3.1. Tautomers–rotamers Selected for Neutral Bases
Among twenty-one isomers possible for nine tautomers of neutral
C and
iC [
6,
7], fourteen structures (
Figure 3) possessing the labile protons at N and O atoms have been selected in this work. They form the isomeric mixture corresponding to AH
2B in acid-base equilibria (
Figure 2). For tautomers possessing exo −OH and/or exo =NH, two extreme conformational and/or configurational isomers (
a and
b) have been considered, respectively. The CH-isomers containing one labile proton at the endo C5 atom (see in
Table 1) have been omitted, owing to their high relative Gibbs energies (Δ
G > 10 kcal mol
−1) estimated in the gas phase at the DFT(B3LYP)/6-311+G(d,p) level [
6,
7]. Taking into account the well-recognized push–pull effects in the tautomeric amide-iminol {O=C(R)−NH− ⇄ HO−C(R)=N−) and amidine (−N=C(R)−NH
2 ⇄ −HN−C(R)=NH} parts, the candidate site of protonation and deprotonation can be easily indicated without calculations. In the protonation reaction, the sp
2 hybridized =O and/or =N− can gain a proton, whereas >NH, −NH
2, and/or −OH containing the sp
3 hybridized heteroatoms can lose the labile proton. In
Figure 3, the selected sites of protonation and deprotonation in the fourteen selected isomer of
C and
iC have been indicated by red and blue arrows, respectively.
The values of Δ
G included in
Figure 3 show variations in isomeric stability resulting not only from prototropy, conformational, and configurational isomerism, but also from possible intramolecular interactions (favorable or unfavorable) between the exo and endo groups. Larger variations are caused by prototropy (from 0 to 33 kcal mol
−1) than by conformational and configurational isomerism (from 1 to 9 kcal mol
−1). The smallest effects take place for these
a and
b pairs, for which the same type of interactions are possible (favorable or unfavorable in both isomers). The highest effects occur when a favorable interaction in one isomer changes into an unfavorable repulsion in the other one and vice versa.
Additional perusal of Δ
Gs estimated for all selected isomers shows some analogy in the isomeric preferences for cytosine and isocytosine. Only six tautomers–rotamers of neutral cytosine (
C-13a,
C-13b,
C-18,
C-38,
C-78a, and
C-78b) and only six tautomers–rotamers of neutral isocytosine (
iC-13a,
iC-13b,
iC-17,
iC-37,
iC-78a, and
iC-78b) possess Δ
G ≤ 10 kcal mol
−1. This indicates that the isomeric mixture of neutral
C and
iC can consist mainly of these six isomers (two imino-oxo, two amino-oxo, and two amino-hydroxy forms). Taking the Mezey et al. rule [
37,
38] into account, the other isomers of neutral
C and
iC can be neglected in the isomeric mixtures in the gas phase (or non-polar environment). The canonical
C-18 isomer is the major form for cytosine, and the other ones can be considered as minor or rare forms. In the case of isocytosine, the DFT-calculated canonical
iC-37 isomer has slightly lower Gibbs energy than
iC-78a. However, both isomers can be treated as major forms. The other isomers are minor or rare forms.
It should be mentioned here that experimental methods do not give the complete picture on relative stabilities of all possible isomers in tautomeric mixtures. The reasons are as follows: (i) prototropic conversions are frequently very fast and reversible processes, (ii) tautomerism is often very sensitive to the environment, (iii) experimental methods have various limits of identification, (iv) only signals of major isomers (1–100%) of significant intensities can be well detected, (v) signals of minor isomers (0.01–1%) can be too small and difficult to distinguish from the background, and (vi) rare isomers (<0.01 %) are usually undetectable. Hence, the number of identified isomers can be different using different methods such as ultraviolet (UV), infrared (IR), matrix isolation IR, IR laser in helium nanodroplets, Raman, microwave (MV), nuclear magnetic resonance (NMR), mass spectrometry (MS), core-level X-ray photoemission, resonant multi-photon ionization (REMPI), near-edge X-ray absorption, etc.
For example, depending on the method employed for gaseous cytosine, one (
C-18), two (
C-18 and
C-78) or three (
C-18,
C-78, and
C-13) tautomers have been identified [
8,
10,
16,
17,
18]. Five isomers of gaseous cytosine (
C-18, two conformational isomers
a and
b of
C-78, and two configurational isomers
a and
b of
C-13) have only been found by Alonso et al. [
9], who applied a laser ablation molecular beam FT-MW spectroscopy. For crystal cytosine, only the canonical isomer
C-18 has been detected [
23,
24], whereas for hydrated cytosine, the isomer
C-38 has been additionally observed [
15,
19,
20,
21,
22].
In the case of neutral isocytosinie, using various spectroscopic techniques different tautomers have also been identified when proceeding from the gas phase to aqueous solution, and next to the solid state [
7,
11,
12,
13,
14,
15,
25,
26,
27]. The tautomer
iC-78 predominates in vacuo or non-polar environment (e.g., lipids), whereas
iC-37 is favored in aqueous solution. Interestingly, two tautomeric amino-oxo forms (
iC-17 and
iC-37) have been found in the crystal state [
25,
26].
3.2. Isomerism in Deprotonated Bases
Some interesting structures for mono- and di-anionic pyrimidine bases that play an important role in chemistry of organic calixarenes have been reported in the literature by Lippert, Sanz Miguel, Freisinger, and their co-workers [
59,
60]. Acting as N-ligands, like aliphatic di-amines (e.g., ethylenediamine) and aromatic di-imines (e.g., 2,2′-bipyridine), they can form various multinuclear complexes, even structures with mixed-metal (Pt
x, Pd
y), mixed-nucleobase (uracil, cytosine), and mixed-amine (ethylenediamine, 2,2′-bipyridine).
Note that the di-deprotonated forms (AB)
2− of the pyrimidine bases
C and
iC have no labile proton. Thus, no prototropic conversion takes place in them. Only configurational isomerism is possible for the exo =NH group. The geometrical isomers
a and
b of di-deprotonated cytosine (
da-C) and isocytosine (
da-iC), being a consequence of configurational isomerism about the C=N bond, are given in
Figure 4. Their Δ
G values calculated at the DFT level are also included in this figure. We can see that the Δ
G values of the configurational isomers are not very large. They are considerably lower than 10 kcal mol
−1. This means that the two di-anionic isomers
a and
b can be considered in the isomeric mixtures of
da-C-0 and
da-iC-0 with different preference of the isomer
a and
b, respectively. To our knowledge, there is no documented example in the literature for the pyrimidine bases on their di-anionic forms in the gas-phase, and no comparison can be made.
A different situation occurs for the mono-deprotonated pyrimidine bases, mono-anions (AHB)
− of
C and
iC. They contain only one labile proton that can move between five conjugated sites. The number of tautomers for the
C and
iC mono-anions (
ma-C and
ma-iC, respectively) is smaller than that for neutrals because of lower number of the labile protons, and also, because two or more neutral isomers can be deprotonated to the same common mono-anion. Consequently, tautomeric conversions in mono-anions of
C and
iC lead to five possible prototropic tautomers. Like for neutral
C and
iC, the CH tautomers can be omitted in the isomeric mixtures of mono-anions. Their Δ
E values are between 15 and 20 kcal mol
−1 at the AM1 level. Hence, only four tautomers with the labile proton at N or O, i.e., at 1-, 3-, 7-, or 8-position, have been examined at the DFT level. Their conformational and configurational isomers, and also their Δ
Gs are included in
Figure 5.
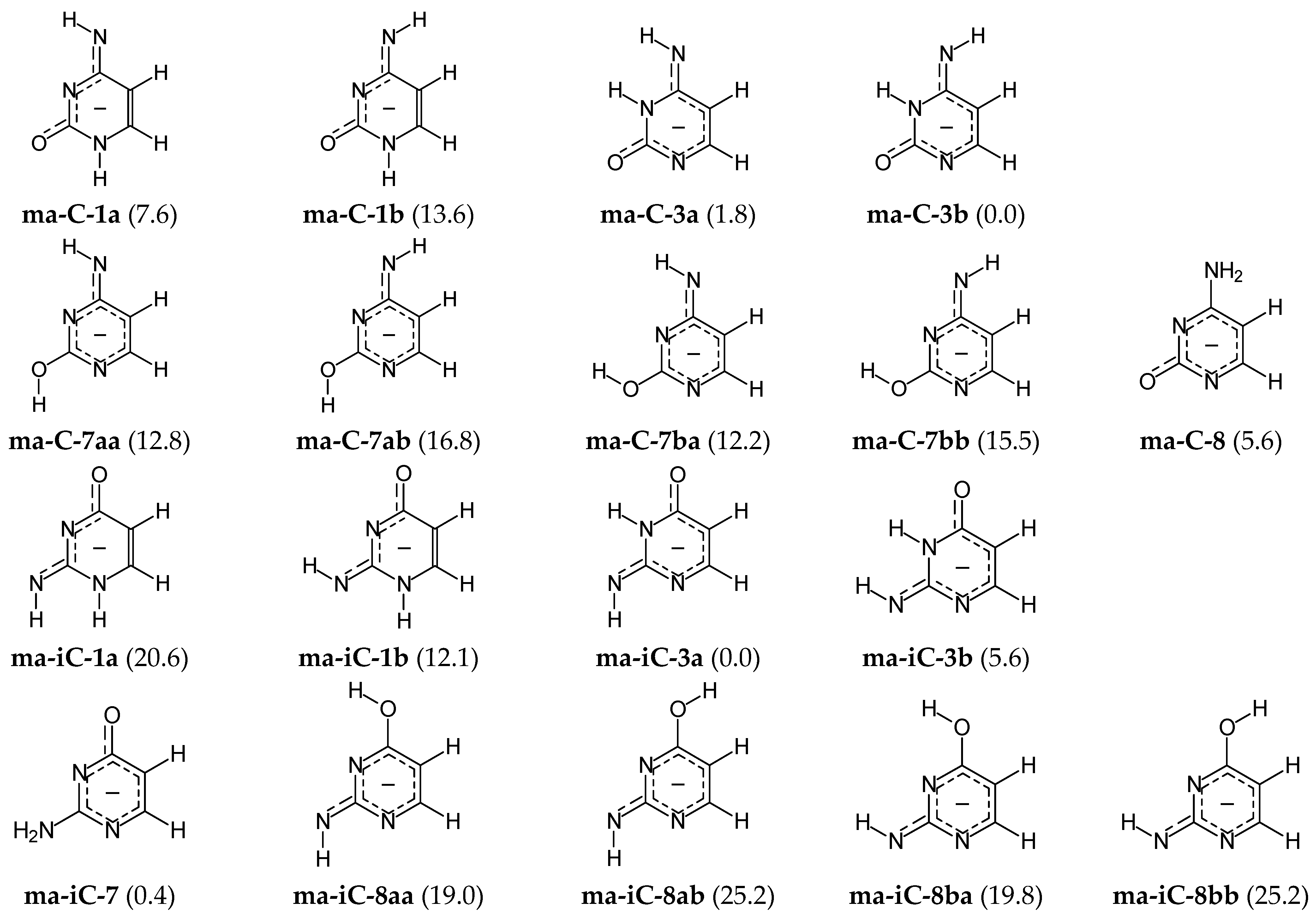
For mono-anionic isomers of cytosine (ma-C), the variations of ΔGs are not very large (0 < ΔG < 17 kcal mol−1), whereas those for isocytosine (ma-iC) are considerably larger (0 < ΔG < 26 kcal mol−1). The difference in stability of the anionic forms involves different number of potential isomers that significantly affect properties of the isomeric mixtures ma-C and ma-iC. Four isomers of mono-anionic cytosine (ma-C-1a, ma-C-3a, ma-C-3b, and ma-C-8) possess ΔGs lower than 10 kcal mol−1, whereas only three isomers (ma-iC-3a, ma-iC-3b, and ma-iC-7) significantly participate in the isomeric mixture of mono-deprotonated isocytosine. The ΔG value (12.1 kcal mol−1) of ma-iC-1b (exhibiting some analogous favorable intramolecular interactions like ma-C-1a) seems to be too high and cannot influence significantly acid-base properties of isocytosine. However, taking into account the analogy with cytosine mono-anion, ma-iC-1b can be considered for the isomeric mixture ma-iC in acid-base equilibria.
DFT calculations show additionally that the favored mono-anionic isomer ma-C-3b cannot be directly formed from the preferred neutral canonical form C-18. Direct deprotonation reaction of C-18 at N1H or N8H leads to three possible isomers ma-C-8, ma-C-1a, and ma-C-1b. All of them possess considerably higher Gibbs energies than the favored one (by more than 5 kcal mol−1) and can be considered rather as rare forms. The change of isomeric preferences shows that prototropy occurs in mono-anionic C. After deprotonation of the canonical isomer C-18 at N1H to the isomer ma-C-8, the labile proton at N8H can move to N3 and the isomer ma-C-3b of the lowest Gibbs energy can be formed. Note that Gibbs energy of its geometrical isomer ma-C-3a is close to that of the favored one (ΔG 1.8 kcal mol−1), indicating that the two geometrical isomers are the major forms in the isomeric mixture of mono-anionic cytosine. The major isomers (ma-C-3a and ma-C-3b) can also be formed from the rare neutral isomer C-38 in deprotonation reaction at N8H, as well as from the minor isomers C-13a or C-13b by deprotonation at N1H.
In the literature, usually, the N1 atom has been proposed as the favored site of cytosine deprotonation in the canonical neutral tautomer
C-18, and the isomer
ma-C-8 considered as the favored form in the gas phase [
61]. Prototropy in the deprotonated form has not been taken into account. Hence, this is not in agreement with our DFT investigations on the complete isomeric mixture of mono-deprotonated cytosine for which
ma-C-3 tautomer is favored in the gas phase. On the other hand, some mono-anionic isomers of cytosine (
ma-C-8,
ma-C-1b,
ma-C-3a, and
ma-C-3b) have been selected for interaction with the metal-cation Ba
2+ and investigated by both theory (DFT) and experiment (IRMPD) [
62]. The lowest Gibbs energy in the gas phase has been found for the
ma-C-8−Ba
2+ adduct with metal-cation chelated by N1 and O7 atoms. However, comparison of the calculated and experimental spectra in the 1550-1630 cm
−1 spectral region indicated that the
ma-C-8−Ba
2+ adduct of higher Gibbs energy with metal-cation chelated by N3 and O7 atoms cannot be neglected in the isomeric mixture in the gas phase. This result is very interesting for metal-cation adducts, but not for proton-transfer chemistry. It is well recognized that metal-cation can change the isomeric preference in tautomeric system and the favored isomer in adduct can be different than that in isolated tautomeric system. Lack of data in the literature for mono-anionic tautomers–rotamers of cytosine in vacuo makes any comparison of our DFT results impossible.
A different situation takes place in the case of isocytosine. From our DFT calculations result, the two major mono-anionic isomers ma-iC-3a and ma-iC-7 (ΔG 0.4 kcal mol−1) can be formed from the two major neutral isomers iC-37 and iC-78a (or partially from iC-78b) in deprotonation at N7H and O8H, respectively. On the other hand, deprotonation of the major neutral isomer iC-37 at N7 leads to the rare mono-anionic isomer ma-iC-3b. This isomer can also be formed from the rare neutral isomers iC-13b and iC-37 in deprotonation at N1 and N7, respectively.
In our previous work on isocytosine-charged radicals [
63], we tested deprotonation reaction of three neutral isocytosine isomers (
iC-17,
iC-37, and
iC-78a). Considering seven among nine mono-anionic tautomers–rotamers studied here, we found analogous stability-order for them at the DFT level. However, G4 calculations performed for the two major mono-anionic isomers showed their reverse stabilities. The isomer
ma-iC-7 has slightly lower Gibbs energy than
ma-iC-3a at the G4 level. This indicates that the two isomers
ma-iC-3a and
ma-iC-7 participate in the isomeric mixture in similar amount. In an aqueous solution, only
ma-iC-7 seems to be favored, and
ma-iC-3a can be treated as a rare isomer. The solvation effect has been investigated by
1H,
13C, and
15N NMR experiments, and also by PCM calculations [
27,
63].
3.3. Isomerism in Protonated Bases
Owing to n-π conjugation in the tautomeric moieties (push-pull effect), the sp
2-hybridized heteroatoms (=N− and =O) are candidate protonation-sites in the neutral pyrimidine bases
C and
iC (
Figure 3). Mono-protonation of the potential sites in fourteen neutral isomers lead to the formation of nine mono-cationic isomers (
Figure 6). They contain three labile protons and the same number of conjugated tautomeric sites—five, such as their neutral forms. Isomers with one labile proton at C5 have been neglected in calculations. Additionally, isomers protonated at the sp
3-hybridized endo and exo amino N atoms, exo hydroxy O atoms, and also sp
2-hybridized endo C atoms, even in the favored neutral forms have also been omitted. Tureček and co-workers [
33] investigating mono-protonation at N8H
2, C5H, and C6H in
C-18 and
C-78 proved by quantum–chemical calculations performed at different levels of theory (B3LYP, MP2, B3-MP2, CCSD(T) with less or more extended basis sets) that the N-sp
3 and C-sp
2 atoms are the last ones for the proton gain. They exhibit very weak basicities. Their protonated structures are insignificant in the isomeric mixture of protonated cytosine. Analogous conclusion has been derived by Lee and co-workers [
32] on the basis of DFT calculations of basicity parameters for N-sp
3 and O-sp
3 in
C-78a, and
C-78b and N-sp
3 in
C-18,
C-13,
C-78a, and
C-78b.
The ΔG values calculated for selected nine mono-cationic isomers (mc-C and mc-iC) vary from 0 to 33 kcal mol−1 for cytosine and from 0 to 24 kcal mol−1 for isocytosine. This suggests that the position-change of exo groups in the isomeric C and iC influences the relative stability of mono-cationic isomers. However, the same number of mono-cationic isomers with ΔG lower than 10 kcal mol−1 can dictate acid-base properties of pyrimidine bases, four of mc-C (mc-C-138, mc-C-178a, mc-C-178b, and mc-C-378a) and four of mc-iC (mc-iC-137, mc-iC-178a, mc-iC-178b, and mc-iC-378b). The three isomers selected for mono-cationic cytosine, major mc-C-138, major mc-C-178b, and rare mc-C-178a can be formed from the favored canonical isomer C-18 in protonation reaction at N3, O7b, and O7a, respectively, whereas the fourth rare isomer mc-C-378a is possible when the major isomer C-78a or rare isomer C-38 are protonated at N3 or O7a, respectively. Analogously for isocytosine, the major isomer mc-iC-178a can be directly formed from the major neutral isomer iC-78a in the protonation reaction at N1. Mono-protonation of the canonical form iC-37 at N1 or O8b leads to the rare isomers mc-iC-137 and mc-iC-378b, respectively.
Note that Tureček and co-workers [
33], investigating theoretically the protonated forms of four candidate isomers of neutral cytosine (
C-13b,
C-18,
C-78a, and
C-78b), considered three potential mono-cationic isomers (
mc-C-138,
mc-C-178b, and
mc-C-378a) and examined their relative stabilities using various levels of theory (B3LYP, MP2, B3-MP2, and CCSD(T) with less or more extended basis sets). This choice of isomers was sufficient to select the two major isomers
mc-C-138 and
mc-C-178b for protonated cytosine with not very different absolute relative energies (Δ
E 0–1.6 kcal mol
−1), and to subsequently study their dissociations in the gas phase by both theory and experiment using different theoretical methods and various experimental techniques of mass spectrometry. Cytosine mono-cations were generated by chemical ionization and fast atom bombardment, and dissociations of mono-cations examined by tandem mass spectrometry. In a completely theoretical work by employing different levels of the MP2 and MP4 methods, Leszczynski and co-workers [
30] also examined stability of some mono-cations. They considered three neutral cytosine isomers: the canonical form (
C-18) and two other minor isomers (
C-13b and
C-78a), and products of their protonation at heteroatoms. For the isomeric mixture of protonated cytosine, they also proposed a coexistence of the two major isomers
mc-C-138 and
mc-C-178b (Δ
G 0.1–1.2 kcal mol
−1) with slight preference of
mc-C-178b. The close stability of the two major mono-cationic isomers
mc-C-138 and
mc-C-178b in the gas phase have been confirmed by Salapin and co-workers [
34], who used both theoretical (DFT) and experimental (IRMPD) methods. The isomers
mc-C-138 and
mc-C-178a have been identified in the 1000–2000 cm
−1 spectral region.
In the case of isocytosine, the relative stabilities of the three potential mono-cationic isomers
mc-iC-137,
mc-iC-178a, and
mc-iC-378b considered as protonated forms of selected three neutral isomers
iC-17,
iC-37, and
iC-78a have been previously studied in our laboratory at the DFT level [
7] and analogous stability-order has been reported as that found here. G4-calculations [
63], performed for the two isomers
mc-iC-137 and
mc-iC-178a, confirmed the DFT-calculated stability-order and showed without any doubt that
mc-iC-178a has the lowest Gibbs energy for protonated isocytosine in the gas phase. However, the mono-cationic isomer
mc-iC-137 predominates in aqueous solution as confirmed by
1H,
13C, and
15N NMR experiments and by PCM calculations [
27,
63].
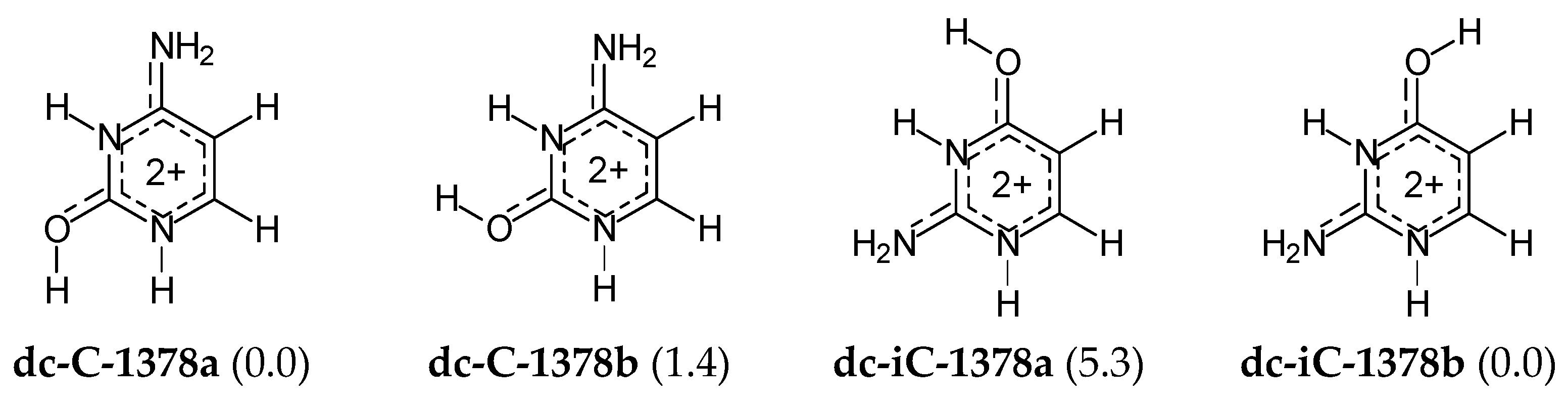
For di-protonated forms of cytosine and isocytosine, the isomeric mixtures are very simple, like for their di-deprotonated species. Neglecting the CH tautomers with Δ
G > 10 kcal mol
−1, only two geometrical isomers for each base (
Figure 7) have been considered in DFT-calculations for the isomeric mixtures. Their Gibbs energies do not differ very much, and thus both isomers contribute significantly to acid-base properties of the isomeric mixture of di-cationic cytosine (
dc-C) and di-cationic isocytosine (
dc-iC). There is no literature report for di-cationic forms of cytosine and isocytosine in the gas phase so no comparison can be made.
3.4. Microscopic Acidity–Basicity Parameters
Gas-phase acidity–basicity parameters, PA and/or GB defined as the enthalpy and/or Gibbs energy changes (Equations (2) and (3), respectively) for reversible protonation–deprotonation reactions (given in
Figure 2) can be analyzed from two view-points, i.e., on the microscopic (kinetic) and/or macroscopic (thermodynamic) scales. The microscopic acidity–basicity refers to equilibrium between the acid and base forms of selected isomer and helps to discuss the favored protonation and deprotonation sites in individual isomer. When tautomer–rotamer exists at sufficient long time to be isolated and analyzed in the gas phase, the microscopic acidity–basicity parameters can be even measured for the favored site(s) [
32]. For this reason, the microscopic PA and GB are called kinetic parameters.
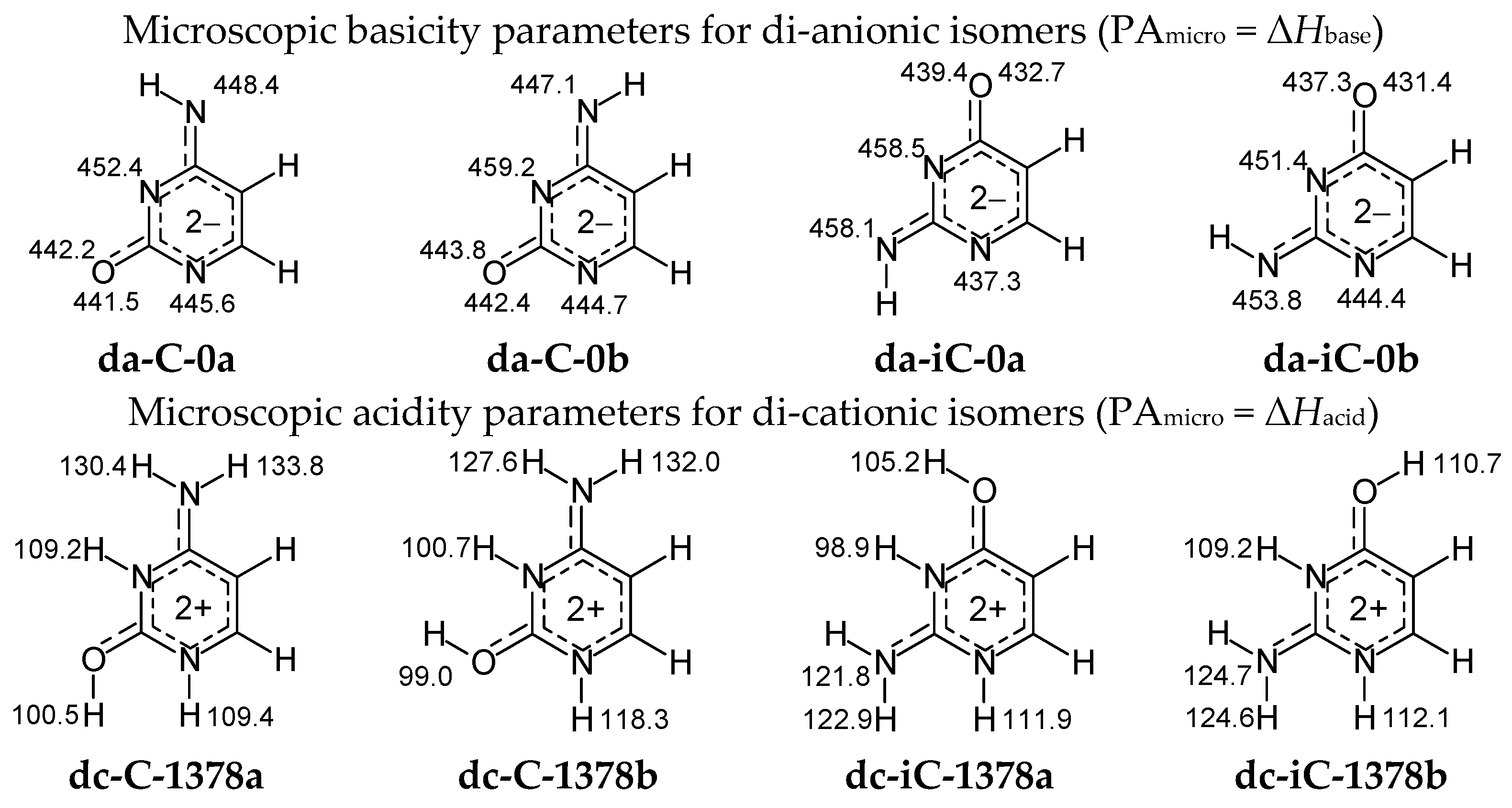
Figure 8 summarizes the microscopic acidity–basicity parameters in the enthalpy scale (PA = Δ
Hbase = Δ
Hacid) for protonation–deprotonation reactions estimated here according to Equation (2) at the DFT level for di-anionic and di-cationic isomeric forms of cytosine and isocytosine. Calculations have been carried out for each potential sites of protonation and deprotonation reaction, respectively, in the selected isomers of
da-C,
da-iC,
dc-C, and
dc-iC.
The highest basicity-parameter value for the di-anionic forms indicates the strongest basicity and the favored site of protonation: N3 in both isomers of da-C, whereas N3 in a and N7 in b of da-iC. Note that PAs of N3 and N7 in da-iC do not differ very much. The order of basicity parameters for heteroatoms in di-anionic cytosine and isocytosine isomers corresponds to the stability-order of the corresponding mono-anionic forms. On the other hand, the lowest acidity-parameter value for the di-cationic forms shows the strongest acidity as well as the preferred site of deprotonation: O7H in both isomers of dc-C, and N3H in both isomers of dc-iC. Note that acidities of N3H in dc-C-1378b and O8H in dc-iC-1378b are very close to those of O7H and N3H, respectively. The order of acidity parameters for NH and OH in di-cationic cytosine and isocytosine isomers refers to the stability-order of their mono-cationic isomers. In the literature, the is no document on gas-phase acidity–basicity parameters of di-anionic and di-cationic forms of cytosine and isocytosine and no comparison can be made.
For neutral
C and
iC isomers, microscopic basicity-parameters have been estimated for the sp
2-hybridized heteroatoms =N− and =O, and acidity-parameters for–NH− and –OH with the sp
3-hybridized heteroatoms. The DFT-calculated parameters in the enthalpy scale (PA = Δ
Hbase = Δ
Hacid) are included in
Figure 9. Variations of their values clearly show a strong relation between acidity–basicity and intramolecular proton-transfer (prototropy). Generally, =N− seems to be more basic (higher PA) than =O for cytosine isomers. Some exception is the favored canonical form
C-18, for which the O7b-carbonyl possesses basicity close to that of the imino N3. Analogous trend takes place for isocytosine with one exception of the rare isomer
iC-17, for which =O8b seems to be more basic than =N3−. In deprotonation reaction, –OH displays higher acidity (lower PA) than −NH− for both
C and
iC. When –OH is absent in isomer, −NH− can be deprotonated, but the favored site depend on the structure of isomer.
The canonical forms play a very significant role in the isomeric mixtures of C and iC. In the cytosine isomer C-18, =N3– (or =O7b) seems to be the favored site of protonation, and –N1H– is preferentially deprotonated, whereas =N1– favorably gains a proton, and –N7bH– (or –N3H–) loses its labile proton in the isocytosine isomer iC-37. In the favored isomer iC-78a, =N1– is also more basic than =N3–, whereas –O8aH is more acidic then –N7H2. The more basic and more acidic sites participate preferentially in protonation and deprotonation reactions, respectively, for isolated isomers.
In the literature, one can find theoretical microscopic basicity parameters for the canonical forms and for some other isomers of pyrimidine bases [
7,
28,
29,
30,
31,
32,
33,
63]. For example, Tureček and co-workers [
33] performing calculations at different levels of theory (B3LYP, MP2, B3-MP2, CCSD(T) with various basis sets) for mono-protonated cytosine showed that the microscopic PA values of the two potential protonation sites (N3 and O7b) in the canonical isomer
C-18 strongly depend on the level of calculations. They are in the following ranges: 225–235 kcal mol
−1 for N3 and 226–235 kcal mol
−1 for O7b. However, at each level of theory, the difference between PAs of N3 and O7b in
C-18 is very small (ca. 0–1 kcal mol
−1) so that both sites can chelate the proton. Analogous PAs at the B3LYP/6-31+G(d) level (226.5 and 225.5 kcal mol
−1 for N3 and O7b in
C-18, respectively) and absolute PA difference (1 kca mol
−1) have been reported by Lee and co-workers [
32], and also earlier by Leszczynski and co-workers [
30], who applied various MP2 and MP4 levels of theory to calculate PAs for N3 and O7b in
C-18 (227.2–230.8 and 228.3–230.6 kcal mol
−1, respectively). Investigating protonation of C5H, C6H and N7H
2 groups in
C-18, Tureček and co-workers [
33] confirmed additionally by calculations of their PAs that these sites display exceptionally weak basicity at each level of applied theory. Similarly, Lee and co-workers [
32], calculating PAs for selected sp
2- and sp
3-hybridized N and O in four cytosine isomers (
C-18,
C-13,
C-78a, and
C-78b) at the DFT level, confirmed higher PA values (stronger basicity) for =N– and =O than for >N– and –O–. In our previous work on prototropy in neutral isocytosine [
7], we reported the DFT-calculated microscopic PAs for N1, N3, and O8 sites in three selected isomers of neutral cytosine (
ic-17,
iC- 37, and
ic-78a). The PAs order is the same as that in
Figure 9.
In the case of mono-deprotonation reaction of neutral cytosine, Lee and co-workers [
32] calculated the microscopic acidity-parameters for the candidate sites in four major and minor neutral cytosine isomers (
C-18,
C-13,
C-78a, and
C-78b) at the B3LYP/6-31+G(d) level. For the canonical isomer
C-18, the order of acidity-parameters (in enthalpy scale) is analogous to that studied here, and their values are as follows: 343.3 (N1H), 346.6 (N8bH) and 352.6 (N8aH) kcal mol
−1, indicating also the highest acidity of the endo N1H group. For other isomers, analogous microscopic acidity–basicity parameters have been reported as those found here. The level of theory applied by Lee and co-workers give the values only slightly lower that that employed in this work.
For isocytosine, our earlier calculations performed at the G4 level [
63] for favored isomer predict almost the same microscopic acidity: 340.6 kcal mol
−1 for O8H in
iC-78a as that at the DFT level (339.9 kcal mol
−1). Difference in the G4- and DFT-calculated PA values is not larger than 1 kcal mol
−1. An exception is the major isomer
iC-37, for which N3H seems to lose the proton first than N7bH at the G4 level. However, differences in the microscopic acidities (338.4 and 339.6 kcal mol
−1 at the G4 level, whereas 339.1 and 338.7 kcal mol
−1 at the DFT level, respectively) are not very large (ca. 1 kcal mol
−1). This suggests that the two sites in
iC-37 can lose the proton with similar probability. It would be interesting to confirm this theoretical prediction in the future by appropriate experiment(s) performed for mono-anionic isocytosine in the gas phase.
3.5. Acid-Base Equilibria for Pyrimidine Bases
For simple bifunctional tautomeric compounds of the following general formula: HA−X=B ⇄ A=X−BH, containing only one labile proton that move between only two conjugated sites (A and B), like in acetaldehyde, formamide, formamidine, etc., acid-base equilibria are very simple. Protonation and deprotonation reactions, analogous to those for amphiprotic compounds, lead to one cation and one anion, respectively, common for the two prototropic tautomers. Only π-electrons and charge in ions are better delocalized than in neutral tautomers. According to Brønsted and Lowry theory, acid-base equilibria for simple bifunctional tautomeric compounds can be written as follows [
2,
4,
64]: (AXB)
− + 2H
+ ⇄ (HA−X=B ⇄ A=X−BH) + H
+ ⇄ (HAXBH)
+. These equilibria have frequently been considered in explanation of the intramolecular proton-transfer (prototropy) mechanism [
4].
However, for polyfunctional tautomeric systems, like pyrimidine bases, the situation is more complex. Containing two labile protons and five conjugated sites, neutral bases exist in the mixture of twenty-one tautomers–rotamers that can be protonated or deprotonated. Possibility of formation of different cations and anions in proton-transfer reactions explains the existence of various ionic isomers for the tautomeric molecules. Consequently, acid-base equilibria, generally written for pyrimidine bases in
Figure 2, are more complex than for bifunctional derivatives. The abbreviations AH
2B, (AB)
2−, (AHB)
−, (AH
2BH)
+, and (AH
2BH
2)
2+ in
Figure 2 refer to the corresponding tautomeric mixtures for neutral
C or
iC and for their ionic forms, respectively.
Figure 10 shows four-steps for acid-base equilibria of tautomeric cytosine and isocytosine: two steps of deprotonation and two steps of protonation. For simplicity, geometrical isomerism of the exo =NH group about the C=N double bond and rotational isomerism of the exo −OH group about the C−O single bond are not displayed. Exceptionally rare isomers are omitted in the isomeric mixtures of both the neutral and ionic forms. Only the structures, selected tautomers for DFT calculations, are included in this scheme, i.e., one structure for (AB)
2−, four tautomers for (AHB)
−, six tautomers for AH
2B, four tautomers for (AH
2BH)
+, and one structure for (AH
2BH
2)
2+.
3.6. Macroscopic Acidity–Basicity Parameters
The macroscopic acidity–basicity parameters correspond to deprotonation-protonation equilibria between the isomeric mixtures of the acid and base forms. They are determined mainly by the major and minor isomers. The rare forms have usually small effect on their values. When all isomeric conversions in the acid and base forms are in equilibrium with intermolecular proton-transfer during acidity–basicity measurements in the gas phase, the experimental acidity–basicity parameters for tautomeric systems refer to the isomeric mixtures of the acid and base forms, and may be compared with theoretically estimated macroscopic acidity–basicity parameters. For this reason, the macroscopic PA and GB for tautomeric systems are named thermodynamic parameters.
It has been shown above (
Figure 3) that at least six tautomers–rotamers for neutral cytosine (
C-13a,
C-13b,
C-18,
C-38,
C-78a, and
C-78b) and at least six tautomers–rotamers for neutral isocytosine (
iC-13a,
iC-13b,
iC-17,
iC-37,
iC-78a, and
iC-78b) should contribute to acid-base properties of
C and
iC. For anionic bases, the following isomers are the most important: two geometrical isomers of di-deprotonated
C (
da-C-a and
da-C-b) and
iC (
da-iC-a and
da-iC-b) that exist in the di-anionic isomeric mixtures (
Figure 4), and at least four isomers of mono-deprotonared
C (
ma-C-1a,
ma-C-3a,
ma-C-3b, and
ma-C-8) and three isomers of mono-deprotonared
iC (
ma-iC-3a,
ma-iC-3b, and
ma-iC-7) that should be considered in the isomeric mixture of mono-anionic bases (
Figure 5). The isomeric mixtures of mono-protonated cytosine (
Figure 6) consist at least of four isomers (
mc-C-138,
mc-C-178a,
mc-C-178b, and
mc-C-378a). The same is true for mono-protonated isocytosine, four isomers can be considered (
mc-iC-137,
mc-iC-178a,
mc-iC-178b, and
mc-iC-378b). For di-protonated bases (
Figure 7), two geometrical isomers should be taken in the isomeric mixtures of cytosine (
da-C-a and
da-C-b) and isocytosine (
da-iC-a and
da-iC-b). Selection of the potential neutral and ionic isomers has been based on the Mezey et al. rule [
37,
38].
The abbreviations of selected neutral and ionic isomers for cytosine and isocytosine and their DFT-calculated percentage contents (
xi) estimated according to Equation (1) are given in
Table 2. The exact structures of chosen isomers in this table are the same as those in
Figure 4,
Figure 5,
Figure 6 and
Figure 7. Differences in acid-base properties of functional groups and intramolecular interactions in tautomeric species dictate
xi of individual isomers. Because functional groups in tautomeric (conjugated) systems upon protonation–deprotonation can change their acidity–basicity, and consequently intramolecular interactions between them can also change, the composition of tautomeric mixtures of neutral and ionic forms and contributions of potential isomers (
xi) also vary. These variations can be clearly seen regarding the
xi data in
Table 2.
For di-anionic bases, the isomers
da-C-0a and
da-iC-0b dominate in the isomeric mixtures. Although exo groups have different positions, the favored di-anionic isomers seem to be stabilized by an analogous intramolecular interaction between exo NH and endo N3. Note that N3 possesses higher basicity than N1 in di-deprotonated forms (
Figure 8).
The mono-anionic bases prefer the structures of the isomers ma-C-3b and ma-iC-3a. In this case, the two isomers seem to be preferentially stabilized by an intramolecular interaction between exo N and endo N3H. Additionally, intramolecular interactions between exo NH2 and both endo N1 and N3 seem to stabilize the isomer ma-iC-7 that significantly participates in the isomeric mixture of ma-iC as an additional major isomer. So favorable interactions for the ma-C-8 isomer are not possible due to repulsion between exo NH2 and endo C5H, which increases the Gibbs energy of this isomer and makes it less significant (rare form) in the mixture of ma-C isomers.
Mono-protonation of cytosine and isocytosine leads to the different tautomeric preferences, mc-C-138 and mc-iC-178a, respectively. The mc-iC-137 isomer—protonated form of the canonical tautomer iC-37—is rather a rare isomer. This difference in the favored tautomers can be a consequence of different internal effects. The protonated canonical isomer of cytosine (mc-C-138) is stabilized by the favorable intramolecular interactions between exo O and two acidic endo N1H and N3H. However, the protonated canonical isomer of isocytosine (mc-iC-137) is stabilized by the interaction between exo O and endo N3H and C5H (less acidic group). On the other hand, mc-C-178b—a second major tautomer in the isomeric mixture of protonated cytosine—is stabilized by an analogous intramolecular interaction between exo OH and endo N3 like the favored mc-iC-178a. The preferred isomers of di-cationic cytosine and isocytosine, dc-C-1378a and dc-iC-1378b are stabilized by analogous intramolecular interactions between exo O and endo N3H.
Taking into account the DFT-estimated percentage contents (
xi) and thermochemical parameters (
Hi and G
i) for all selected isomers, the macroscopic acidity–basicity parameters (PA
macro and GB
macro) have been estimated for the isomeric mixtures using Equations (4) and (5). Their values, summarized in
Table 3, indicate some differences in thermodynamic acid-base properties of the two pyrimidine bases. However, the absolute differences between macroscopic PAs (or GBs) are not larger than 6 kcal mol
−1.
In the literature, there is no article on theoretical estimation of the macroscopic (thermodynamic) acidity–basicity parameters for the complete isomeric mixtures of neutral and ionic cytosine. There are only some microscopic parameters estimated for particular acid-base sites in the canonical form of neutral cytosine and in its potential isomers (vide infra) [
28,
29,
30,
31,
32,
33]. Some exception is an estimation of Leszczynski and co-workers [
30], who considering only three neutral isomers (
C-18,
C-13b, and
C-78a) and two mono-cationic forms (
mc-C-138 and
mc-C-178b), proposed the average PA for cytosine (225.3–230.8 kcal mol
−1, calculated at various MP2 and MP4 levels). Concerning the experimental literature-data, one can find acidity–basicity parameters only for neutral cytosine, measured in different laboratories by various kinetic and equilibrium methods. For mono-deprotonation reaction of neutral base AH
2B ⇄ (AXB)
− + H
+, proton acidity parameters have been measured by Lee and co-workers [
32] using two procedures, bracketing and extended Cooks kinetic methods, and the following PA values have been reported 342 ± 3 and 343 ± 3 kcal mol
−1, respectively. The GB value of 335 ± 3 kcal mol
−1 has been found by the bracketing method. Analogous PA (340 kcal mol
−1) has been earlier derived for cytosine by Chen et al. [
66] employing electron impact spectra and acid dissociation constants determined in DMSO. For mono-protonation reaction of neutral cytosine AH
2B + H
+ ⇄ (HAXBH)
+, Hunter and Lias [
65] in 1998 compiled the PAs and GBs data, earlier reported by other chemists, and proposed for cytosine the following values 227.0 and 219.4 kcal mol
−1, respectively. Ten year later Lee and co-workers [
32] investigated proton basicity of neutral cytosine and found PA 228 ± 3 and GB 220 ± 3 kcal mol
−1 by the bracketing method, and PA 227 ± 3 kcal mol
−1 by the extended Cooks kinetic method. The calculated here macroscopic acidity–basicity parameters for neutral cytosine are close to those experimentally determined.
To our knowledge, the literature experimental gas-phase acidity–basicity parameters for isocytosine have not yet been documented [
55]. Preliminary prediction of proton acidity and proton basicity at the G4 level for neutral isocytosine considering only the favored isomers leads to the following PA values: 339.2 and 223.2 kcal mol
−1 in the enthalpy scale for (AHB)
− + H
+ ⇄ AH
2B and AH
2B + H
+ ⇄ (AH
2BH)
+ reactions, respectively [
63]. They are close to those estimated in this work at the DFT level. It should also be mentioned here that the experimental p
Ka values for the two steps of acid-base equilibria in aqueous solution (9.6 and 4.0 [
27]) correspond also to the macroscopic p
Kas.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}