of no significant importance.
All onychites are more or less bent into an arch and tapered at their tips, although some (
O. amalthei, O. ornatus, and
O. contractus, based on illustrations in Engeser [
3]) have broad tips. These three relatively blunt forms would be unsuitable for stabbing. The remaining onychites must have served this function, as the necessity of holding a female by embracing her does not justify the presence of sharp tips that could have hurt her, unless holding the female required anchoring the male by driving the hooks into the female body. Efficient holding, regardless of the exact mode, would have required an appropriate hook curvature.
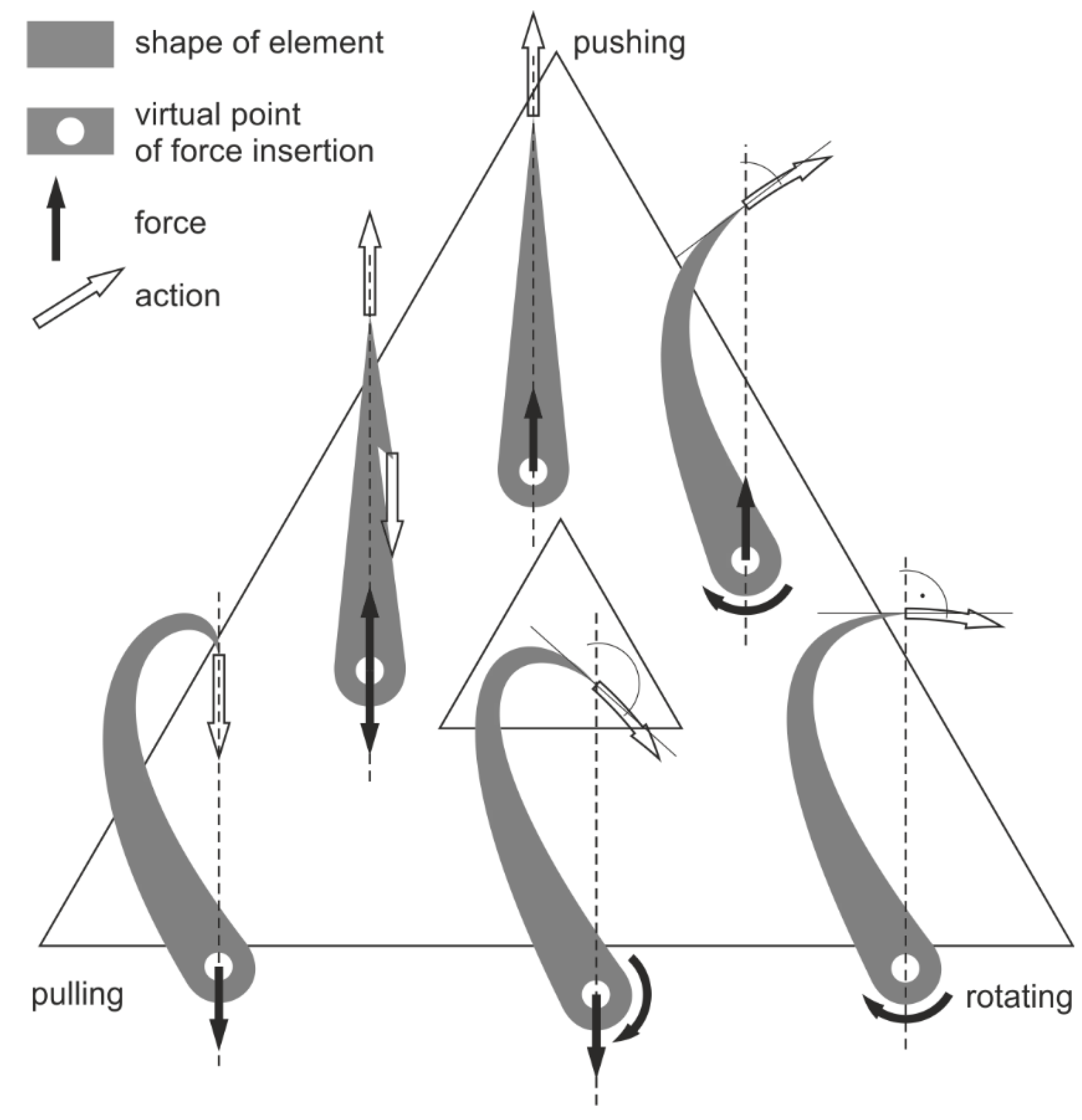
The shapes of sharp-tipped anatomical elements (serving for stabbing or piercing) depend on the work mechanics. In the simplest cases, in pushed elements, the tip is directed against the attachment point (as is typical of spines). In pulled elements such as hooks, the tip is directed toward the attachment point, and, in elements that rotate around an axis extending across the line linking the tip and the attachment (such as claws), the tip is directed perpendicularly relative to that axis, within the plane of rotation. The actual morphological diversity of sharp elements is due to more complex function, using two different forces. Thus, teeth of carnivores are often bent toward the rear. This serves to pierce by closing the jaw, which moves along an arch, and to both prevent the prey from setting itself free and facilitate moving the prey toward the esophagus (i.e., pushing and rotating). Harpoons have a spine directed toward the front and a second one (or more) directed toward the rear, in order to prevent the escape of the pierced prey (pushing and pulling). Claws are often bent further than perpendicular relative to the anchor point in order to effortlessly hold the captured prey (rotating and pulling). I cannot conceive any naturally occurring sharp object that would use a combination of all three of these forces or use the fourth possible force, i.e., revolving around an axis extending between the tip and the attachment point, such as a corkscrew or a screw. The work mechanism of these elements is schematically represented in
Figure 6.
Given all of the above, the function of arm hooklets is also easily explained. Individuals with diverse hooklets preserved make especially interesting cases. In the diplobelid
Chondroteuthis wunnenbergi illustrated by Hoffmann et al. [
14], hooklets characteristically differ according to their location along an arm ([
14] Figure 9). The closer to the tips of the arms, the stronger the curvature of the hooklets, which suggests that they served to capture and transfer the prey toward the less curved, proximal hooklets, onto which the prey was threaded. The pronounced difference in the size of some hooklets may be explained by their use against considerably larger objects; belemnite prey was, on average, circa an order of magnitude smaller than belemnites themselves [
15,
16]. Fighting males, however, were similar in size. The cited workers [
14] reached similar conclusions on the basis of more complex analyses.
In the studied specimen (as with most other onychite paraspecies), the tip is directed away from the base, inclined ~45° relative to the basal line. Morphologically, therefore, it falls precisely between a spine and a hook from
Figure 6, which suggests it served for stabbing with rotation. Teeth of carnivorous vertebrates, arthropod chelicerae, or scorpion stings, which operate in precisely the same way, sometimes have similar shapes. Notably, in both arthropods and vertebrates, these elements are supported by the skeleton, which―being resistant to compression―enables pushing them toward the prey. Theoretically, in their heads and arms, belemnites could only use their hydraulic skeleton (like a wasp extending its sting), but the structure of the onychite base and the curvature of other onychites argue against this. Onychites were probably supported by an elastic cartilage and were moved by muscles that could direct them in the appropriate direction, but could not push them away from the body.
An element rotated by a muscle needs support from elsewhere than muscle attachment. The larger the distance between the point of support and the muscle attachment, the longer the lever, and the higher torque required from the muscles. No apparent muscle attachment is seen on the studied element. They were definitely absent beyond the base; circular structures on the shaft of the specimen illustrated by Hammer et al. [
4] are, in my opinion, silicification nodes or similar structures described by Klug et al. [
17] formed during diagenesis rather than muscle impressions. Both the shaft and the uncinus lack structures resembling an orbicular scar or longitudinal ridges, which indicates that the element was attached to the body only through the base, and it is the base diameter that determines the maximum lever length. Considering the cartilaginous (i.e., elastic) substratum and the diminutive diameter of the base relative to the total length of the onychite, torque levels must have been low. It is highly unlikely that a belemnite could efficiently hold anything with such structures, including a female, especially given that the onychite tips were directed away from the base. It is also entirely impossible that the onychites were positioned at the ends or in the middle of the arms’ length, as reconstructed by Stevens [
5]. The arms lacked a skeleton and, therefore, could not offer sufficient support for a lever the length of an onychite in order to operate it in any way.
Complete belemnite specimens [
16,
18] reveal that onychites were anchored at the head or at the base of a pair of arms, with tips directed toward the body axis. The stabbed object must have been located in front of the animal, and the purpose of the action must have been to bring the prey to the tips of the onychite by means of the arms or to direct these tips toward the aggressor, which charged on its own and drew the competitor toward itself. In the latter case, it could have been a form of defense against getting bitten on the head by the beak of the opponent.
The slight curvature of the hooks did not facilitate holding the stabbed object and, further, the dominant direction of the wrinkles likely did not favor a deep stab. Assuming that onychites served for male fights, they were likely a defensive weapon, used for rapid repulsion of an attack and deterrence. The curvature helped to dispose of the stabbed element by abduction of the onychites (following the mechanism described by Hammer et al. [
4] for the logarithmic spiral). This would have been impossible with hypothetical straight onychites (which have never been found).
Possible onychite mobility could be hypothesized on using the shapes of the base and muscle attachments. Unfortunately, no muscle attachments are seen, and the shape reconstruction is only tentative because the base is crushed (
Figure 2). An interesting feature of the base, typical also for fossil arm hooks, is that the base was directed toward the body axis. Arm hooks, however, bear orbicular scars and longitudinal ridges, indicating that they were anchored or supported also on the internal side. Nothing similar is to be seen on the specimen examined here. This would suggest that the studied specimen was attached to the head rather than an arm. If it was attached to an arm, however, it would have been placed at the base of an arm and more likely offset to a side rather than attached on the internal side. The strong base asymmetry suggests a functional difference between the two onychite sides (i.e., processes-bearing side and the one lacking processes). Perhaps these hooks were not facing one another but placed at an angle on one side of the animal (ventral or dorsal), and they worked in partly intersecting planes. This could explain their parallel position in the specimen illustrated by Dick et al. [
18].
A hypothetical muscle anchored in the onychite’s interior could have passed through the opening in the base. If the posterior part of the base with the posterior process was the point of support, such a muscle could have served for rotating the onychite toward the body axis. Furthermore, it acted to increase the force of onychite anchoring in the body, and, if the muscle was set sufficiently deep in the onychite, it could have reduced bending moments within the onychite walls during some actions (similar to musculus intramandibularis in archosaurian mandibles), reducing the skeletal material required to form an onychite.
3.4. Geographic Variation of Published Onychite Morphologies and Functions
The above considerations cannot be accurate with regard to some other onychite paraspecies. Some had a relatively longer base (O. fraasi), which may have enabled higher torque. Some appear to have lacked a well-delineated base altogether (O. uncus, although the distinctly crushed distal part may suggest that it gradually transitioned into cartilage). O. uncus and (to a lesser extent) O. ornatus were more curved, and their tips were perpendicular to a line connecting them to the base. Thus, they were not suitable for stabbing toward the front, but instead could embrace and hold an object with little effort. Moreover, O. ornatus had broad and likely blunt tips which made it useless for stabbing. At present, little can be said on the functional mechanics of weakly curved, thick, and probably blunt O. amalthei and O. contractus. Their only purpose may have been for showing off.
The tuberculated surface not only made using the onychite for stabbing more difficult, but it could also have served as a sexual ornament, both visual and sensory; today’s cephalopods are sensitive to the texture of the surfaces they touch.
In this context, it is notable that onychites display a geographic variation. Specimens from New Zealand (included in one species,
O. macnaughti [
5], despite a high diversity) differ from the remaining ones, known from the NW Tethys region. The former are more strongly sculpted than nearly all Tethyan specimens and usually less strongly curved (the tip is distant from the basal line and sometimes even points away from the basal line). The differences in onychite forms may be explained by differences in arbitrary preferences of females rather than differences in environmental conditions.
It is also intriguing (as also noted by Stevens [
5]) that
O. macnaughti is the most similar to
O. contractus from the Pliensbachian of Germany.
O. macnaughti co-occurs with belemnites from the suborder Belemnopseina, which first occurs in the Bajocian, and
O. contractus has to be linked to the suborder Belemnitina, which means that the similarity between the two species is almost certainly due to convergence. It is impossible to determine, however, how this related to similarities in mating rituals.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





