

P1 Component Discloses Gender-Related Hemispheric Asymmetries during IAPS Processing


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Behavioral Measures
2.3. Experimental Setting
2.4. EEG Preprocessing
2.5. Behavioral Statistical Analysis
2.6. ERP Analysis
2.7. EEG Source Localization
3. Results
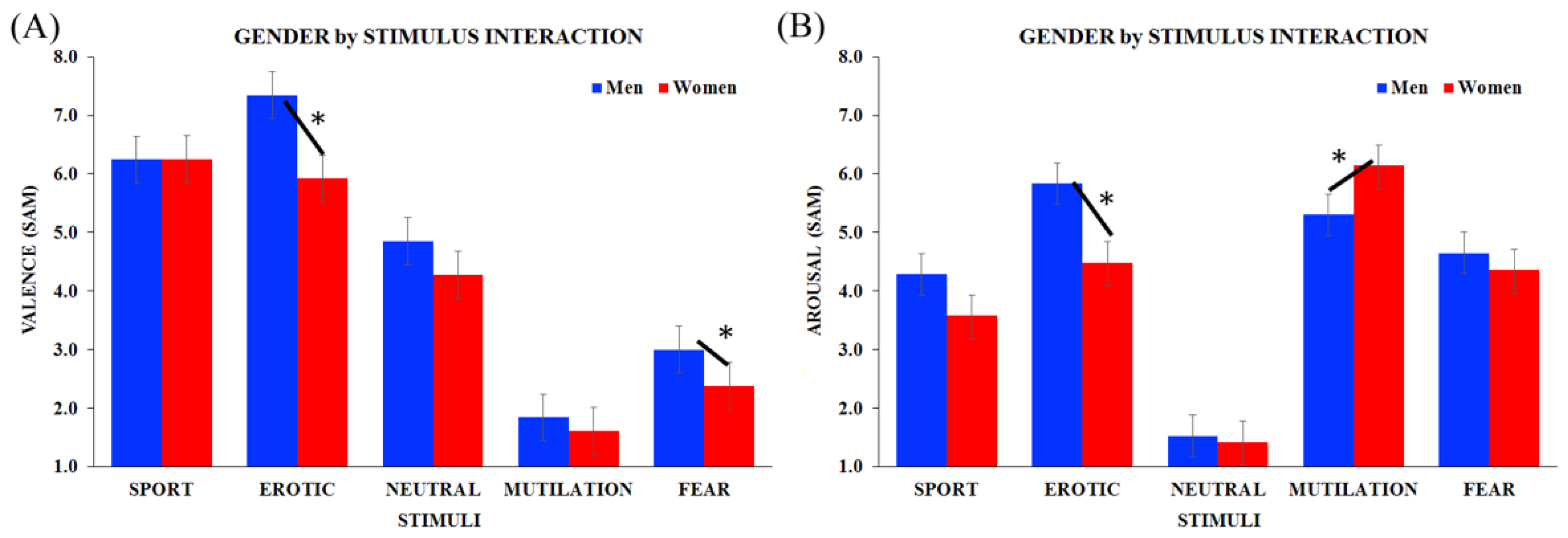
3.1. Empathy Traits, Valence and Arousal
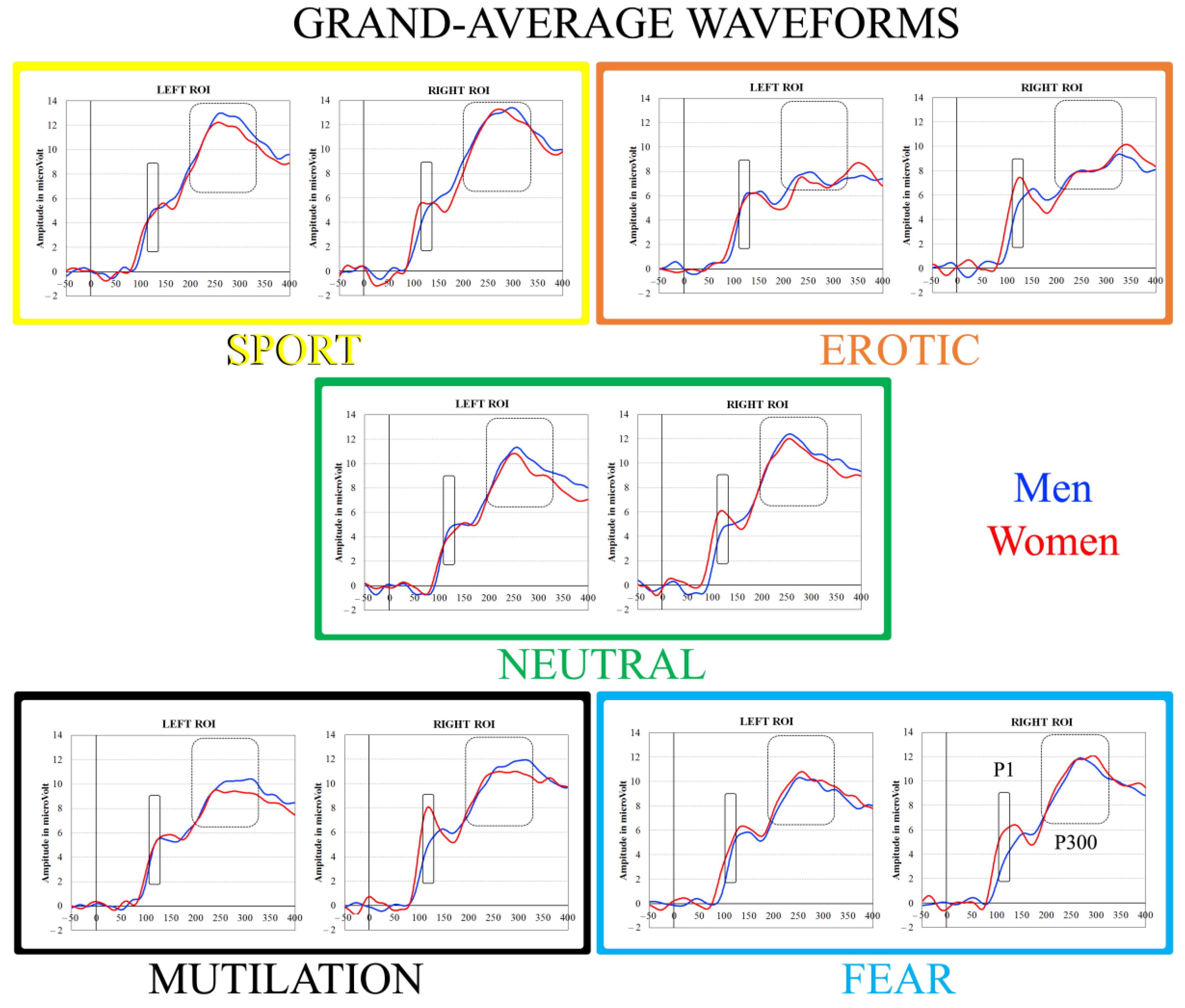
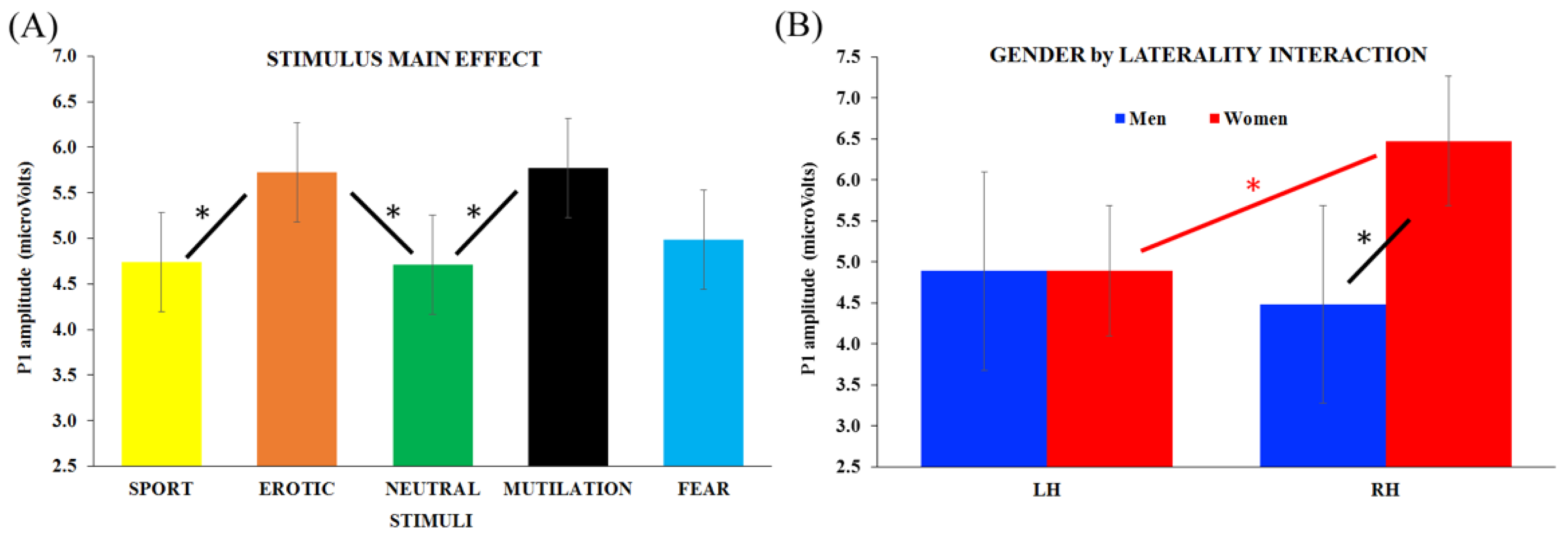
3.2. ERP Analysis: P1 Component
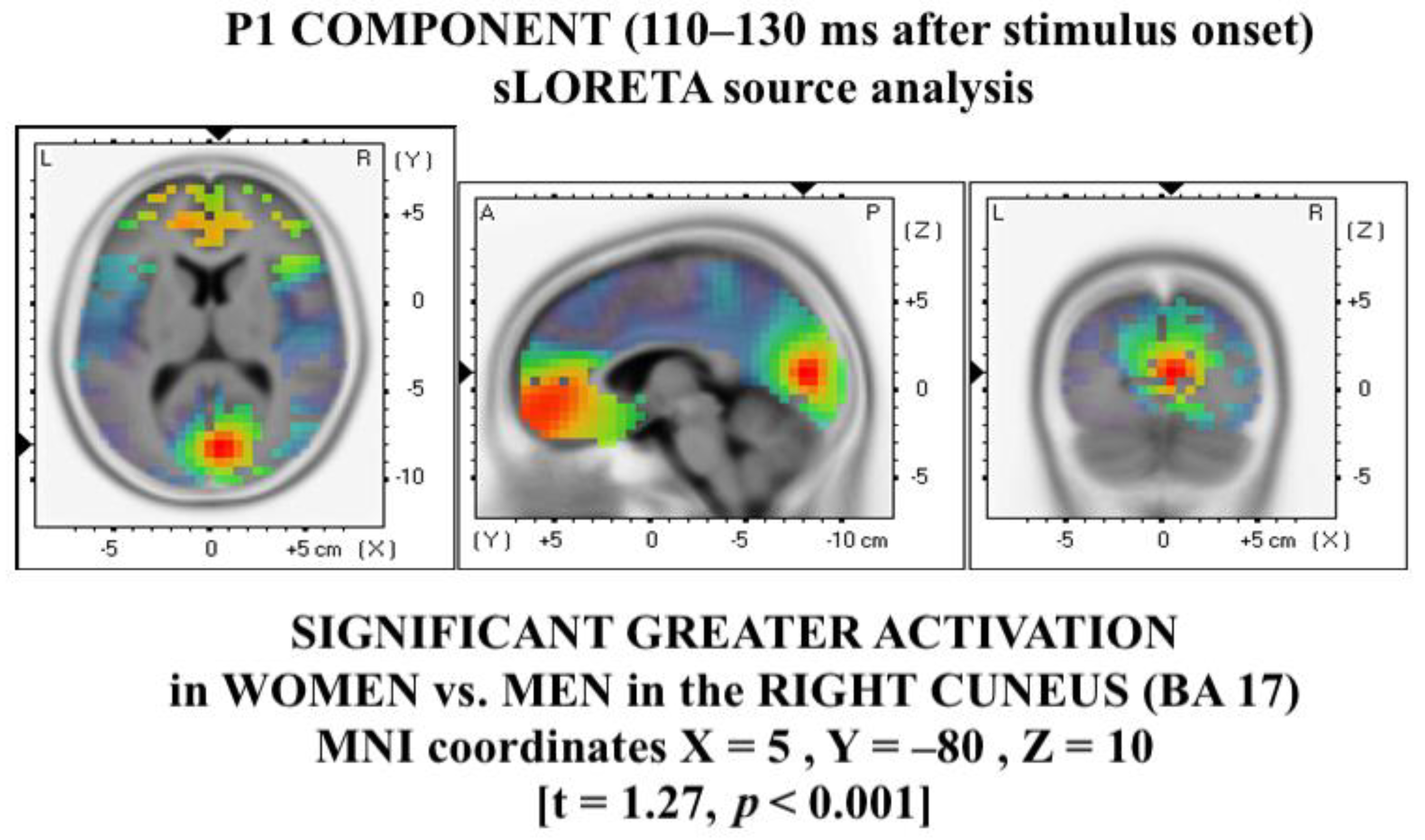
3.3. Source Analysis on P1 Components
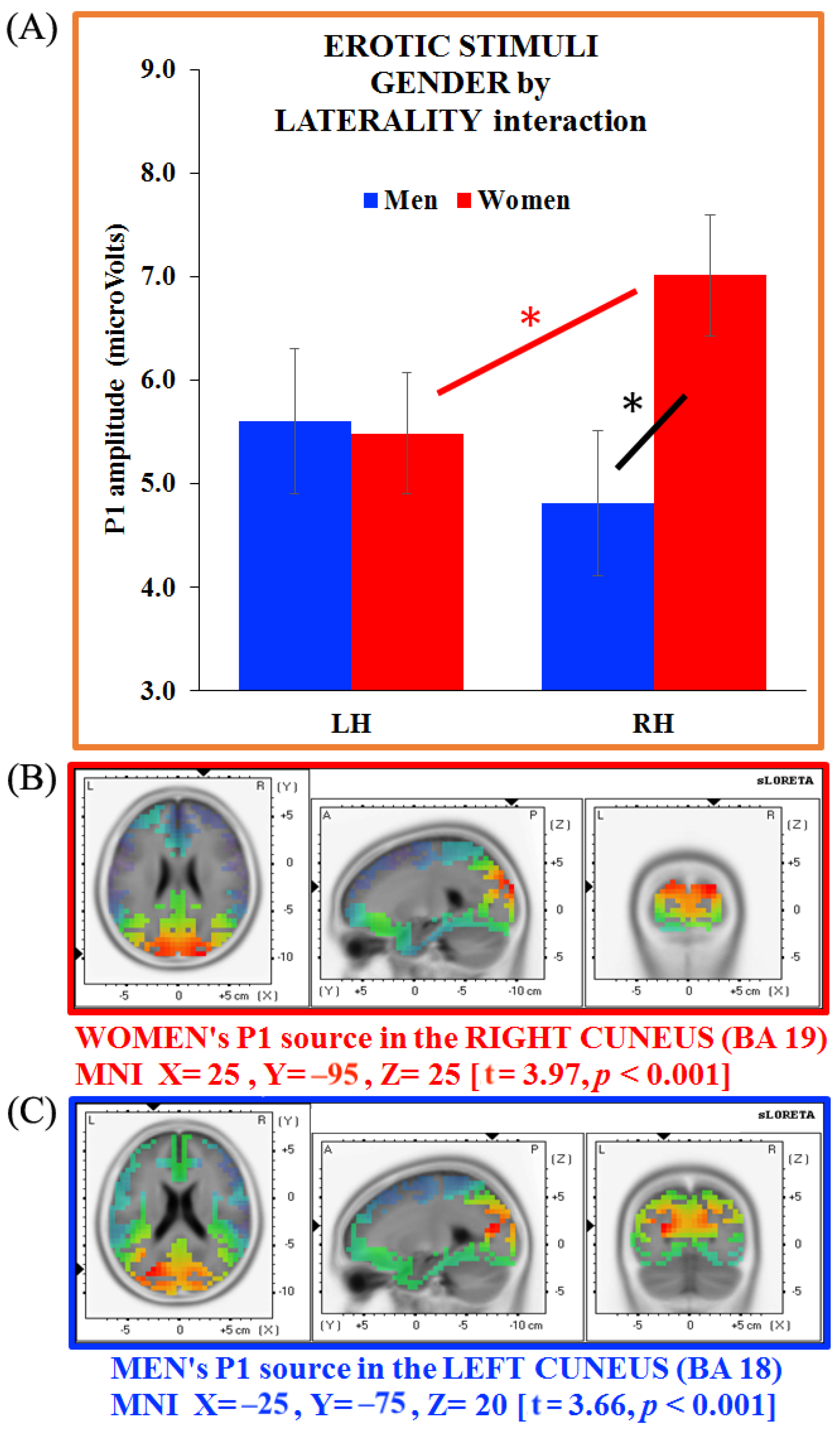
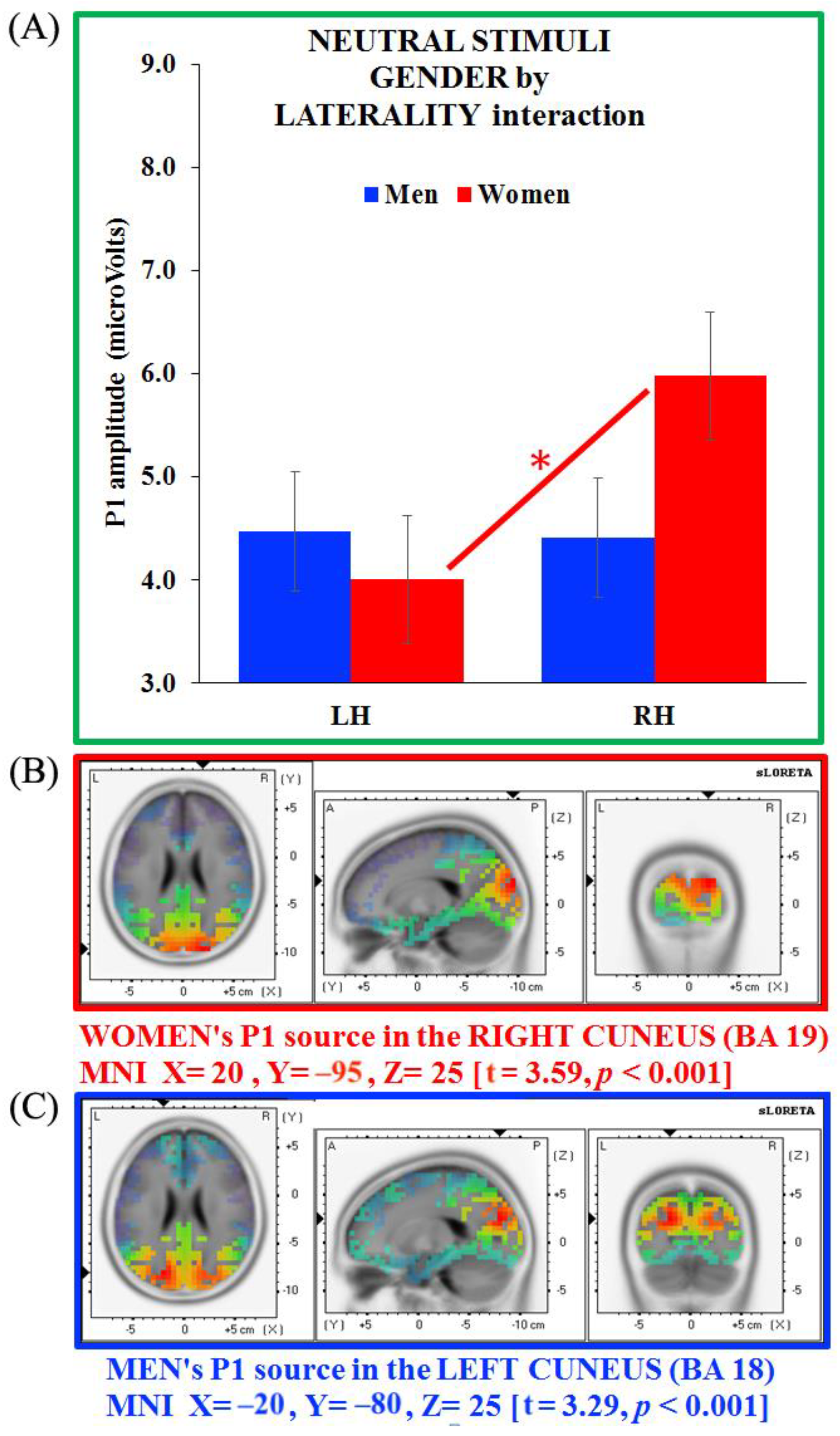
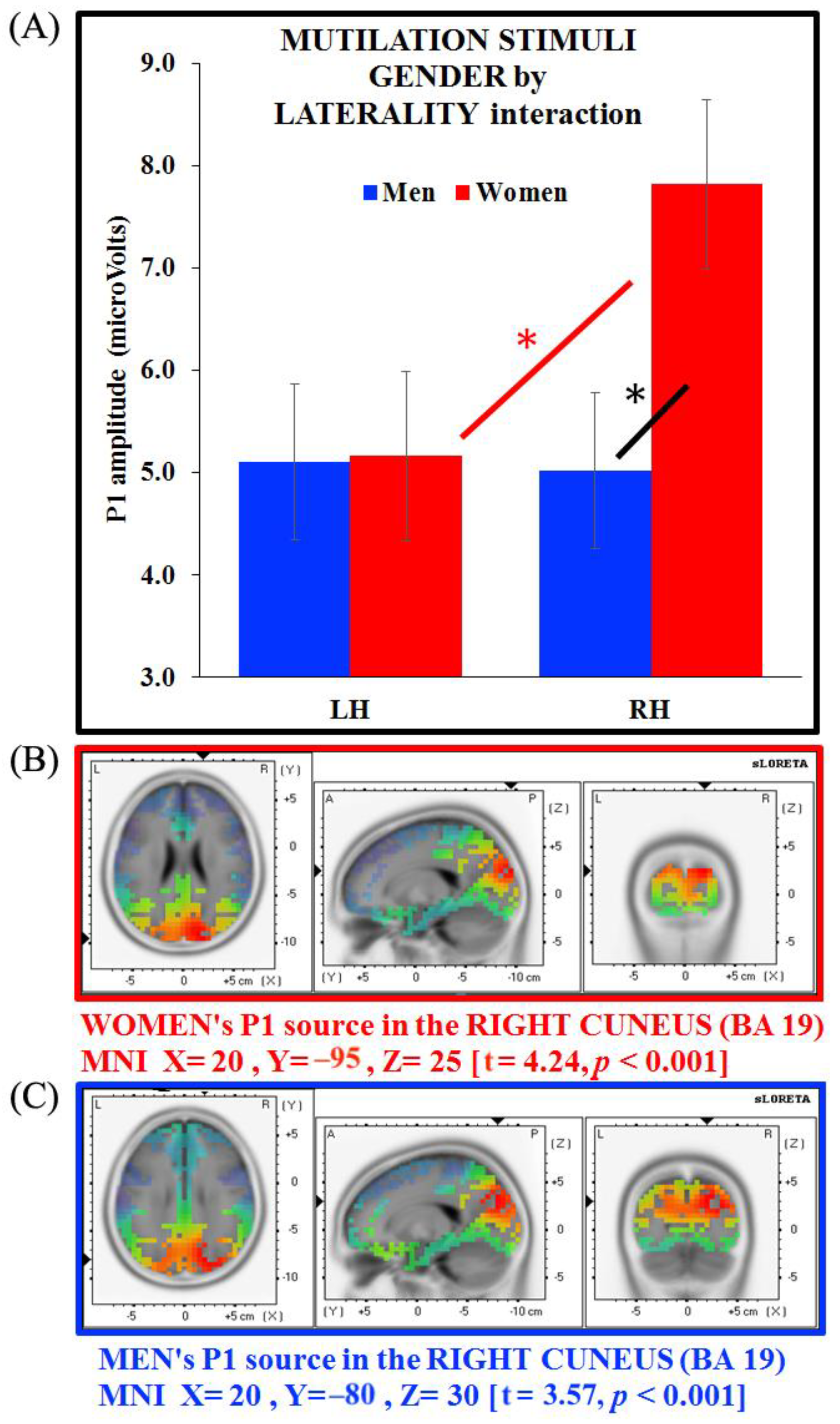
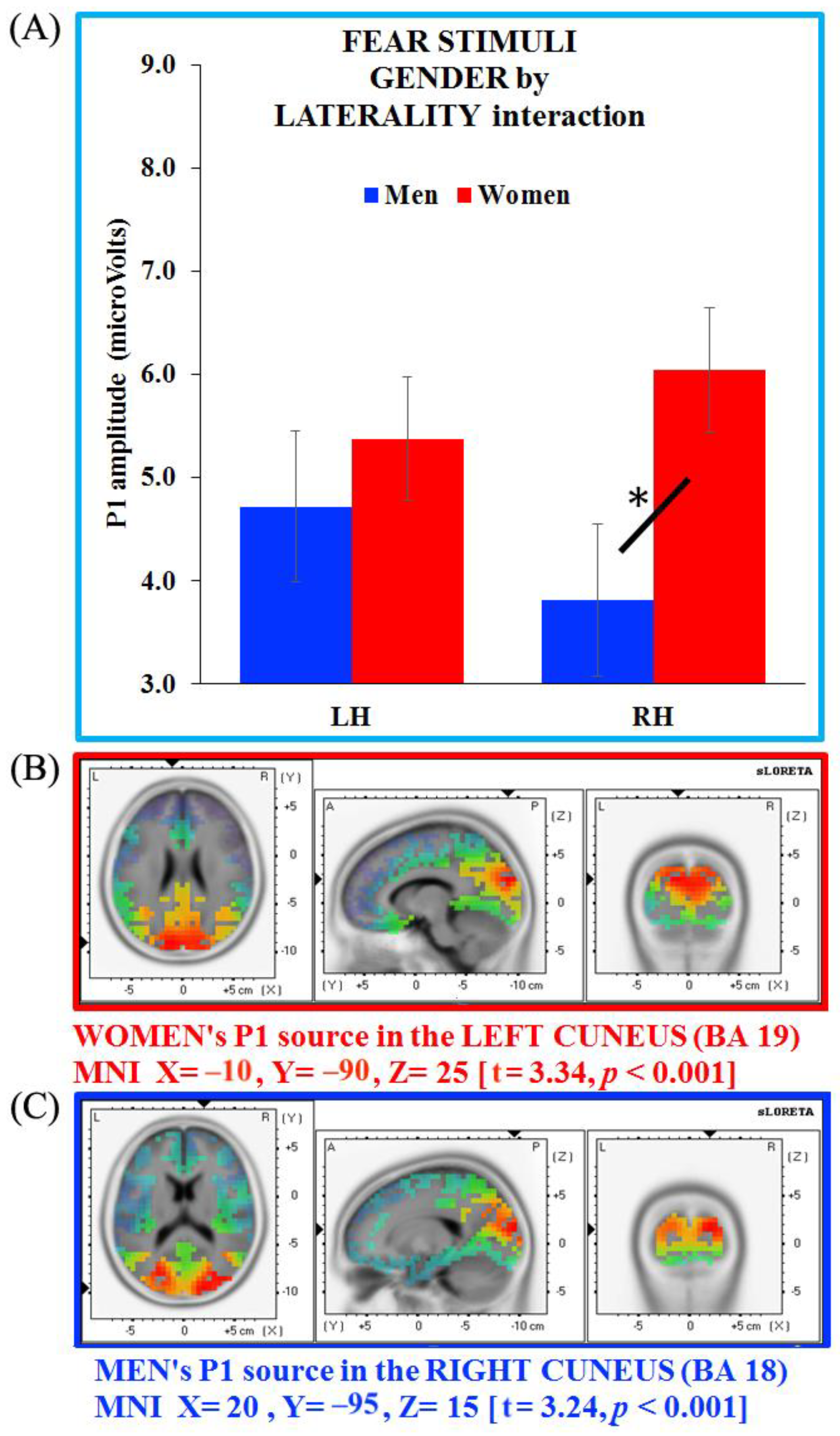
3.4. Separate ANOVAs and Source Analysis on Emotional Stimuli (P1 Component)
3.5. ERP Analysis: P300 Component
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Romeo, Z.; Fusina, F.; Semenzato, L.; Bonato, M.; Angrilli, A.; Spironelli, C. Comparison of Slides and Video Clips as Different Methods for Inducing Emotions: An Electroencephalographic Alpha Modulation Study. Front. Hum. Neurosci. 2022, 16, 901422. [Google Scholar] [CrossRef]
- Alves, N.T.; Aznar-Casanova, J.A.; Fukusima, S.S. Patterns of brain asymmetry in the perception of positive and negative facial expressions. Laterality 2009, 14, 256–272. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Bradley, M.; Cuthbert, B.N. International affective picture system (IAPS): Technical manual and affective ratings. NIMH Cent. Study Emot. Atten. 1997, 1, 39–58. [Google Scholar]
- Maddock, R.J.; Garrett, A.S.; Buonocore, M.H. Posterior cingulate cortex activation by emotional words: fMRI evidence from a valence decision task. Hum. Brain Mapp. 2003, 18, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Moser, E.; Peper, M. fMRI of Emotion. In fMRI Techniques and Protocols; Springer: Berlin/Heidelberg, Germany, 2016; Volume 119, pp. 451–494. [Google Scholar]
- Rahman, M.M.; Sarkar, A.K.; Hossain, M.A.; Hossain, M.S.; Islam, M.R.; Hossain, M.B.; Quinn, J.M.W.; Moni, M.A. Recognition of human emotions using EEG signals: A review. Comput. Biol. Med. 2021, 136, 104696. [Google Scholar] [CrossRef]
- Bianchin, M.; Angrilli, A. Gender differences in emotional responses: A psychophysiological study. Physiol. Behav. 2012, 105, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, J.K.; Nordin, S.; Sequeira, H.; Polich, J. Affective picture processing: An integrative review of ERP findings. Biol. Psychol. 2008, 77, 247–265. [Google Scholar] [CrossRef]
- Ding, R.; Li, P.; Wang, W.; Luo, W. Emotion Processing by ERP Combined with Development and Plasticity. Neural Plast. 2017, 2017, 5282670. [Google Scholar] [CrossRef]
- Carretié, L.; Mercado, F.; Tapia, M.; Hinojosa, J.A. Hinojosa Emotion, attention, and the ‘negativity bias’, studied through event-related potentials. Int. J. Psychophysiol. 2001, 41, 75–85. [Google Scholar] [CrossRef]
- Carretié, L.; Martín-Loeches, M.; Hinojosa, J.A.; Mercado, F. Emotion and attention interaction studied through event-related potentials. J. Cogn. Neurosci. 2001, 13, 1109–1128. [Google Scholar] [CrossRef]
- Conroy, M.A.; Polich, J. Affective valence and P300 when stimulus arousal level is controlled. Cogn. Emot. 2007, 21, 891–901. [Google Scholar] [CrossRef]
- Schupp, H.T.; Cuthbert, B.N.; Bradley, M.; Cacioppo, J.T.; Tiffany, I.; Lang, P.J. Affective picture processing: The late positive potential is modulated by motivational relevance. Psychophysiology 2000, 37, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, B.N.; Schupp, H.T.; Bradley, M.; Birbaumer, N.; Lang, P.J. Brain potentials in affective picture processing: Covariation with autonomic arousal and affective report. Biol. Psychol. 2000, 52, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Hajcak, G.; Macnamara, A.; Olvet, D.M. Event-related potentials, emotion, and emotion regulation: An integrative review. Dev. Neuropsychol. 2010, 35, 129–155. [Google Scholar] [CrossRef] [PubMed]
- Mini, A.; Palomba, D.; Angrilli, A.; Bravi, S. Emotional Information Processing and Visual Evoked Brain Potentials. Percept. Mot. Skills 1996, 83, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Palomba, D.; Angrilli, A.; Mini, A. Visual evoked potentials, heart rate responses and memory to emotional pictorial stimuli. Int. J. Psychophysiol. 1997, 27, 55–67. [Google Scholar] [CrossRef]
- Cahill, L. Why sex matters for neuroscience. Nat. Rev. Neurosci. 2006, 7, 477–484. [Google Scholar] [CrossRef]
- Kret, M.E.; De Gelder, B. A review on sex differences in processing emotional signals. Neuropsychologia 2012, 50, 1211–1221. [Google Scholar] [CrossRef]
- Davidson, R.J. Anxiety and affective style: Role of prefrontal cortex and amygdala. Biol. Psychiatry 2002, 51, 68–80. [Google Scholar] [CrossRef]
- Bradley, M.; Codispoti, M.; Sabatinelli, D.; Lang, P.J. Emotion and motivation II: Sex differences in picture processing. Emotion 2001, 1, 300–319. [Google Scholar] [CrossRef]
- Chivers, M.L.; Seto, M.C.; Lalumière, M.L.; Laan, E.; Grimbos, T. Agreement of self-reported and genital measures of sexual arousal in men and women: A meta-analysis. Arch. Sex. Behav. 2010, 39, 5–56. [Google Scholar] [CrossRef] [PubMed]
- Lithari, C.; Frantzidis, C.A.; Papadelis, C.; Vivas, A.B.; Klados, M.A.; Kourtidou-Papadeli, C.; Pappas, C.; Ioannides, A.A.; Bamidis, P.D. Are females more responsive to emotional stimuli? A neurophysiological study across arousal and valence dimensions. Brain Topogr. 2010, 23, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Proverbio, A.M.; Adorni, R.; Zani, A.; Trestianu, L. Sex differences in the brain response to affective scenes with or without humans. Neuropsychologia 2009, 47, 2374–2388. [Google Scholar] [CrossRef] [PubMed]
- Groen, Y.; Wijers, A.A.; Tucha, O.; Althaus, M. Are there sex differences in ERPs related to processing empathy-evoking pictures? Neuropsychologia 2013, 51, 142–155. [Google Scholar] [CrossRef]
- Gasbarri, A.; Arnone, B.; Pompili, A.; Pacitti, F.; Pacitti, C.; Cahill, L. Sex-related hemispheric lateralization of electrical potentials evoked by arousing negative stimuli. Brain Res. 2007, 1138, 178–186. [Google Scholar] [CrossRef]
- Luck, S.J.; Woodman, G.F.; Vogel, E.K. Event-related potential studies of attention. Trends Cogn. Sci. 2000, 4, 432–440. [Google Scholar] [CrossRef]
- Müller-Bardorff, M.; Bruchmann, M.; Mothes-Lasch, M.; Zwitserlood, P.; Schlossmacher, I.; Hofmann, D.; Miltner, W.; Straube, T. Early brain responses to affective faces: A simultaneous EEG-fMRI study. Neuroimage 2018, 178, 660–667. [Google Scholar] [CrossRef]
- Schindler, S.; Bublatzky, F. Attention and emotion: An integrative review of emotional face processing as a function of attention. Cortex 2020, 130, 362–386. [Google Scholar] [CrossRef]
- Denefrio, S.; Simmons, A.; Jha, A.; Dennis-Tiwary, T.A. Emotional cue validity effects: The role of neurocognitive responses to emotion. PLoS ONE 2017, 12, e0179714. [Google Scholar] [CrossRef]
- Smith, N.K.; Cacioppo, J.T.; Larsen, J.T.; Chartrand, T.L. May I have your attention, please: Electrocortical responses to positive and negative stimuli. Neuropsychologia 2003, 41, 171–183. [Google Scholar] [CrossRef]
- Spironelli, C.; Romeo, Z.; Maffei, A.; Angrilli, A. Comparison of automatic visual attention in schizophrenia, bipolar disorder, and major depression: Evidence from P1 event-related component. Psychiatry Clin. Neurosci. 2019, 73, 331–339. [Google Scholar] [CrossRef]
- Davis, M.H. Measuring individual differences in empathy: Evidence for a multidimensional approach. J. Pers. Soc. Psychol. 1983, 44, 113–126. [Google Scholar] [CrossRef]
- Wolpe, J.; Lang, P.J. A fear survey schedule for use in behaviour therapy. Behav. Res. Ther. 1964, 2, 27–30. [Google Scholar] [CrossRef]
- Spielberger, C.D.; Gorsuch, R.L.; Lushene, R.; Vagg, P.R.; Jacobs, G.A. Manual For the State-Trait Anxiety Inventory; Consulting Psychologists Press: Palo Alto, CA, USA, 1983. [Google Scholar]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and Validation of Brief Measures of Positive and Negative Affect: The PANAS Scales. J. Pers. Soc. Psychol. 1988, 54, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Tadel, F.; Bock, E.; Niso, G.; Mosher, J.C.; Cousineau, M.; Pantazis, D.; Leahy, R.M.; Baillet, S. MEG/EEG group analysis with brainstorm. Front. Neurosci. 2019, 13, 76. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Esslen, M.; Kochi, K.; Lehmann, D. Functional imaging with low-resolution brain electromagnetic tomography (LORETA): A review. Methods Find. Exp. Clin. Pharmacol. 2002, 24, 91–95. [Google Scholar]
- Maffei, A.; Spironelli, C.; Angrilli, A. Affective and cortical EEG gamma responses to emotional movies in women with high vs low traits of empathy. Neuropsychologia 2019, 133, 107175. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.P.; Pinheiro, A.P.; Costa, A.; Frade, C.S.; Comesaña, M.; Pureza, R. Adaptation of the International Affective Picture System (IAPS) for European Portuguese. Behav. Res. Methods 2014, 47, 1159–1177. [Google Scholar] [CrossRef] [PubMed]
- Bradley, M.; Lang, P.J. The International Affective Digitized Sounds (IADS): Stimuli, Instruction Manual and Affective Ratings; The Center for Research in Psychophysiology, University of Florida: Gainesville, FL, USA, 1999. [Google Scholar]
- Poláčková Šolcová, I.; Lačev, A. Differences in male and female subjective experience and physiological reactions to emotional stimuli. Int. J. Psychophysiol. 2017, 117, 75–82. [Google Scholar] [CrossRef]
- Clements, A.M.; Rimrodt, S.L.; Abel, J.R.; Blankner, J.G.; Mostofsky, S.H.; Pekar, J.J.; Denckla, M.B.; Cutting, L.E. Sex differences in cerebral laterality of language and visuospatial processing. Brain Lang. 2006, 98, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Vanston, J.E.; Strother, L. Sex differences in the human visual system. J. Neurosci. Res. 2017, 95, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Bradley, M.; Fitzsimmons, J.R.; Cuthbert, B.N.; Scott, J.D.; Moulder, B.; Nangia, V. Emotional arousal and activation of the visual cortex: An fMRI analysis. Psychophysiology 1998, 35, 199–210. [Google Scholar] [CrossRef]
- Stevens, W.D.; Kahn, I.; Wig, G.S.; Schacter, D.L. Hemispheric asymmetry of visual scene processing in the human brain: Evidence from repetition priming and intrinsic activity. Cereb. Cortex 2012, 22, 1935–1949. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xin, X.; Zhu, H.; Li, F.; Xiong, H.; Zhang, T.; Lai, Y. A comparative study on the dynamic EEG center of mass with different references. Front. Neurosci. 2017, 11, 509. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romeo, Z.; Spironelli, C. P1 Component Discloses Gender-Related Hemispheric Asymmetries during IAPS Processing. Symmetry 2023, 15, 610. https://doi.org/10.3390/sym15030610
Romeo Z, Spironelli C. P1 Component Discloses Gender-Related Hemispheric Asymmetries during IAPS Processing. Symmetry. 2023; 15(3):610. https://doi.org/10.3390/sym15030610
Chicago/Turabian StyleRomeo, Zaira, and Chiara Spironelli. 2023. "P1 Component Discloses Gender-Related Hemispheric Asymmetries during IAPS Processing" Symmetry 15, no. 3: 610. https://doi.org/10.3390/sym15030610
APA StyleRomeo, Z., & Spironelli, C. (2023). P1 Component Discloses Gender-Related Hemispheric Asymmetries during IAPS Processing. Symmetry, 15(3), 610. https://doi.org/10.3390/sym15030610