Tracking Organomineralization Processes from Living Microbial Mats to Fossil Microbialites

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
4.1. Study of Living Microorganisms Providing Hints to Their Identification in the Fossil Record
4.2. Microtextures and Prevailing Mineralogy
4.3. Calcite Precipitation in Crusts, Induced versus Influenced Organomineralization
4.4. Noncarbonate Phases
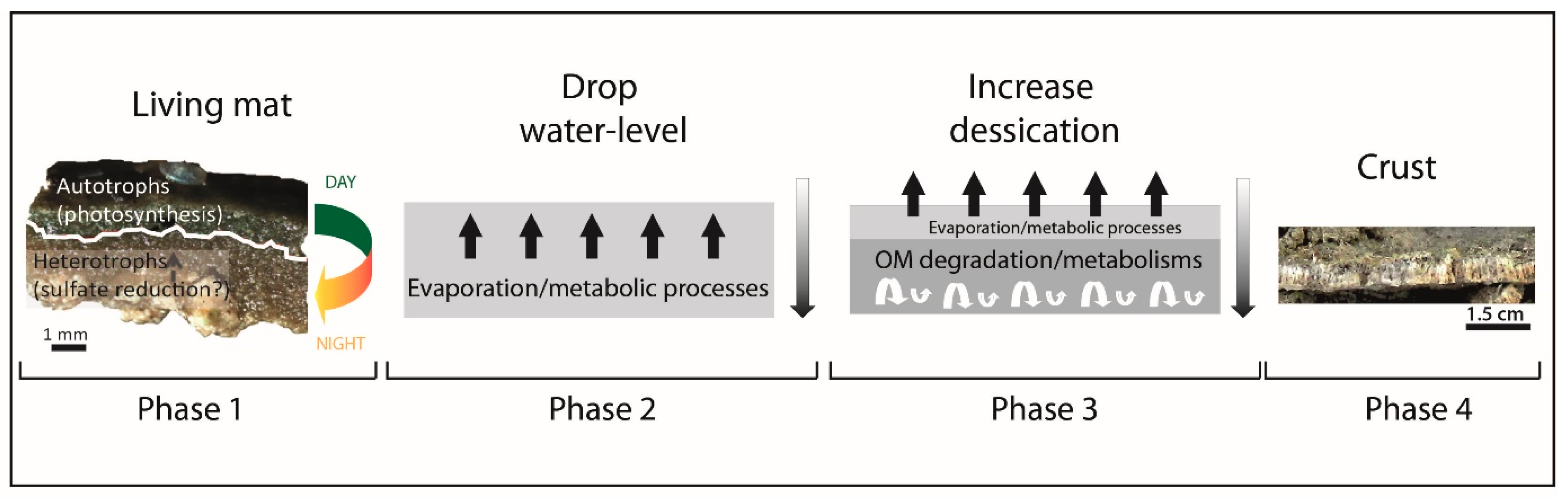
4.5. Introducing a Model of Formation for CaCO3 Crusts in the Maquinchao Basin
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Gemerden, H. Microbial mats: A joint venture. Mar. Geol. 1993, 113, 3–25. [Google Scholar] [CrossRef]
- Jorgensen, B.B.; Revsbech, N.P.; Cohen, Y. Photosynthesis and structure of benthic microbial mats: Microelectrode and SEM studies of four cyanobacterial communities1. Limnol. Oceanogr. 1983, 28, 1075–1093. [Google Scholar] [CrossRef]
- Stolz, J.F. Structure of Microbial Mats and Biofilms. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–8. ISBN 978-3-662-04036-2. [Google Scholar]
- Ginsburg, R.N. Controversies about stromatolites: Vices and virtues. In Controversies in Modern Geology; Müller, D.W., McKENZIE, J.A., Weissert, H.J., Eds.; Academic Press: London, UK, 1991; pp. 25–36. [Google Scholar]
- Riding, R. Microbialites, Stromatolites, and Thrombolites. In Encyclopedia of Geobiology; Reitner, J., Thiel, V., Eds.; Encyclopedia of Earth Sciences Series; Springer: Dordrecht, The Netherlands, 2011; pp. 635–654. [Google Scholar]
- Burne, R.V.; Moore, L.S. Microbialites: Organosedimentary Deposits of Benthic Microbial Communities. PALAIOS 1987, 2, 241. [Google Scholar] [CrossRef]
- Allwood, A.C.; Walter, M.R.; Kamber, B.S.; Marshall, C.P.; Burch, I.W. Stromatolite reef from the Early Archaean era of Australia. Nature 2006, 441, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Awramik, S.M.; Buchheim, H.P. A giant, Late Archean lake system: The Meentheena Member (Tumbiana Formation; Fortescue Group), Western Australia. Precambrian Res. 2009, 174, 215–240. [Google Scholar] [CrossRef]
- Lepot, K.; Benzerara, K.; Brown, G.E., Jr.; Philippot, P. Microbially influenced formation of 2724-million-year-old stromatolites. Nat. Geosci. 2008, 1, 118. [Google Scholar] [CrossRef]
- Andres, M.S.; Sumner, D.Y.; Reid, R.P.; Swart, P.K. Isotopic fingerprints of microbial respiration in aragonite from Bahamian stromatolites. Geology 2006, 34, 973–976. [Google Scholar] [CrossRef]
- Jahnert, R.J.; Collins, L.B. Characteristics, distribution and morphogenesis of subtidal microbial systems in Shark Bay, Australia. Mar. Geol. 2012, 303–306, 115–136. [Google Scholar] [CrossRef]
- Jahnert, R.J.; Collins, L.B. Controls on microbial activity and tidal flat evolution in Shark Bay, Western Australia. Sedimentology 2013, 60, 1071–1099. [Google Scholar] [CrossRef]
- Reid, R.P.; Macintyre, I.G.; Browne, K.M.; Steneck, R.S.; Miller, T. Modern marine stromatolites in the Exuma Cays, Bahamas: Uncommonly common. Facies 1995, 33, 1–17. [Google Scholar] [CrossRef]
- Arp, G.; Thiel, V.; Reimer, A.; Michaelis, W.; Reitner, J. Biofilm exopolymers control microbialite formation at thermal springs discharging into the alkaline Pyramid Lake, Nevada, USA. Sediment. Geol. 1999, 126, 159–176. [Google Scholar] [CrossRef]
- Farías, M.E.; Rascovan, N.; Toneatti, D.M.; Albarracín, V.H.; Flores, M.R.; Poiré, D.G.; Collavino, M.M.; Aguilar, O.M.; Vazquez, M.P.; Polerecky, L. The Discovery of Stromatolites Developing at 3570 m above Sea Level in a High-Altitude Volcanic Lake Socompa, Argentinean Andes. PLoS ONE 2013, 8, e53497. [Google Scholar] [CrossRef] [PubMed]
- Petryshyn, V.A.; Juarez Rivera, M.; Agić, H.; Frantz, C.M.; Corsetti, F.A.; Tripati, A.E. Stromatolites in Walker Lake (Nevada, Great Basin, USA) record climate and lake level changes ~35,000 years ago. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 451, 140–151. [Google Scholar] [CrossRef]
- Roche, A.; Vennin, E.; Bundeleva, I.; Bouton, A.; Payandi-Rolland, D.; Amiotte-Suchet, P.; Gaucher, E.C.; Courvoisier, H.; Visscher, P.T. The Role of the Substrate on the Mineralization Potential of Microbial Mats in a Modern Freshwater River (Paris Basin, France). Minerals 2019, 9, 359. [Google Scholar] [CrossRef]
- Suosaari, E.P.; Reid, R.P.; Playford, P.E.; Foster, J.S.; Stolz, J.F.; Casaburi, G.; Hagan, P.D.; Chirayath, V.; Macintyre, I.G.; Planavsky, N.J.; et al. New multi-scale perspectives on the stromatolites of Shark Bay, Western Australia. Sci. Rep. 2016, 6, srep20557. [Google Scholar] [CrossRef]
- Vasconcelos, C.; Warthmann, R.; McKenzie, J.A.; Visscher, P.T.; Bittermann, A.G.; van Lith, Y. Lithifying microbial mats in Lagoa Vermelha, Brazil: Modern Precambrian relics? Sediment. Geol. 2006, 185, 175–183. [Google Scholar] [CrossRef]
- Arenas, C.; Jones, B. Temporal and environmental significance of microbial lamination: Insights from Recent fluvial stromatolites in the River Piedra, Spain. Sedimentology 2017, 64, 1597–1629. [Google Scholar] [CrossRef]
- Bouton, A.; Vennin, E.; Boulle, J.; Pace, A.; Bourillot, R.; Thomazo, C.; Brayard, A.; Désaubliaux, G.; Goslar, T.; Yokoyama, Y.; et al. Linking the distribution of microbial deposits from the Great Salt Lake (Utah, USA) to tectonic and climatic processes. Biogeosciences 2016, 13, 5511–5526. [Google Scholar] [CrossRef]
- Merz-Preiß, M. Calcification in Cyanobacteria. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 50–56. ISBN 978-3-662-04036-2. [Google Scholar]
- Solari, M.A.; Hervé, F.; Le Roux, J.P.; Airo, A.; Sial, A.N. Paleoclimatic significance of lacustrine microbialites: A stable isotope case study of two lakes at Torres del Paine, southern Chile. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 70–82. [Google Scholar] [CrossRef]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth Sci. Rev. 2009, 96, 141–162. [Google Scholar] [CrossRef]
- Trichet, J.; Défarge, C. Non-biologically supported organomineralization. Bulletin de l’Institut Océanographique (Monaco) 1995, 14, 203–236. [Google Scholar]
- Reitner, J.; Gautret, P.; Martin, F.; Neuweiler, F. Automicrites in modern marine microbialite. Formation model via organic matrices (Lizard Island, Great Barrier Reef, Australia). Bulletin de l’Institut Océanographique (Monaco) 1995, numéro special 14, 237–264. [Google Scholar]
- Altermann, W.; Böhmer, C.; Gitter, F.; Heimann, F.; Heller, I.; Läuchli, B.; Putz, C. Defining biominerals and organominerals: Direct and indirect indicators of life. Perry et al., Sedimentary Geology, 201, 157–179. Sediment. Geol. 2009, 213, 150–151. [Google Scholar] [CrossRef]
- Défarge, C.; Gautret, P.; Reitner, J.; Trichet, J. Defining organominerals: Comment on ‘Defining biominerals and organominerals: Direct and indirect indicators of life’ by Perry et al. (2007, Sedimentary Geology, 201, 157–179). Sediment. Geol. 2009, 213, 152–155. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Knoll, A.H. Stromatolites in Precambrain carbonates: Evolutionary Mileposts or Environmental Dipsticks? Annu. Rev. Earth Planet. Sci. 1999, 27, 313–358. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.T.; Reid, R.P.; Bebout, B.M.; Hoeft, S.E.; Thompson, A.T., Jr. Formation of lithified micritic laminae in modern marine stromatolites (Bahamas): The role of sulfur cycling. Am. Mineral. 1998, 83, 1482–1493. [Google Scholar] [CrossRef]
- Pentecost, A. Travertine; Springer: Dordrecht, The Netherlands, 2005; ISBN 978-1-4020-3523-4. [Google Scholar]
- Della Porta, G.D. Carbonate build-ups in lacustrine, hydrothermal and fluvial settings: Comparing depositional geometry, fabric types and geochemical signature. Geol. Soc. Lond. Spec. Publ. 2015, 418, SP418.4. [Google Scholar] [CrossRef]
- Visscher, P.T.; Stolz, J.F. Microbial mats as bioreactors: Populations, processes, and products. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 87–100. [Google Scholar] [CrossRef]
- Eymard, I.; Bilmes, A.; Alvarez, M.D.P.; Feo, R.; Hunger, G.; Vasconcelos, C.; Arizteguí, D. Growth morphologies and plausible stressors ruling the formation of Late Pleistocene lacustrine carbonate buildups in the Maquinchao Basin (Argentina). Depos. Rec. 2019, 5, 498–514. [Google Scholar] [CrossRef]
- Pacton, M.; Hunger, G.; Martinuzzi, V.; Cusminsky, G.; Burdin, B.; Barmettler, K.; Vasconcelos, C.; Ariztegui, D. Organomineralization processes in freshwater stromatolites: A living example from eastern Patagonia. Depos. Rec. 2015, 1, 130–146. [Google Scholar] [CrossRef]
- Bilmes, A.; D’Elia, L.; Lopez, L.; Richiano, S.; Varela, A.; del Pilar Alvarez, M.; Bucher, J.; Eymard, I.; Muravchik, M.; Franzese, J.; et al. Digital outcrop modelling using “structure-from- motion” photogrammetry: Acquisition strategies, validation and interpretations to different sedimentary environments. J. S. Am. Earth Sci. 2019, 96, 102325. [Google Scholar] [CrossRef]
- Visscher, P.T.; Reid, R.P.; Bebout, B.M. Microscale observations of sulfate reduction: Correlation of microbial activity with lithified micritic laminae in modern marine stromatolites. Geology 2000, 28, 919–922. [Google Scholar] [CrossRef]
- Bathurst, R.G.C. Boring algae, micrite envelopes and lithification of molluscan biosparites. Geol. J. 1966, 5, 15–32. [Google Scholar] [CrossRef]
- Payandi-Rolland, D.; Roche, A.; Vennin, E.; Visscher, P.T.; Amiotte-Suchet, P.; Thomas, C.; Bundeleva, I.A. Carbonate Precipitation in Mixed Cyanobacterial Biofilms Forming Freshwater Microbial Tufa. Minerals 2019, 9, 409. [Google Scholar] [CrossRef]
- Freytet, P.; Verrecchia, E.P. Freshwater organisms that build stromatolites: A synopsis of biocrystallization by prokaryotic and eukaryotic algae. Sedimentology 1998, 45, 535–563. [Google Scholar] [CrossRef]
- Gradziński, M. Factors controlling growth of modern tufa: Results of a field experiment. Geol. Soc. Lond. Spec. Publ. 2010, 336, 143–191. [Google Scholar] [CrossRef]
- Pedley, M. Freshwater (phytoherm) reefs: The role of biofilms and their bearing on marine reef cementation. Sediment. Geol. 1992, 79, 255–274. [Google Scholar] [CrossRef]
- Pedley, M. Ambient Temperature Freshwater Microbial Tufas. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 179–186. ISBN 978-3-662-04036-2. [Google Scholar]
- Pedley, H.M. Classification and environmental models of cool freshwater tufas. Sediment. Geol. 1990, 68, 143–154. [Google Scholar] [CrossRef]
- Barbin, V.; Schvoerer, M. Cathodoluminescence géosciences. Comptes Rendus de l’Académie des Sci. Ser. IIA Earth Planet. Sci. 1997, 325, 157–169. [Google Scholar] [CrossRef]
- Machel, H.G. Application of Cathodoluminescence to Carbonate Diagenesis. In Cathodoluminescence in Geosciences; Pagel, M., Barbin, V., Blanc, P., Ohnenstetter, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 271–301. ISBN 978-3-662-04086-7. [Google Scholar]
- Merz-Preiß, M.; Riding, R. Cyanobacterial tufa calcification in two freshwater streams: Ambient environment, chemical thresholds and biological processes. Sediment. Geol. 1999, 126, 103–124. [Google Scholar] [CrossRef]
- Gomez, F.J.; Mlewski, C.; Boidi, F.J.; Farías, M.E.; Gérard, E. Calcium Carbonate Precipitation in Diatom-rich Microbial Mats: The Laguna Negra Hypersaline Lake, Catamarca, Argentina. J. Sediment. Res. 2018, 88, 727–742. [Google Scholar] [CrossRef]
- Gomez, F.J.; Kah, L.C.; Bartley, J.K.; Astini, R.A. Microbialites in a High-Altitude Andean Lake: Multiple Controls on Carbonate Precipitation and Lamina Accretion. PALAIOS 2014, 29, 233–249. [Google Scholar] [CrossRef]
- Wacey, D.; Urosevic, L.; Saunders, M.; George, A.D. Mineralisation of filamentous cyanobacteria in Lake Thetis stromatolites, Western Australia. Geobiology 2018, 16, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Dupraz, C.; Visscher, P.T. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar] [CrossRef]
- Vasconcelos, C.; Dittrich, M.; McKenzie, J.A. Evidence of microbiocoenosis in the formation of laminae in modern stromatolites. Facies 2014, 60, 3–13. [Google Scholar] [CrossRef]
- Mattson, M.D. Alkalinity of Freshwater. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-409548-9. [Google Scholar]
- Burne, R.V.; Moore, L.S.; Christy, A.G.; Troitzsch, U.; King, P.L.; Carnerup, A.M.; Hamilton, P.J. Stevensite in the modern thrombolites of Lake Clifton, Western Australia: A missing link in microbialite mineralization? Geology 2014, 42, 575–578. [Google Scholar] [CrossRef]
- Pace, A.; Bourillot, R.; Bouton, A.; Vennin, E.; Galaup, S.; Bundeleva, I.; Patrier, P.; Dupraz, C.; Thomazo, C.; Sansjofre, P.; et al. Microbial and diagenetic steps leading to the mineralisation of Great Salt Lake microbialites. Sci. Rep. 2016, 6, 31495. [Google Scholar] [CrossRef]
- Wacey, D.; Gleeson, D.; Kilburn, M.R. Microbialite taphonomy and biogenicity: New insights from NanoSIMS. Geobiology 2010, 8, 403–416. [Google Scholar] [CrossRef]
- Bontognali, T.R.R.; Vasconcelos, C.; Warthmann, R.J.; Bernasconi, S.M.; Dupraz, C.; Strohmenger, C.J.; McKenzie, J.A. Dolomite formation within microbial mats in the coastal sabkha of Abu Dhabi (United Arab Emirates). Sedimentology 2010, 57, 824–844. [Google Scholar] [CrossRef]
- Zeyen, N.; Benzerara, K.; Li, J.; Groleau, A.; Balan, E.; Robert, J.-L.; Estève, I.; Tavera, R.; Moreira, D.; López-García, P. Formation of low-T hydrated silicates in modern microbialites from Mexico and implications for microbial fossilization. Front. Earth Sci. 2015, 3. [Google Scholar] [CrossRef]
- Souza-Egipsy, V.; Wierzchos, J.; Ascaso, C.; Nealson, K.H. Mg–silica precipitation in fossilization mechanisms of sand tufa endolithic microbial community, Mono Lake (California). Chem. Geol. 2005, 217, 77–87. [Google Scholar] [CrossRef]
- Farías, M.E.; Poiré, D.G.; Arrouy, M.J.; Albarracin, V.H. Modern Stromatolite Ecosystems at Alkaline and Hypersaline High-Altitude Lakes in the Argentinean Puna. In STROMATOLITES: Interaction of Microbes with Sediments; Tewari, V., Seckbach, J., Eds.; Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 2011; pp. 427–441. ISBN 978-94-007-0397-1. [Google Scholar]
- Bruggmann, S. Growth Constraints and Environmental Influences on a Modern Stromatolite, Lagoa Vermelha, Brasil. Master’s Thesis, ETH-Zürich, Zürich, Switzerland, 2015. [Google Scholar]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eymard, I.; Alvarez, M.d.P.; Bilmes, A.; Vasconcelos, C.; Ariztegui, D. Tracking Organomineralization Processes from Living Microbial Mats to Fossil Microbialites. Minerals 2020, 10, 605. https://doi.org/10.3390/min10070605
Eymard I, Alvarez MdP, Bilmes A, Vasconcelos C, Ariztegui D. Tracking Organomineralization Processes from Living Microbial Mats to Fossil Microbialites. Minerals. 2020; 10(7):605. https://doi.org/10.3390/min10070605
Chicago/Turabian StyleEymard, Inès, María del Pilar Alvarez, Andrés Bilmes, Crisogono Vasconcelos, and Daniel Ariztegui. 2020. "Tracking Organomineralization Processes from Living Microbial Mats to Fossil Microbialites" Minerals 10, no. 7: 605. https://doi.org/10.3390/min10070605
APA StyleEymard, I., Alvarez, M. d. P., Bilmes, A., Vasconcelos, C., & Ariztegui, D. (2020). Tracking Organomineralization Processes from Living Microbial Mats to Fossil Microbialites. Minerals, 10(7), 605. https://doi.org/10.3390/min10070605