
Asymmetry of Endocast Surface Shape in Modern Humans Based on Diffeomorphic Surface Matching


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
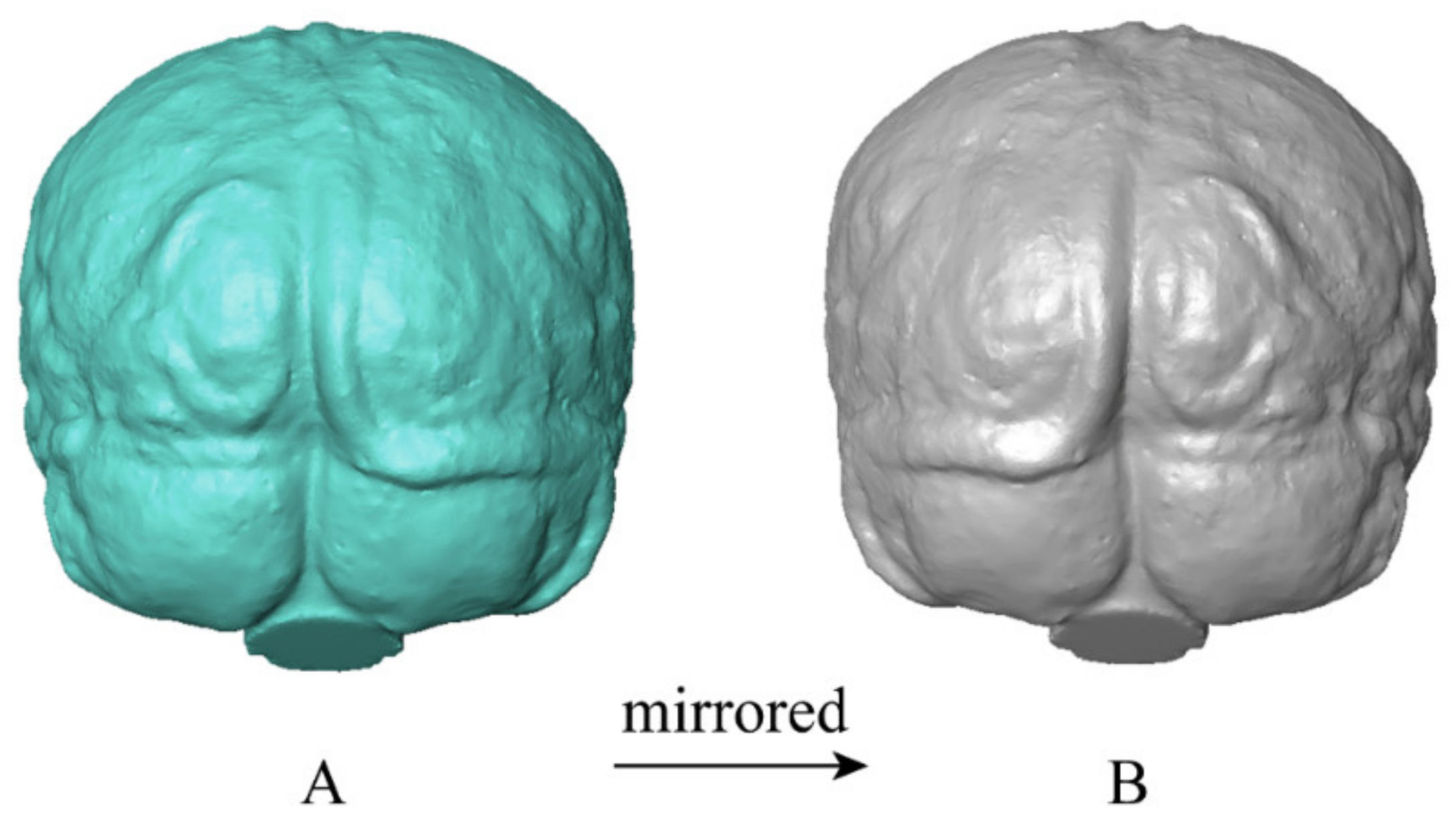
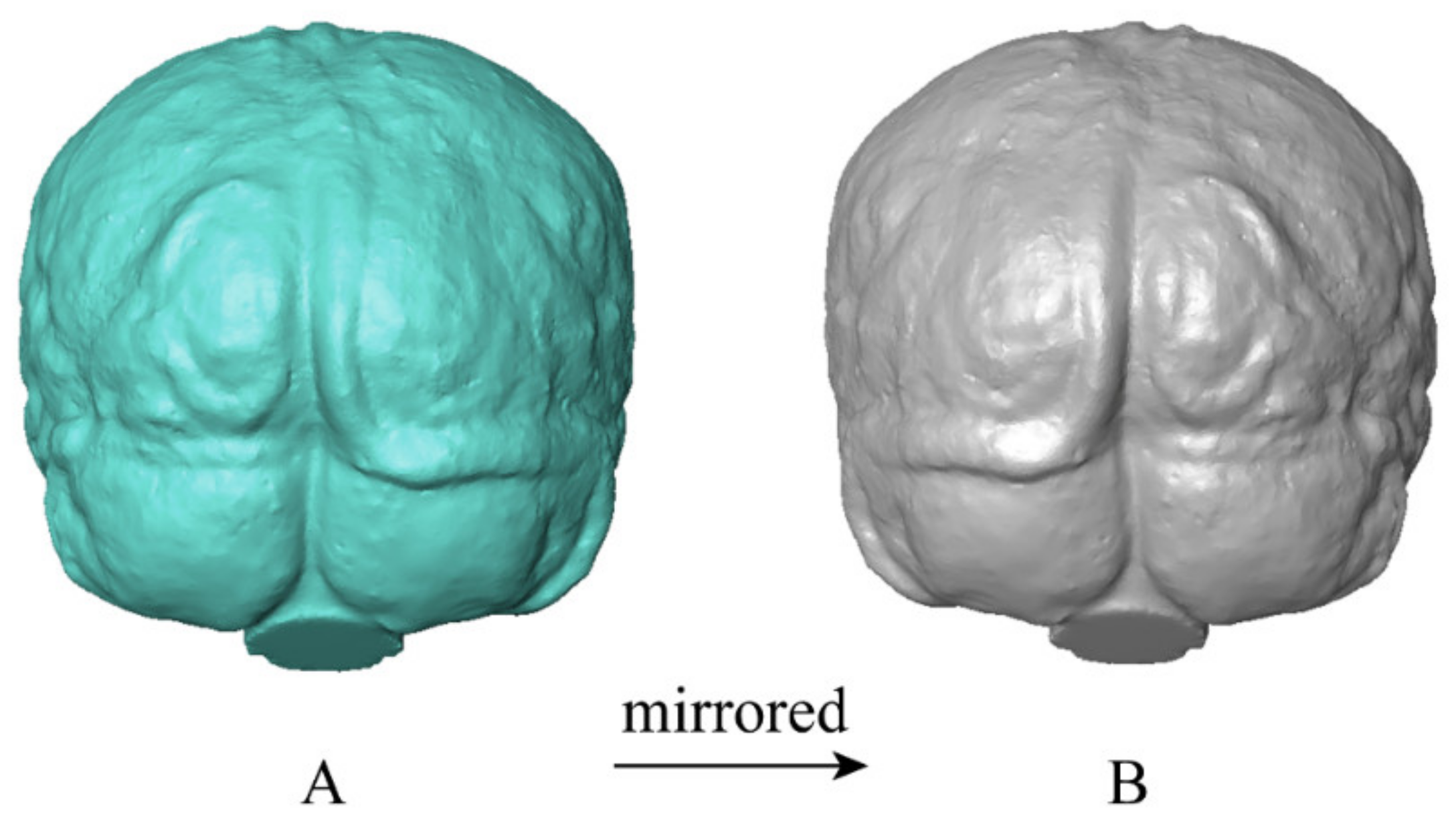
2.2. Endocast Reconstruction and Processing
2.3. Diffeomorphic Surface Asymmetry
3. Results
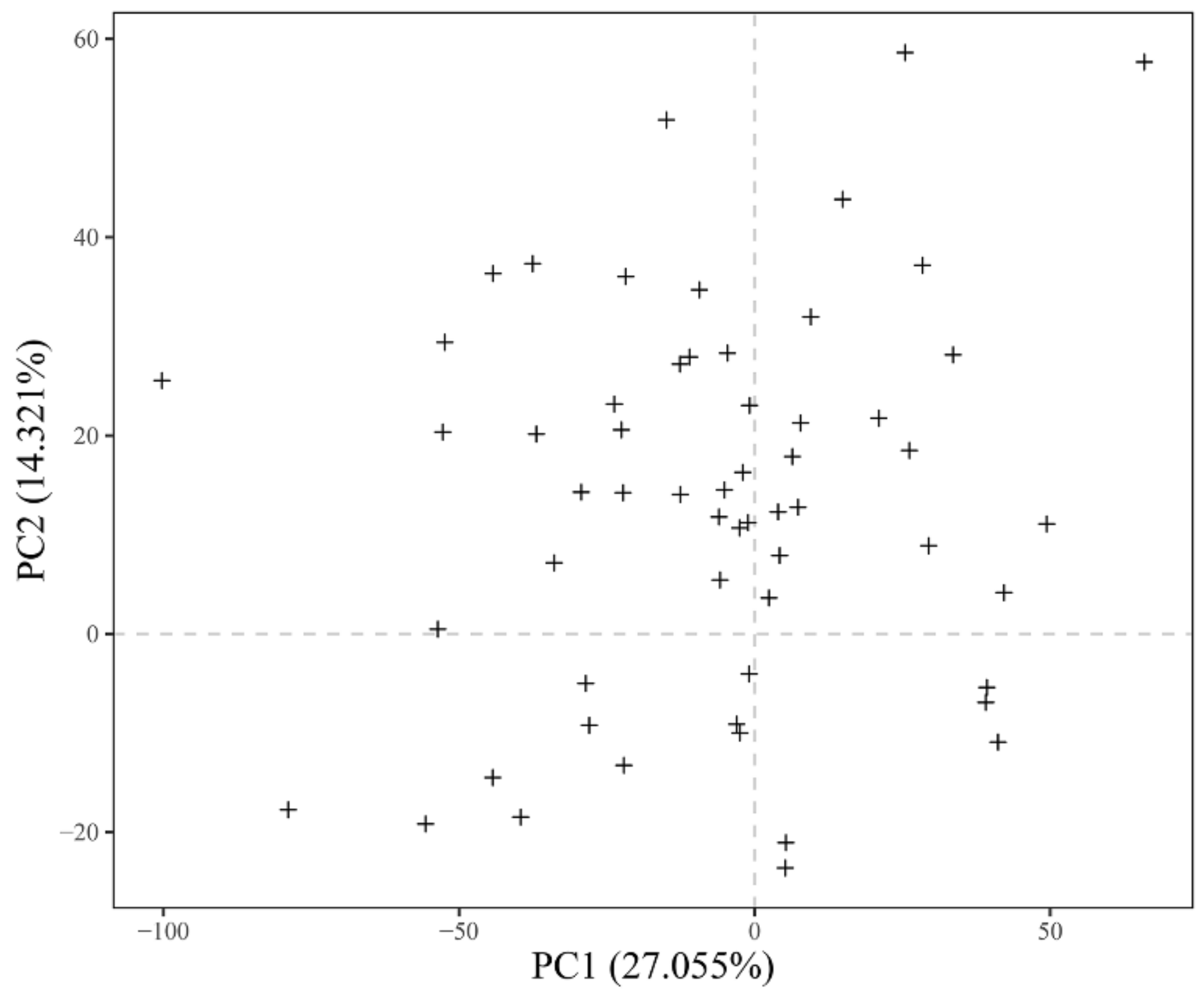
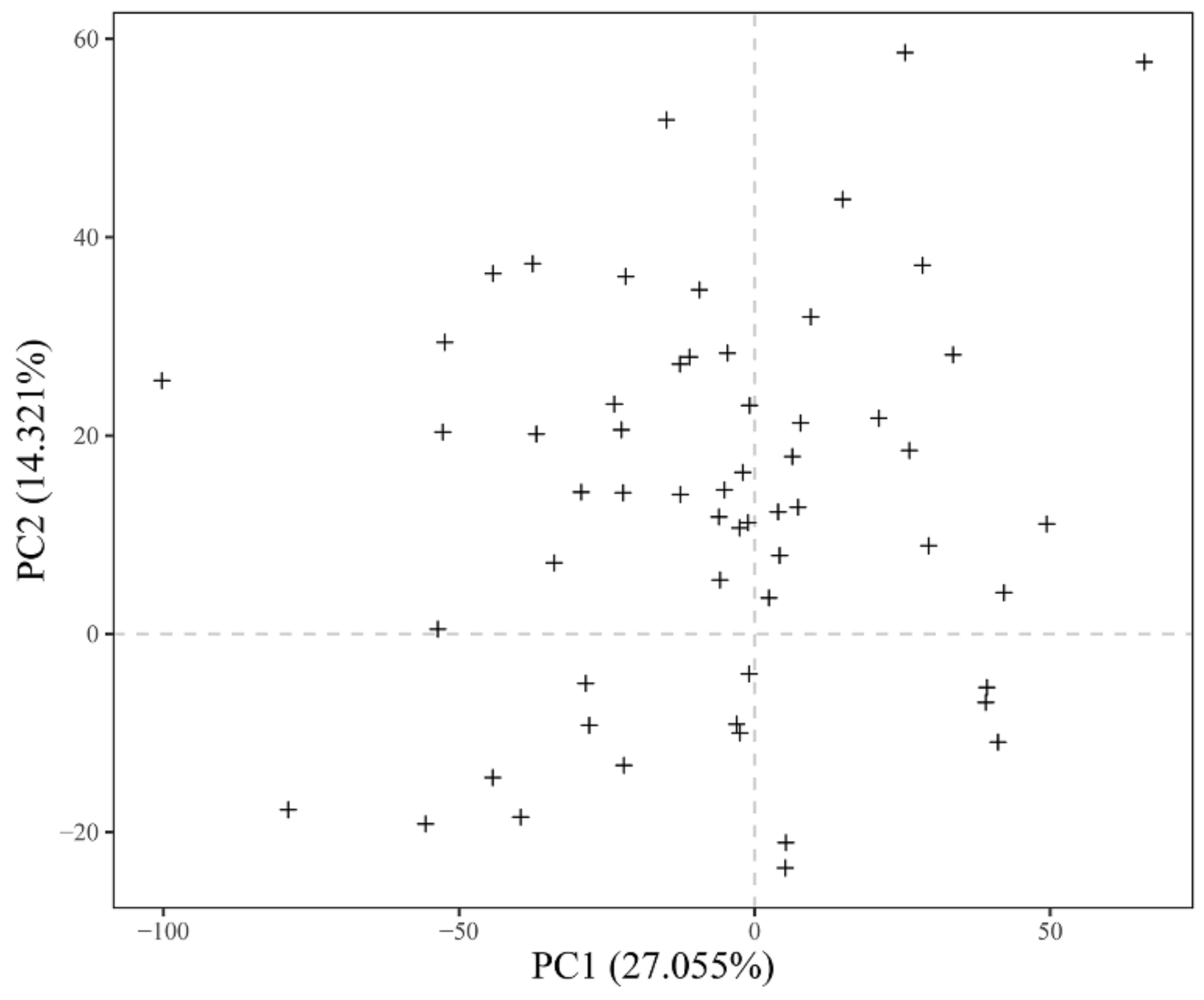
3.1. Principal Component Analysis
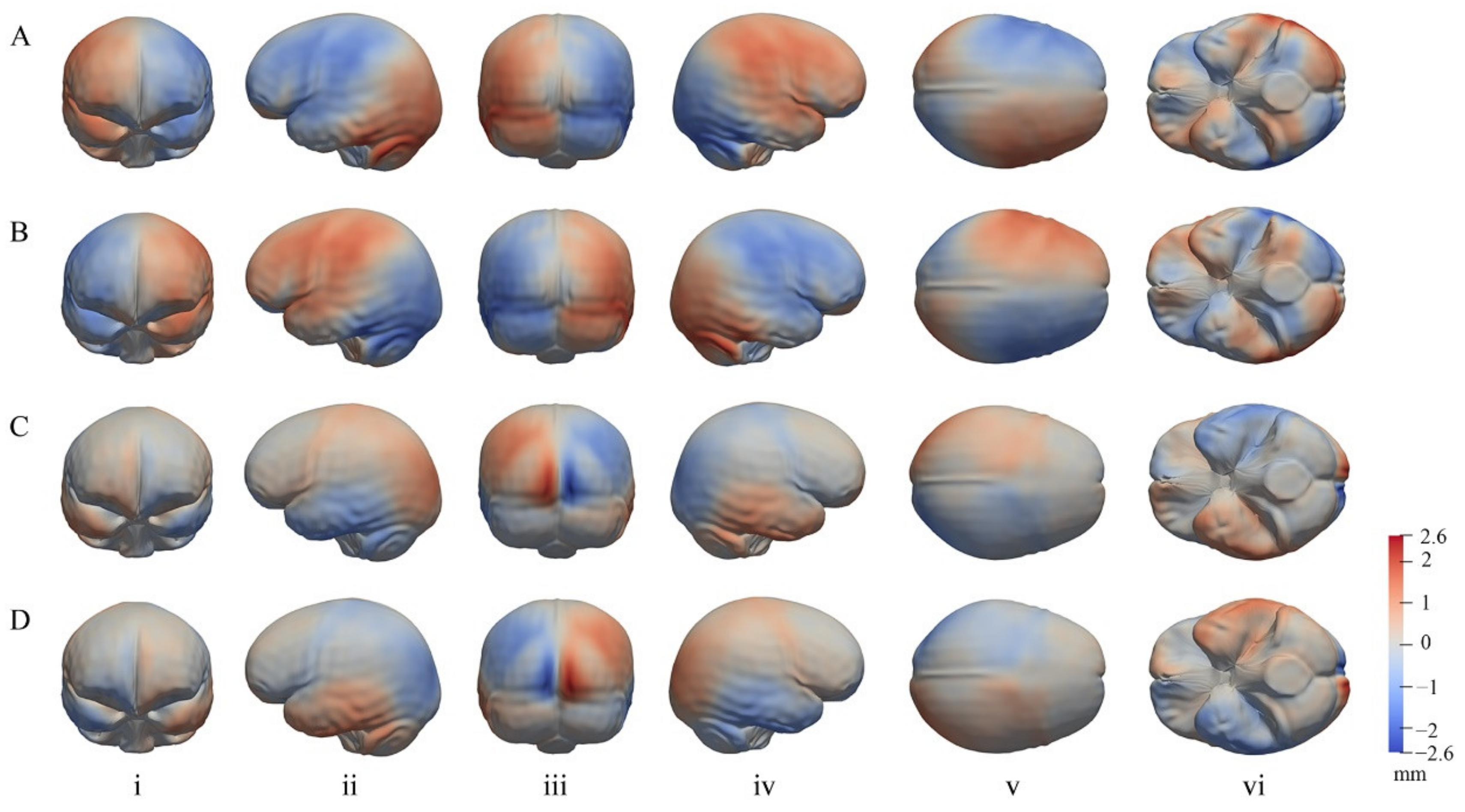
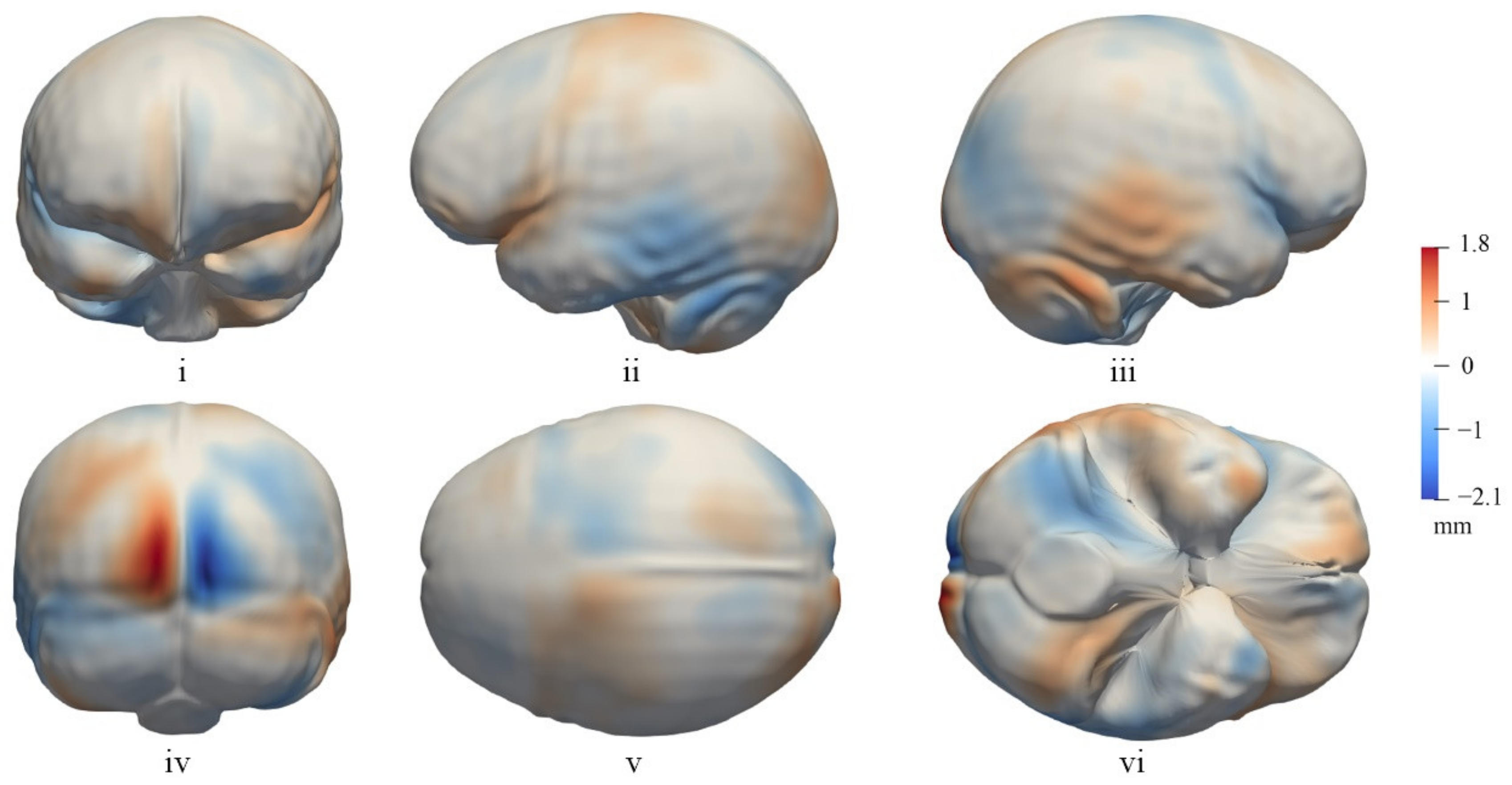
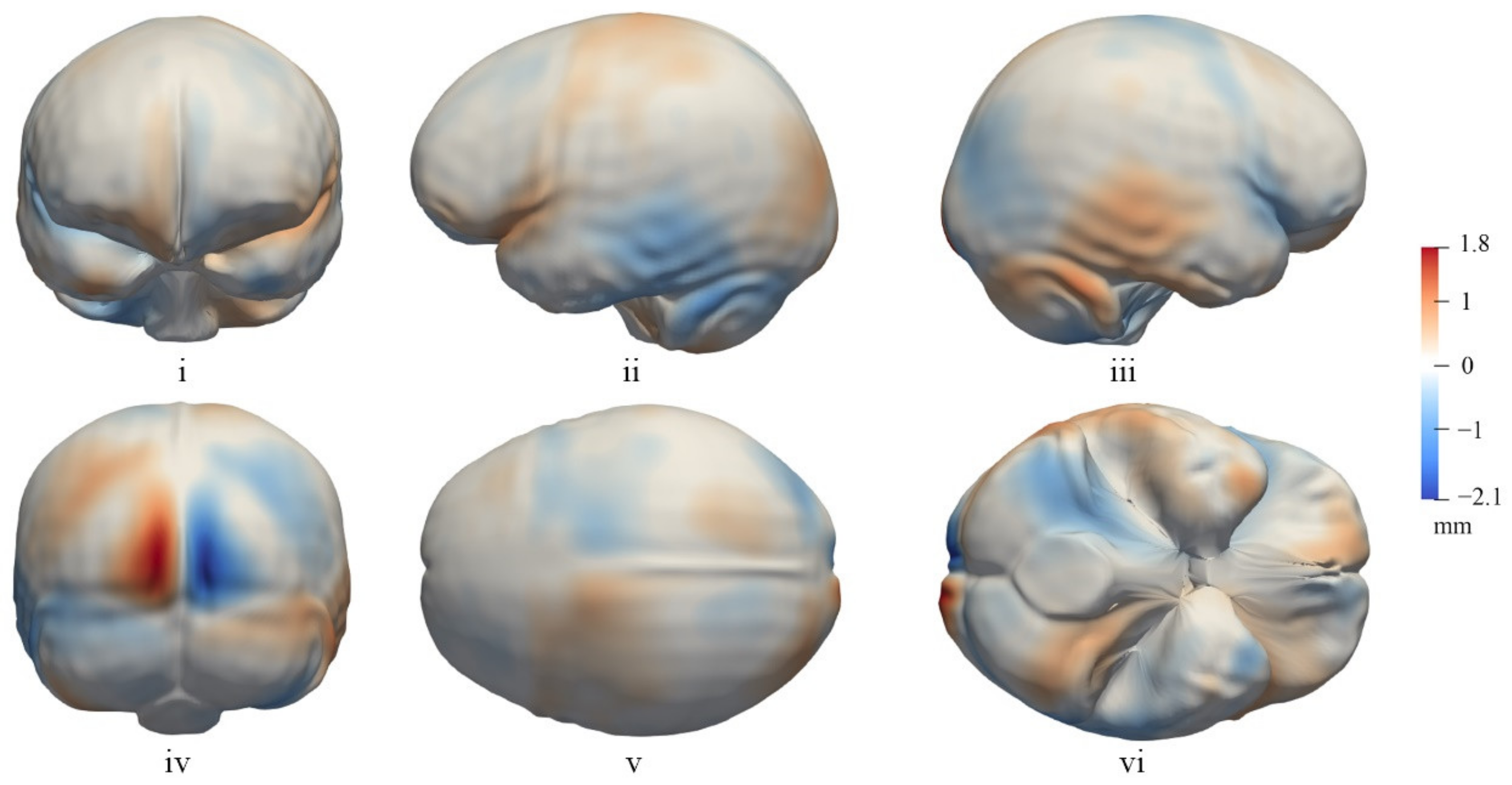
3.2. The General Pattern of Endocast Surface Asymmetry
4. Discussion
4.1. Global Endocast Asymmetry
4.2. Local Asymmetries of the Cerebrum
4.3. Asymmetry of the Cerebellum
5. Significance and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zilles, K.; Dabringhaus, A.; Geyer, S.; Amunts, K.; Qü, M.; Schleicher, A.; Gilissen, E.; Schlaug, G.; Steinmetz, H. Structural asymmetries in the human forebrain and the forebrain of non-human primates and rats. Neurosci. Biobehav. Rev. 1996, 20, 593–605. [Google Scholar] [CrossRef]
- Bear, D.; Schiff, D.; Saver, J.; Greenberg, M.; Freeman, R. Quantitative analysis of cerebral asymmetries: Fronto-occipital correlation, sexual dimorphism and association with handedness. Arch. Neurol. 1986, 43, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Balzeau, A.; Gilissen, E.; Grimaud-Hervé, D. Shared pattern of endocranial shape asymmetries among great apes, anatomically modern humans, and fossil hominins. PLoS ONE 2012, 7, e29581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzeau, A.; Holloway, R.L.; Grimaud-Hervé, D. Variations and asymmetries in regional brain surface in the genus Homo. J. Hum. Evol. 2012, 62, 696–706. [Google Scholar] [CrossRef]
- Kitchell, L.M.; Schoenemann, P.T.; Loyet, M. Structural asymmetries in the human brain assessed via MRI. Am. J. Phys. Anthropol. 2013, 167. [Google Scholar]
- Wang, D.; Buckner, R.L.; Liu, H. Cerebellar asymmetry and its relation to cerebral asymmetry estimated by intrinsic functional connectivity. J. Neurophysiol. 2012, 109, 46–57. [Google Scholar] [CrossRef]
- LeMay, M. Morphological cerebral asymmetries of modern man, fossil man, and nonhuman primate. Ann. N. Y. Acad. Sci. 1976, 280, 349–366. [Google Scholar] [CrossRef]
- Kuo, F.; Massoud, T.F. Structural asymmetries in normal brain anatomy: A brief overview. Ann. Anat.-Anat. Anz. 2022, 241, 151894. [Google Scholar] [CrossRef]
- Sha, Z.; Pepe, A.; Schijven, D.; Carrión-Castillo, A.; Roe, J.M.; Westerhausen, R.; Joliot, M.; Fisher, S.E.; Crivello, F.; Francks, C. Handedness and its genetic influences are associated with structural asymmetries of the cerebral cortex in 31,864 individuals. Proc. Natl. Acad. Sci. USA 2021, 118, e2113095118. [Google Scholar] [CrossRef]
- Dumoncel, J.; Subsol, G.; Durrleman, S.; Bertrand, A.; de Jager, E.; Oettle, A.C.; Lockhat, Z.; Suleman, F.E.; Beaudet, A. Are endocasts reliable proxies for brains? A 3D quantitative comparison of the extant human brain and endocast. J. Anat. 2021, 238, 480–488. [Google Scholar] [CrossRef]
- Holloway, R.L.; Broadfield, D.C.; Yuan, M.S.; Schwartz, J.H.; Tattersall, I. The Human Fossil Record, Brain Endocasts: The Paleoneurological Evidence; John Wiley & Sons: Hoboken, NJ, USA, 2004; Volume 3. [Google Scholar]
- Holloway, R.L.; De La Costelareymondie, M.C. Brain endocast asymmetry in pongids and hominids: Some preliminary findings on the paleontology of cerebral dominance. Am. J. Phys. Anthropol. 1982, 58, 101–110. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; McIntyre, G.S. Geometric morphometrics of developmental instability: Analyzing patterns of fluctuating asymmetry with procrustes methods. Evolution 1998, 52, 1363–1375. [Google Scholar] [CrossRef]
- Neubauer, S.; Gunz, P.; Scott, N.A.; Hublin, J.-J.; Mitteroecker, P. Evolution of brain lateralization: A shared hominid pattern of endocranial asymmetry is much more variable in humans than in great apes. Sci. Adv. 2020, 6, eaax9935. [Google Scholar] [CrossRef] [Green Version]
- Gundara, N.; Zivanovic, S. Asymmetry in East African skulls. Am. J. Phys. Anthropol. 1968, 28, 331–337. [Google Scholar] [CrossRef]
- Hadžiselimović, H.; Čuš, M. The appearance of internal structures of the brain in relation to configuration of the human skull. Acta Anat. 1966, 63, 289–299. [Google Scholar] [CrossRef]
- Holloway, R.L. Volumetric and asymmetry determinations on recent hominid endocasts-Spy-I and Spy-II, Djebel lhroud-I, and the Sale Homo-erectus specimens, with some notes on Neandertal brain size. Am. J. Phys. Anthropol. 1981, 55, 385–393. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E. Endocranial shape asymmetries in Pan paniscus, Pan troglodytes and Gorilla gorilla assessed via skull based landmark analysis. J. Hum. Evol. 2010, 59, 54–69. [Google Scholar] [CrossRef]
- Geschwind, D.H.; Miller, B.L.; DeCarli, C.; Carmelli, D. Heritability of lobar brain volumes in twins supports genetic models of cerebral laterality and handedness. Proc. Natl. Acad. Sci. USA 2002, 99, 3176–3181. [Google Scholar] [CrossRef] [Green Version]
- Good, C.D.; Johnsrude, I.; Ashburner, J.; Henson, R.N.A.; Friston, K.J.; Frackowiak, R.S.J. Cerebral asymmetry and the effects of sex and handedness on brain structure: A voxel-based morphometric analysis of 465 normal adult human brains. NeuroImage 2001, 14, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Matloff, W.; Shi, Y.; Cabeen, R.P.; Toga, A.W. Mapping complex brain torque components and their genetic architecture and phenomic associations in 24,112 individuals. Biol. Psychiatry 2022, 91, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Marino, L. Asymmetries in cerebral width in nonhuman primate brains as revealed by magnetic resonance imaging (MRI). Neuropsychologia 2000, 38, 493–499. [Google Scholar] [CrossRef]
- Toga, A.W.; Thompson, P.M. Mapping brain asymmetry. Nat. Rev. Neurosci. 2003, 4, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Kitchell, L.M. Asymmetry of the Modern Human Endocranium. Ph.D. Thesis, Institute of Archaeology, University College London, London, UK, 2015. [Google Scholar]
- Broca, P. Remarques sur le siège de la faculté du langage articulé, suivies d’une observation d’aphémie (perte de la parole). Bull. Mem. Soc. Anat. Paris 1861, 6, 330–357. [Google Scholar]
- Shaywitz, B.A.; Shaywltz, S.E.; Pugh, K.R.; Constable, R.T.; Skudlarski, P.; Fulbright, R.K.; Bronen, R.A.; Fletcher, J.M.; Shankweiler, D.P.; Katz, L.; et al. Sex differences in the functional organization of the brain for language. Nature 1995, 373, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Kitchell, L.M. Evaluating Language-Related Asymmetry in Endocasts Using Non-Rigid Diffeomorphic Image Registration. 2017. Available online: https://www.researchgate.net/publication/314299269_Evaluating_Language-Related_Asymmetry_in_Endocasts_using_Non-rigid_Diffeomorphic_Image_Registration (accessed on 5 October 2021).
- Tobias, P.V. The brain of Homo habilis: A new level of organization in cerebral evolution. J. Hum. Evol. 1987, 16, 741–761. [Google Scholar] [CrossRef]
- Holloway, R.L. Human paleontological evidence relevant to language behavior. Hum. Neurobiol. 1983, 2, 105–114. [Google Scholar]
- De Sousa, A.; Cunha, E. Hominins and the emergence of the modern human brain. Prog. Brain Res. 2012, 195, 293–322. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E.; Holloway, R.L.; Prima, S.; Grimaud-Hervé, D. Variations in size, shape and asymmetries of the third frontal convolution in hominids: Paleoneurological implications for hominin evolution and the origin of language. J. Hum. Evol. 2014, 76, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Barrick, T.R.; Mackay, C.E.; Prima, S.; Maes, F.; Vandermeulen, D.; Crow, T.J.; Roberts, N. Automatic analysis of cerebral asymmetry: An exploratory study of the relationship between brain torque and planum temporale asymmetry. NeuroImage 2005, 24, 678–691. [Google Scholar] [CrossRef]
- Foundas, A.L.; Leonard, C.M.; Hanna-Pladdy, B. Variability in the anatomy of the planum temporale and posterior ascending ramus: Do right- and left handers differ? Brain Lang. 2002, 83, 403–424. [Google Scholar] [CrossRef]
- Watkins, K.E.; Paus, T.; Lerch, J.P.; Zijdenbos, A.; Collins, D.L.; Neelin, P.; Taylor, J.; Worsley, K.J.; Evans, A.C. Structural asymmetries in the human brain: A voxel-based statistical analysis of 142 MRI scans. Cereb. Cortex 2001, 11, 868–877. [Google Scholar] [CrossRef]
- Geschwind, N.; Levitsky, W. Human brain: Left-right asymmetries in temporal speech region. Science 1968, 161, 186–187. [Google Scholar] [CrossRef]
- Wada, J.A.; Clarke, R.; Hamm, A. Cerebral hemispheric asymmetry in humans: Cortical speech zones in 100 adult and 100 infant brains. Arch. Neurol. 1975, 32, 239–246. [Google Scholar] [CrossRef]
- Witelson, S.F.; Pallie, W. Left hemisphere specialization for language in the newborn: Neuroanatomical evidence of asymmetry. Brain 1973, 96, 641–646. [Google Scholar] [CrossRef]
- Wu, X.J.; Zhang, X. Progress in endocast and human brain evolution on Chinese human fossils. Acta Anthropol. Sin. 2018, 37, 371–383. (In Chinese) [Google Scholar]
- Melchionna, M.; Profico, A.; Castiglione, S.; Sansalone, G.; Serio, C.; Mondanaro, A.; Di Febbraro, M.; Rook, L.; Pandolfi, L.; Di Vincenzo, F.; et al. From smart apes to human brain boxes. A uniquely derived brain shape in late hominins clade. Front. Earth Sci. 2020, 8, 273. [Google Scholar] [CrossRef]
- Wu, X.J.; Schepartz, L.A.; Falk, D.; Liu, W. Endocranial cast of Hexian Homo erectus from South China. Am. J. Phys. Anthropol. 2006, 130, 445–454. [Google Scholar] [CrossRef]
- Wu, X.J.; Liu, W.; Dong, W.; Que, J.M.; Wang, Y.F. The brain morphology of Homo Liujiang cranium fossil by three-dimensional computed tomography. Sci. Bull. 2008, 53, 2513–2519. [Google Scholar] [CrossRef] [Green Version]
- Bruner, E.; Ripani, M. A quantitative and descriptive approach to morphological variation of the endocranial base in modern humans. Am. J. Phys. Anthropol. 2008, 137, 30–40. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Marcus, L.F. A revolution morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Beaudet, A.; Dumoncel, J.; de Beer, F.; Duployer, B.; Durrleman, S.; Gilissen, E.; Hoffman, J.; Tenailleau, C.; Thackeray, J.F.; Braga, J. Morphoarchitectural variation in South African fossil cercopithecoid endocasts. J. Hum. Evol. 2016, 101, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Beaudet, A.; Dumoncel, J.; de Beer, F.; Durrleman, S.; Gilissen, E.; Oettle, A.; Subsol, G.; Thackeray, J.F.; Braga, J. The endocranial shape of Australopithecus africanus: Surface analysis of the endocasts of Sts 5 and Sts 60. J. Anat. 2018, 232, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Durrleman, S.; Pennec, X.; Trouvé, A.; Ayache, N.; Braga, J. Comparison of the endocranial ontogenies between chimpanzees and bonobos via temporal regression and spatiotemporal registration. J. Hum. Evol. 2012, 62, 74–88. [Google Scholar] [CrossRef]
- Urciuoli, A.; Zanolli, C.; Beaudet, A.; Dumoncel, J.; Santos, F.; Moyà-Solà, S.; Alba, D.M. The evolution of the vestibular apparatus in apes and humans. eLife 2020, 9, e51261. [Google Scholar] [CrossRef]
- Braga, J.; Zimmer, V.; Dumoncel, J.; Samir, C.; de Beer, F.; Zanolli, C.; Pinto, D.; Rohlf, F.J.; Grine, F.E. Efficacy of diffeomorphic surface matching and 3D geometric morphometrics for taxonomic discrimination of Early Pleistocene hominin mandibular molars. J. Hum. Evol. 2019, 130, 21–35. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, X.J. 3D virtual reconstruction and morphological variations of the mastoid air cells—Take modern skulls from yunnan province for example. Quat. Sci. 2017, 37, 747–753. (In Chinese) [Google Scholar]
- Zhang, X.; Zhang, Y.M.; Wu, X.J. The influence of quality parameter selection on 3D virtual reconstruction model precision based on dry skull. Acta Anthropol. Sin. 2020, 39, 270–281. [Google Scholar] [CrossRef]
- Cignoni, P.; Callieri, M.; Corsini, M.; Dellepiane, M.; Ganovelli, F.; Ranzuglia, G. Meshlab: An open-source mesh processing tool. In Proceedings of the Eurographics Italian Chapter Conference, Salerno, Italy, 2–4 July 2008; Eurographics Association: Aire-la-Ville, Switzerland, 2008; pp. 129–136. [Google Scholar]
- Bne, A.; Louis, M.; Martin, B.; Durrleman, S. Deformetrica 4: An open-source software for statistical shape analysis. In Proceedings of the International Workshop on Shape in Medical Imaging, Granada, Spain, 20 September 2018; Lecture Notes in Computer Science. Springer: Cham, Switzerland, 2018; Volume 11167, pp. 3–13. [Google Scholar] [CrossRef] [Green Version]
- Durrleman, S. Statistical Models of Currents for Measuring the Variability of Anatomical Curves, Surfaces and Their Evolution. Ph.D. Thesis, Université Nice Sophia Antipolis, Nice, France, 2010. [Google Scholar]
- Dumoncel, J. RToolsForDeformetrica. 2020. Available online: https://gitlab.com/jeandumoncel/tools-for-deformetrica/-/tree/master/src/R_script/RToolsForDeformetrica (accessed on 14 December 2021).
- Dray, S.; Dufour, A.-B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Team, R. R: A language and environment for statistical computing. R foundation for statistical computing:Vienna, Austria. Computing 2009, 14, 12–21. [Google Scholar]
- Wickham, H. ggplot2; Springer: New York, NY, USA, 2009. [Google Scholar]
- Xiang, L.; Crow, T.; Roberts, N. Cerebral torque is human specific and unrelated to brain size. Brain Struct. Funct. 2019, 224, 1141–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, F.; Cai, Q.; Bogart, S.L.; Dubois, J.; Coulon, O.; Monzalvo, K.; Fischer, C.; Glasel, H.; Van der Haegen, L.; Bénézit, A.; et al. New human-specific brain landmark: The depth asymmetry of superior temporal sulcus. Proc. Natl. Acad. Sci. USA 2015, 112, 1208–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falzi, G.; Perrone, P.; Vignolo, L.A. Right-left asymmetry in anterior speech region. Arch. Neurol. 1982, 39, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Best, C.T. The emergence of cerebral asymmetries in early human development: A literature review and a neuroembryological model. In Brain Lateralization in Children: Developmental Implications; Molfese, D.L., Segalowitz, S.J., Eds.; Guilford Press: New York, NY, USA, 1988; pp. 5–34. [Google Scholar]
- Strick, P.L.; Dum, R.P.; Fiez, J.A. Cerebellum and nonmotor function. Annu. Rev. Neurosci. 2009, 32, 413–434. [Google Scholar] [CrossRef] [Green Version]
- Schmahmann, J.D. Disorders of the cerebellum: Ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J. Neuropsych. Clin. Neurosci. 2004, 16, 367–378. [Google Scholar] [CrossRef]
- Stoodley, C.J. The cerebellum and cognition: Evidence from functional imaging studies. Cerebellum 2012, 11, 352–365. [Google Scholar] [CrossRef]
- O’Reilly, J.X.; Beckmann, C.F.; Tomassini, V.; Ramnani, N.; Johansen-Berg, H. Distinct and overlapping functional zones in the cerebellum defined by resting state functional connectivity. Cereb. Cortex 2010, 20, 953–965. [Google Scholar] [CrossRef] [Green Version]
- Krienen, F.M.; Buckner, R.L. Segregated fronto-cerebellar circuits revealed by intrinsic functional connectivity. Cereb. Cortex 2009, 19, 2485–2497. [Google Scholar] [CrossRef] [Green Version]
- Buckner, R.L. The cerebellum and cognitive function: 25 years of insight from anatomy and neuroimaging. Neuron 2013, 80, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Foundas, A.L.; Leonard, C.M.; Gilmore, R.; Fennell, E.; Heilman, K.M. Planum temporale asymmetry and language dominance. Neuropsychologia 1994, 32, 1225–1231. [Google Scholar] [CrossRef]
- Beaudet, A. The emergence of language in the hominin lineage: Perspectives from fossil endocasts. Front. Hum. Neurosci. 2017, 11, 427. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, S.; Hublin, J.J.; Gunz, P. The evolution of modern human brain shape. Sci. Adv. 2018, 4, eaao5961. [Google Scholar] [CrossRef] [Green Version]
- Gannon, P.J.; Holloway, R.L.; Broadfield, D.C.; Braun, A.R. Asymmetry of chimpanzee planum temporale: Humanlike pattern of Wernicke’s brain language area homolog. Science 1998, 279, 220–222. [Google Scholar] [CrossRef]
- Cantalupo, C.; Hopkins, W.D. Asymmetric Broca’s area in great apes. Nature 2001, 414, 505. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Zhao, Y.; Xing, S. Asymmetry of Endocast Surface Shape in Modern Humans Based on Diffeomorphic Surface Matching. Symmetry 2022, 14, 1459. https://doi.org/10.3390/sym14071459
Lin S, Zhao Y, Xing S. Asymmetry of Endocast Surface Shape in Modern Humans Based on Diffeomorphic Surface Matching. Symmetry. 2022; 14(7):1459. https://doi.org/10.3390/sym14071459
Chicago/Turabian StyleLin, Sungui, Yuhao Zhao, and Song Xing. 2022. "Asymmetry of Endocast Surface Shape in Modern Humans Based on Diffeomorphic Surface Matching" Symmetry 14, no. 7: 1459. https://doi.org/10.3390/sym14071459
APA StyleLin, S., Zhao, Y., & Xing, S. (2022). Asymmetry of Endocast Surface Shape in Modern Humans Based on Diffeomorphic Surface Matching. Symmetry, 14(7), 1459. https://doi.org/10.3390/sym14071459