Ribosome Display Technology: Applications in Disease Diagnosis and Control



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
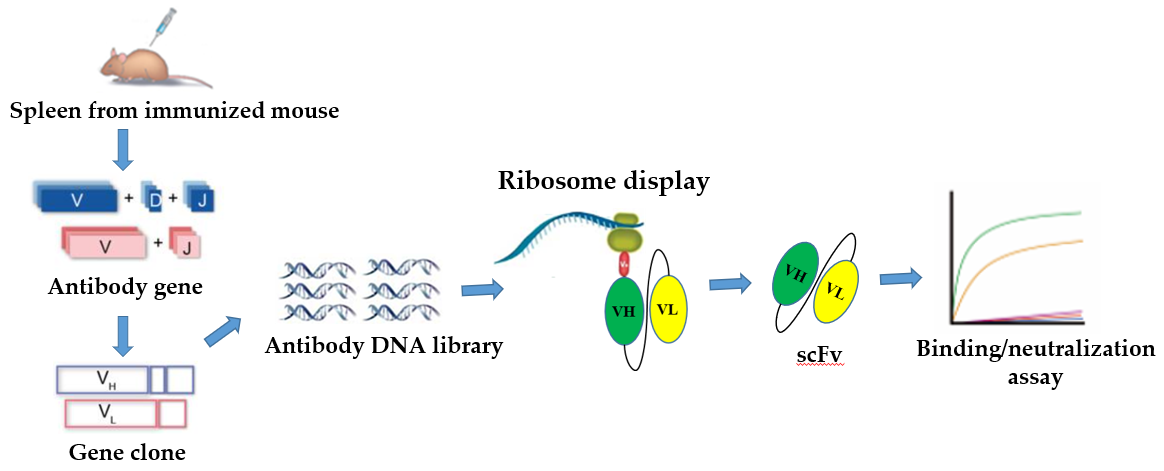
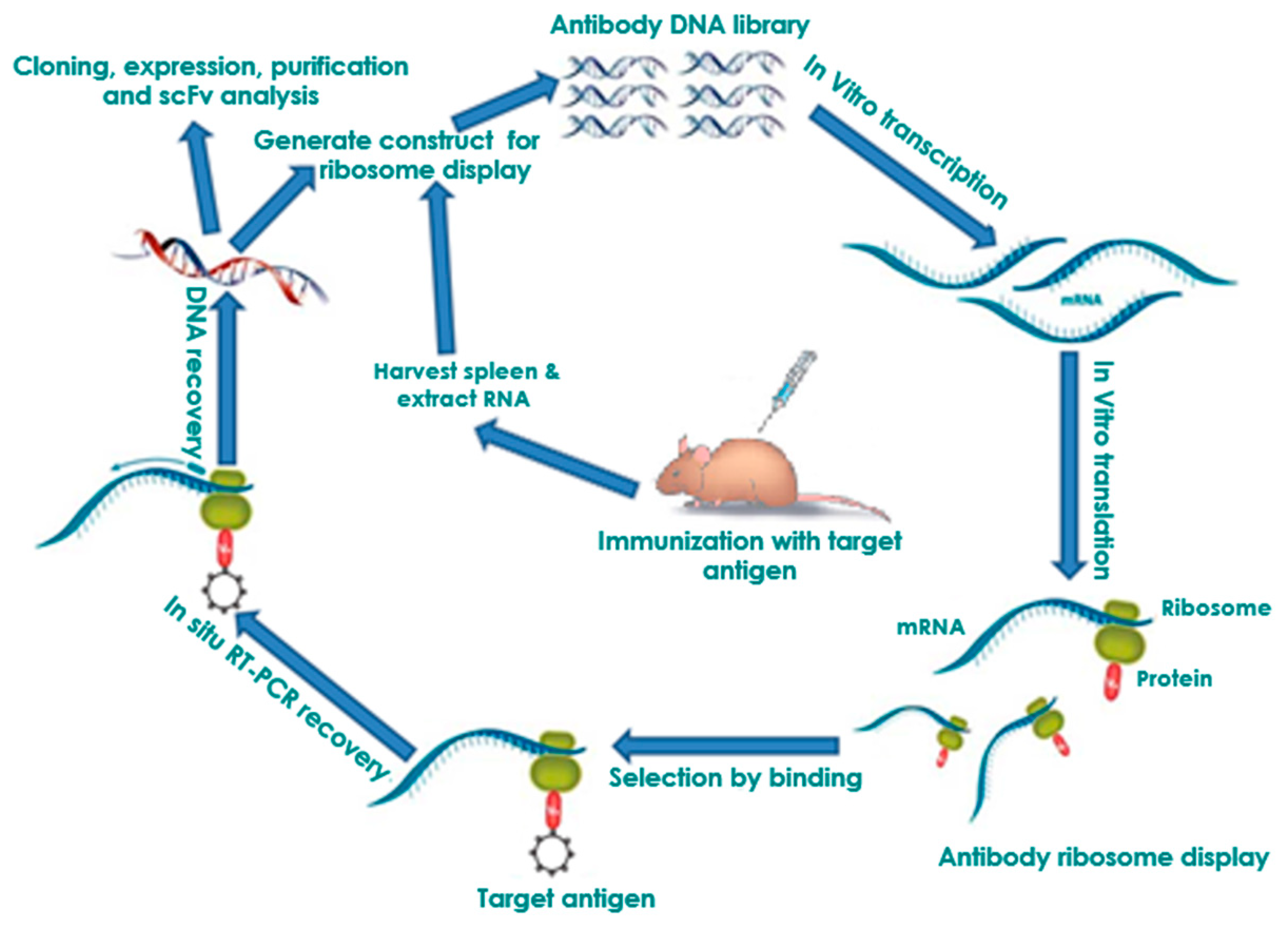
2. In Vitro Ribosome Display
2.1. Selection of Antibodies by Panning
2.2. Affinity Maturation and Modification of Ribosome Display Antibodies
2.3. Ribosome Display Antibody Gene Libraries
3. Ribosome Display Technology in Disease Diagnostics and Control
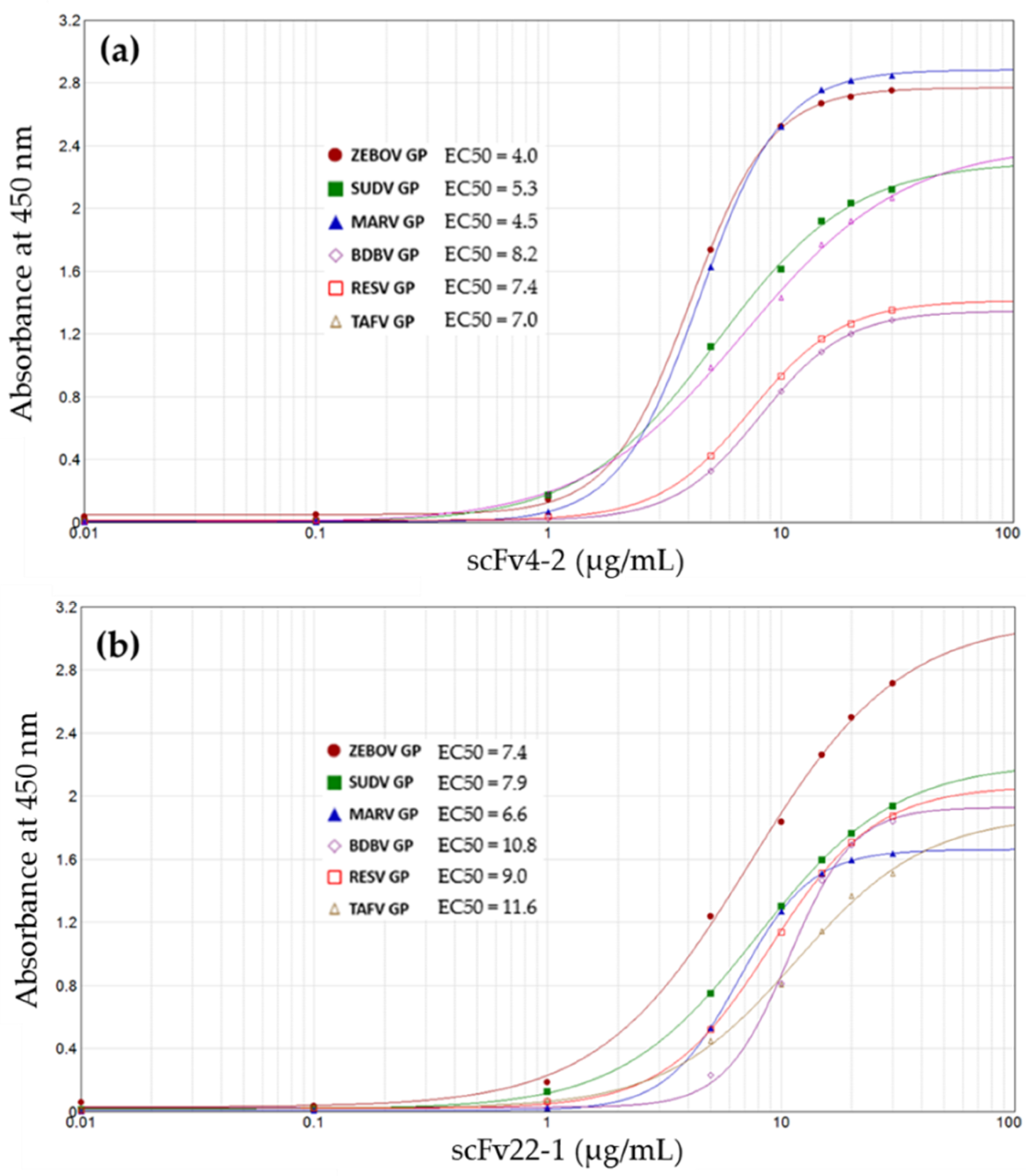
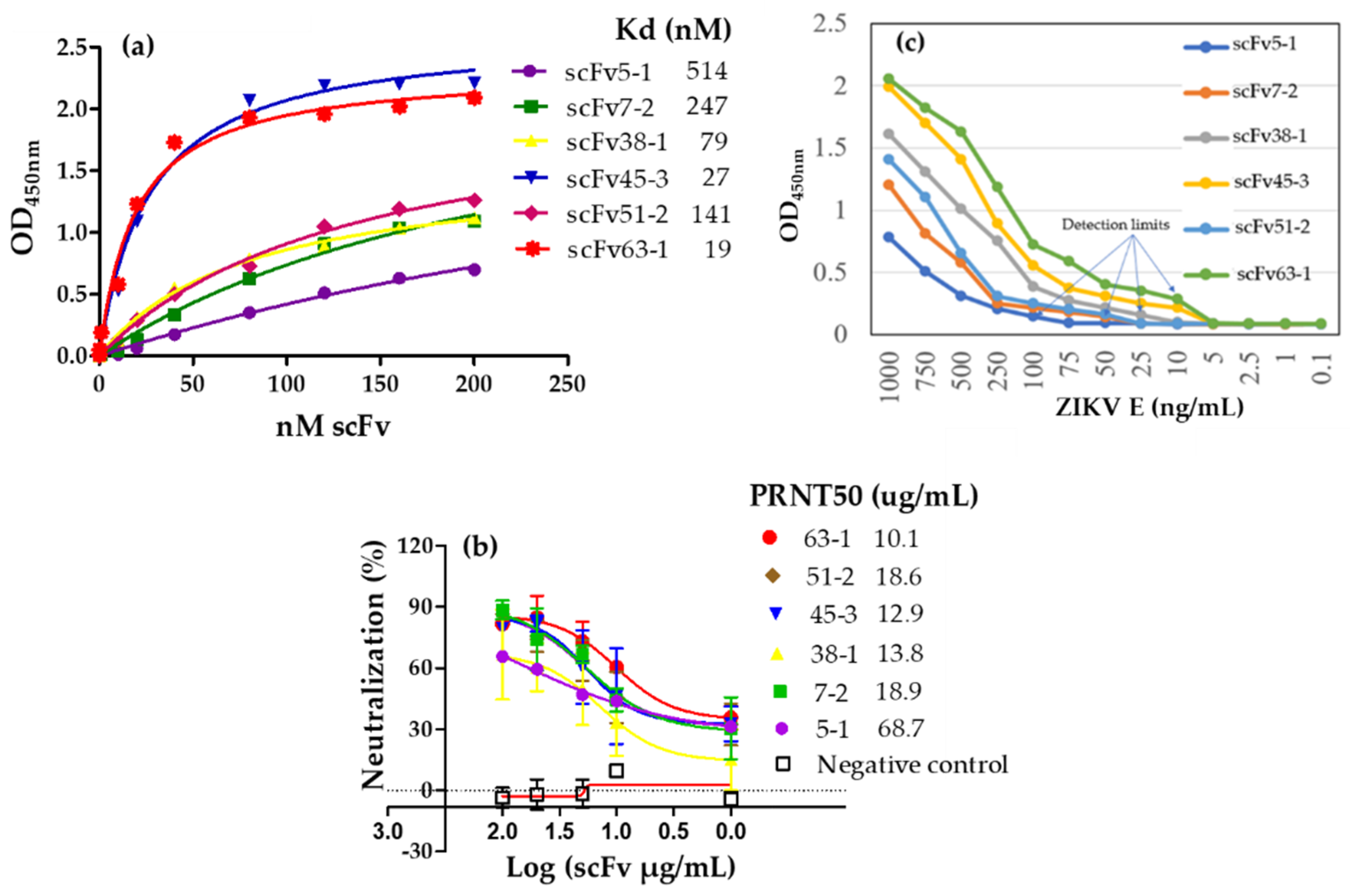
3.1. Human Infectious Diseases
3.2. Cancer
3.3. Acquired Immunodeficiency Syndrome (AIDS)
3.4. Plant Disease: Pierce’s Disease
4. Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ribosome Display Technology | RDT |
| Monoclonal antibodies | mAbs |
| Recombinant antibody | rAb |
| Single-chain fragment variable | scFv |
| Immunoglobulin G | IgG |
| Deoxyribonucleic acid | DNA |
| Ribonucleic acid | RNA |
| Polymerase chain reaction | PCR |
| Protein-ribosome-mRNA | PRM |
| Reverse transcription-PCR | RT-PCR |
| Enzyme-linked immunosorbent assay | ELISA |
| Terminal protein | TP |
| Designed Ankyrin Repeat Proteins | DARPins |
| Pierce’s Disease | PD |
| Glassy-winged sharpshooter | GWSS |
| Immunofluorescence assay | IFA |
| Hepatitis B virus | HBV |
| Ebola Virus | EBOV |
| Zaire ebolavirus | ZEBOV |
| Sudan virus | SUDV |
| Tai Forest virus | TAFV |
| Bundibugyo virus | BDBV |
| Marburg virus | MARV |
| Glycoprotein | GP |
| Zika virus | ZIKV |
| Coronavirus Disease 2019 | COVID-19 |
| Severe Acute Respiratory Syndrome | SARS |
| Middle East Respiratory Syndrome | MERS |
| Tuberculosis | TB |
| Early Secretory Antigenic Target-6 | ESAT-6 |
| Acquired Immunodeficiency Syndrome | AIDS |
| Human Immunodeficiency Virus | HIV |
| Simian Immunodeficiency Virus | SIV |
| Antiretroviral therapy | ART |
| Peripheral Blood Mononuclear Cell | PBMC |
References
- Weisser, N.E.; Hall, J.C. Applications of single-chain variable fragment antibodies in therapeutics and diagnostics. Biotechnol. Adv. 2009, 27, 502–520. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, M.A.; van de Winkel, J.G.J. Human antibodies as next generation therapeutics. Curr. Opin. Chem. Biol. 2001, 5, 368–374. [Google Scholar] [CrossRef]
- Taussig, M.J.; Stoevesandt, O.; Borrebaeck, C.A.; Bradbury, A.R.; Cahill, D.; Cambillau, C.; de Daruvar, A.; Dubel, S.; Eichler, J.; Frank, R.; et al. ProteomeBinders: Planning a European resource of affinity reagents for analysis of the human proteome. Nat. Methods 2007, 4, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.; Hamid, M. scFv antibody: Principles and clinical application. Clin. Dev. Immunol. 2012, 2012, 980250. [Google Scholar] [CrossRef] [PubMed]
- Farajnia, S.; Ahmadzadeh, V.; Tanomand, A.; Veisi, K.; Khosroshahi, S.A.; Rahbarnia, L. Development trends for generation of single-chain antibody fragments. Immunopharmacol. Immunotoxicol. 2014, 36, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Griffiths, A.D.; Hawkins, R.E.; Hoogenboom, H.R. Making antibodies by phage display technology. Annu. Rev. Immunol. 1994, 12, 433–455. [Google Scholar] [CrossRef]
- Georgiou, G.; Stathopoulos, C.; Daugherty, P.S.; Nayak, A.R.; Iverson, B.L.; Curtiss, R., 3rd. Display of heterologous proteins on the surface of microorganisms: From the screening of combinatorial libraries to live recombinant vaccines. Nat. Biotechnol. 1997, 15, 29–34. [Google Scholar] [CrossRef]
- Shusta, E.V.; VanAntwerp, J.; Wittrup, K.D. Biosynthetic polypeptide libraries. Curr. Opin. Biotechnol. 1999, 10, 117–122. [Google Scholar] [CrossRef]
- Mattheakis, L.C.; Bhatt, R.R.; Dower, W.J. An in vitro polysome display system for identifying ligands from very large peptide libraries. Proc. Natl. Acad. Sci. USA 1994, 91, 9022–9026. [Google Scholar] [CrossRef]
- Hanes, J.; Pluckthun, A. In vitro selection and evolution of functional proteins by using ribosome display. Proc Natl Acad Sci USA 1997, 94, 4937–4942. [Google Scholar] [CrossRef]
- He, M.; Taussig, M.J. Antibody-ribosome-mRNA (ARM) complexes as efficient selection particles for in vitro display and evolution of antibody combining sites. Nucleic Acids Res. 1997, 25, 5132–5134. [Google Scholar] [CrossRef]
- Roberts, R.W.; Szostak, J.W. RNA-peptide fusions for the in vitro selection of peptides and proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 12297–12302. [Google Scholar] [CrossRef]
- He, M.; Edwards, B.M.; Kastelic, D.; Taussig, M.J. Eukaryotic ribosome display with in situ DNA recovery. Methods Mol. Biol. 2012, 805, 75–85. [Google Scholar] [PubMed]
- He, M.; Taussig, M.J. Eukaryotic ribosome display with in situ DNA recovery. Nat. Methods 2007, 4, 281–288. [Google Scholar] [CrossRef]
- He, M.; Taussig, M.J. Ribosome display: Cell-free protein display technology. Brief. Funct. Genom. Proteom. 2002, 1, 204–212. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Taussig, M.J. Selection of recombinant antibodies by eukaryotic ribosome display. Methods Mol. Biol. 2008, 484, 193–205. [Google Scholar] [PubMed]
- Stafford, R.; Matsumoto, M.; Yin, G.; Cai, Q.; Fung, J.J.; Stephenson, H.; Gill, A.; You, M.; Lin, S.-H.; Wang, W.; et al. In vitro Fab display: A cell-free system for IgG discovery. Protein Eng. Des. Sel. Peds 2014, 27. [Google Scholar] [CrossRef]
- Mattheakis, L.C.; Dias, J.M.; Dower, W.J. Cell-free synthesis of peptide libraries displayed on polysomes. Methods Enzym. 1996, 267, 195–207. [Google Scholar]
- Gersuk, G.M.; Corey, M.J.; Corey, E.; Stray, J.E.; Kawasaki, G.H.; Vessella, R.L. High-affinity peptide ligands to prostate-specific antigen identified by polysome selection. Biochem. Biophys. Res. Commun. 1997, 232, 578–582. [Google Scholar] [CrossRef]
- Hanes, J.; Jermutus, L.; Weber-Bornhauser, S.; Bosshard, H.R.; Pluckthun, A. Ribosome display efficiently selects and evolves high-affinity antibodies in vitro from immune libraries. Proc. Natl. Acad. Sci. USA 1998, 95, 14130–14135. [Google Scholar] [CrossRef]
- Hanes, J.; Jermutus, L.; Schaffitzel, C.; Pluckthun, A. Comparison of Escherichia coli and rabbit reticulocyte ribosome display systems. Febs Lett. 1999, 450, 105–110. [Google Scholar] [CrossRef]
- Lee, M.S.; Kwon, M.H.; Kim, K.H.; Shin, H.J.; Park, S.; Kim, H.I. Selection of scFvs specific for HBV DNA polymerase using ribosome display. J. Immunol. Methods 2004, 284, 147–157. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Taussig, M.J. Ribosome display of antibodies: Expression, specificity and recovery in a eukaryotic system. J. Immunol. Methods 2005, 297, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Shin, H.J.; Kim, K.; Lee, M.S. A pseudoknot improves selection efficiency in ribosome display. Mol. Biotechnol. 2007, 36, 32–37. [Google Scholar] [CrossRef]
- Qi, Y.; Wu, C.; Zhang, S.; Wang, Z.; Huang, S.; Dai, L.; Wang, S.; Xia, L.; Wen, K.; Cao, X.; et al. Selection of anti-sulfadimidine specific ScFvs from a hybridoma cell by eukaryotic ribosome display. PLoS ONE 2009, 4, e6427. [Google Scholar] [CrossRef]
- Kastelic, D.; He, M. Ribosome display and screening for protein therapeutics. Methods Mol. Biol. 2012, 899, 61–72. [Google Scholar]
- Edwards, B.M.; He, M. Evolution of antibodies in vitro by ribosome display. Methods Mol. Biol. 2012, 907, 281–292. [Google Scholar]
- Douthwaite, J.A. Eukaryotic ribosome display selection using rabbit reticulocyte lysate. Methods Mol. Biol. 2012, 805, 45–57. [Google Scholar]
- Tang, J.; Wang, L.; Markiv, A.; Jeffs, S.A.; Dreja, H.; McKnight, A.; He, M.; Kang, A.S. Accessing of recombinant human monoclonal antibodies from patient libraries by eukaryotic ribosome display. Hum. Antibodies 2012, 21, 1–11. [Google Scholar] [CrossRef]
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: Affinity purification of target genes. Gene 1988, 73, 305–318. [Google Scholar] [CrossRef]
- Bittner, M.; Kupferer, P.; Morris, C.F. Electrophoretic transfer of proteins and nucleic acids from slab gels to diazobenzyloxymethyl cellulose or nitrocellulose sheets. Anal. Biochem. 1980, 102, 459–471. [Google Scholar] [CrossRef]
- Hawlisch, H.; Muller, M.; Frank, R.; Bautsch, W.; Klos, A.; Kohl, J. Site-specific anti-C3a receptor single-chain antibodies selected by differential panning on cellulose sheets. Anal. Biochem. 2001, 293, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.; Borgen, T.; Stacy, J.; Kausmally, L.; Simonsen, B.; Marvik, O.J.; Brekke, O.H.; Braunagel, M. Identification of scFv antibody fragments that specifically recognise the heroin metabolite 6-monoacetylmorphine but not morphine. J. Immunol. Methods 2003, 280, 139–155. [Google Scholar] [CrossRef]
- Breitling, F.; Dubel, S.; Seehaus, T.; Klewinghaus, I.; Little, M. A surface expression vector for antibody screening. Gene 1991, 104, 147–153. [Google Scholar] [CrossRef]
- Hust, M.; Maiss, E.; Jacobsen, H.J.; Reinard, T. The production of a genus-specific recombinant antibody (scFv) using a recombinant potyvirus protease. J. Virol. Methods 2002, 106, 225–233. [Google Scholar] [CrossRef]
- Barbas, C.F., 3rd; Kang, A.S.; Lerner, R.A.; Benkovic, S.J. Assembly of combinatorial antibody libraries on phage surfaces: The gene III site. Proc. Natl. Acad. Sci. USA 1991, 88, 7978–7982. [Google Scholar] [CrossRef] [PubMed]
- Sanna, P.P.; Williamson, R.A.; De Logu, A.; Bloom, F.E.; Burton, D.R. Directed selection of recombinant human monoclonal antibodies to herpes simplex virus glycoproteins from phage display libraries. Proc. Natl. Acad. Sci. USA 1995, 92, 6439–6443. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Milstein, C. Man-made antibodies. Nature 1991, 349, 293–299. [Google Scholar] [CrossRef]
- Hoogenboom, H.R. Selecting and screening recombinant antibody libraries. Nat Biotechnol 2005, 23, 1105–1116. [Google Scholar] [CrossRef]
- Hust, M.; Frenzel, A.; Schirrmann, T.; Dubel, S. Selection of recombinant antibodies from antibody gene libraries. Methods Mol Biol 2014, 1101, 305–320. [Google Scholar]
- Hust, M.; Dubel, S.; Schirrmann, T. Selection of recombinant antibodies from antibody gene libraries. Methods Mol. Biol. 2007, 408, 243–255. [Google Scholar] [PubMed]
- Hallborn, J.; Carlsson, R. Automated screening procedure for high-throughput generation of antibody fragments. Biotechniques 2002, 33 (Suppl. S6), 30–37. [Google Scholar] [CrossRef]
- Konthur, Z.; Hust, M.; Dubel, S. Perspectives for systematic in vitro antibody generation. Gene 2005, 364, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Schirrmann, T.; Al-Halabi, L.; Dubel, S.; Hust, M. Production systems for recombinant antibodies. Front Biosci. 2008, 13, 4576–4594. [Google Scholar] [CrossRef] [PubMed]
- Hust, M.; Meyer, T.; Voedisch, B.; Rulker, T.; Thie, H.; El-Ghezal, A.; Kirsch, M.I.; Schutte, M.; Helmsing, S.; Meier, D.; et al. A human scFv antibody generation pipeline for proteome research. J. Biotechnol. 2011, 152, 159–170. [Google Scholar] [CrossRef]
- Groves, M.A.; Nickson, A.A. Affinity maturation of phage display antibody populations using ribosome display. Methods Mol. Biol. 2012, 805, 163–1690. [Google Scholar] [PubMed]
- Kobayashi, N.; Oyama, H.; Kato, Y.; Goto, J.; Soderlind, E.; Borrebaeck, C.A. Two-step in vitro antibody affinity maturation enables estradiol-17beta assays with more than 10-fold higher sensitivity. Anal. Chem. 2010, 82, 1027–1038. [Google Scholar] [CrossRef]
- Finlay, W.J.; Cunningham, O.; Lambert, M.A.; Darmanin-Sheehan, A.; Liu, X.; Fennell, B.J.; Mahon, C.M.; Cummins, E.; Wade, J.M.; O’Sullivan, C.M.; et al. Affinity maturation of a humanized rat antibody for anti-RAGE therapy: Comprehensive mutagenesis reveals a high level of mutational plasticity both inside and outside the complementarity-determining regions. J. Mol. Biol. 2009, 388, 541–558. [Google Scholar] [CrossRef]
- Lipovsek, D.; Pluckthun, A. In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 2004, 290, 51–67. [Google Scholar] [CrossRef]
- Hanes, J.; Schaffitzel, C.; Knappik, A.; Pluckthun, A. Picomolar affinity antibodies from a fully synthetic naive library selected and evolved by ribosome display. Nat. Biotechnol. 2000, 18, 1287–1292. [Google Scholar] [CrossRef]
- Schaffitzel, C.; Berger, I.; Postberg, J.; Hanes, J.; Lipps, H.J.; Pluckthun, A. In vitro generated antibodies specific for telomeric guanine-quadruplex DNA react with Stylonychia lemnae macronuclei. Proc. Natl. Acad. Sci. USA 2001, 98, 8572–8577. [Google Scholar] [CrossRef] [PubMed]
- Jermutus, L.; Honegger, A.; Schwesinger, F.; Hanes, J.; Pluckthun, A. Tailoring in vitro evolution for protein affinity or stability. Proc. Natl. Acad. Sci. USA 2001, 98, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.M.; Weiss, G.A. Optimizing the affinity and specificity of proteins with molecular display. Mol. Biosyst. 2006, 2, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Dreier, B.; Pluckthun, A. Rapid selection of high-affinity binders using ribosome display. Methods Mol. Biol. 2012, 805, 261–286. [Google Scholar] [PubMed]
- Lewis, L.; Lloyd, C. Optimisation of antibody affinity by ribosome display using error-prone or site-directed mutagenesis. Methods Mol. Biol. 2012, 805, 139–161. [Google Scholar] [PubMed]
- Zahnd, C.; Spinelli, S.; Luginbuhl, B.; Amstutz, P.; Cambillau, C.; Pluckthun, A. Directed in vitro evolution and crystallographic analysis of a peptide-binding single chain antibody fragment (scFv) with low picomolar affinity. J. Biol. Chem. 2004, 279, 18870–18877. [Google Scholar] [CrossRef] [PubMed]
- Chin, S.E.; Ferraro, F.; Groves, M.; Liang, M.; Vaughan, T.J.; Dobson, C.L. Isolation of high affinity, neutralizing anti-idiotype antibodies by phage and ribosome display for application in immunogenicity and pharmacokinetic analyses. J. Immunol. Methods 2015, 416, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Groves, M.A.; Amanuel, L.; Campbell, J.I.; Rees, D.G.; Sridharan, S.; Finch, D.K.; Lowe, D.C.; Vaughan, T.J. Antibody VH and VL recombination using phage and ribosome display technologies reveals distinct structural routes to affinity improvements with VH-VL interface residues providing important structural diversity. MAbs 2014, 6, 236–245. [Google Scholar] [CrossRef]
- Hu, D.; Tateno, H.; Hirabayashi, J. Directed evolution of lectins by an improved error-prone PCR and ribosome display method. Methods Mol. Biol. 2014, 1200, 527–538. [Google Scholar]
- Kanamori, T.; Fujino, Y.; Ueda, T. PURE ribosome display and its application in antibody technology. Biochim. Biophys. Acta 2014, 1844, 1925–1932. [Google Scholar] [CrossRef]
- Darmanin-Sheehan, A.; Finlay, W.J.; Cunningham, O.; Fennell, B.J. Molecular scanning: Combining random mutagenesis, ribosome display, and bioinformatic analysis for protein engineering. Methods Mol. Biol. 2012, 907, 487–503. [Google Scholar] [PubMed]
- Lei, L. Identification of candidate vaccine genes using ribosome display. Methods Mol. Biol. 2012, 805, 299–314. [Google Scholar] [PubMed]
- Groves, M.A.; Osbourn, J.K. Applications of ribosome display to antibody drug discovery. Expert Opin. Biol. 2005, 5, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Chodorge, M.; Fourage, L.; Ravot, G.; Jermutus, L.; Minter, R. In vitro DNA recombination by L-Shuffling during ribosome display affinity maturation of an anti-Fas antibody increases the population of improved variants. Protein Eng. Des. Sel. Peds 2008, 21, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Heyduk, E.; Heyduk, T. Ribosome display enhanced by next generation sequencing: A tool to identify antibody-specific peptide ligands. Anal. Biochem. 2014, 464, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Kang, G.; Hu, M.; Huang, H. Ribosome Display: A Potent Display Technology used for Selecting and Evolving Specific Binders with Desired Properties. Mol. Biotechnol. 2019, 61, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Ryabova, L.A.; Desplancq, D.; Spirin, A.S.; Plückthun, A. Functional antibody production using cell-free translation: Effects of protein disulfide isomerase and chaperones. Nat. Biotechnol. 1997, 15, 79–84. [Google Scholar] [CrossRef]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: Filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar] [CrossRef]
- Glockshuber, R.; Malia, M.; Pfitzinger, I.; Plueckthun, A. A comparison of strategies to stabilize immunoglobulin Fv-fragments. Biochemistry 1990, 29, 1362–1367. [Google Scholar] [CrossRef]
- Groves, M.; Lane, S.; Douthwaite, J.; Lowne, D.; Rees, D.G.; Edwards, B.; Jackson, R.H. Affinity maturation of phage display antibody populations using ribosome display. J. Immunol. Methods 2006, 313, 129–139. [Google Scholar] [CrossRef]
- Sun, Y.; Ning, B.; Liu, M.; Gao, X.; Fan, X.; Liu, J.; Gao, Z. Selection of diethylstilbestrol-specific single-chain antibodies from a non-immunized mouse ribosome display library. PLoS ONE 2012, 7, e33186. [Google Scholar]
- Whiteaker, J.R.; Zhao, L.; Frisch, C.; Ylera, F.; Harth, S.; Knappik, A.; Paulovich, A.G. High-affinity recombinant antibody fragments (Fabs) can be applied in peptide enrichment immuno-MRM assays. J. Proteome Res. 2014, 13, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Ylera, F.; Harth, S.; Waldherr, D.; Frisch, C.; Knappik, A. Off-rate screening for selection of high-affinity anti-drug antibodies. Anal. Biochem. 2013, 441, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Binz, H.K.; Stumpp, M.T.; Forrer, P.; Amstutz, P.; Pluckthun, A. Designing repeat proteins: Well-expressed, soluble and stable proteins from combinatorial libraries of consensus ankyrin repeat proteins. J. Mol. Biol. 2003, 332, 489–503. [Google Scholar] [CrossRef]
- Binz, H.K.; Amstutz, P.; Kohl, A.; Stumpp, M.T.; Briand, C.; Forrer, P.; Grutter, M.G.; Pluckthun, A. High-affinity binders selected from designed ankyrin repeat protein libraries. Nat. Biotechnol. 2004, 22, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Dreier, B.; Pluckthun, A. Ribosome display: A technology for selecting and evolving proteins from large libraries. Methods Mol. Biol. 2011, 687, 283–306. [Google Scholar]
- Schilling, J.; Schoppe, J.; Pluckthun, A. From DARPins to LoopDARPins: Novel LoopDARPin design allows the selection of low picomolar binders in a single round of ribosome display. J. Mol. Biol. 2014, 426, 691–721. [Google Scholar] [CrossRef]
- Pluckthun, A. Ribosome display: A perspective. Methods Mol. Biol. 2012, 805, 3–28. [Google Scholar]
- Stefan, N.; Martin-Killias, P.; Wyss-Stoeckle, S.; Honegger, A.; Zangemeister-Wittke, U.; Pluckthun, A. DARPins recognizing the tumor-associated antigen EpCAM selected by phage and ribosome display and engineered for multivalency. J. Mol. Biol. 2011, 413, 826–843. [Google Scholar] [CrossRef]
- Schilling, J.; Schoppe, J.; Sauer, E.; Pluckthun, A. Co-crystallization with conformation-specific designed ankyrin repeat proteins explains the conformational flexibility of BCL-W. J. Mol. Biol. 2014, 426, 2346–2362. [Google Scholar] [CrossRef]
- Scholz, O.; Hansen, S.; Pluckthun, A. G-quadruplexes are specifically recognized and distinguished by selected designed ankyrin repeat proteins. Nucleic Acids Res. 2014, 42, 9182–9194. [Google Scholar] [CrossRef] [PubMed]
- Brauchle, M.; Hansen, S.; Caussinus, E.; Lenard, A.; Ochoa-Espinosa, A.; Scholz, O.; Sprecher, S.G.; Pluckthun, A.; Affolter, M. Protein interference applications in cellular and developmental biology using DARPins that recognize GFP and mCherry. Biol. Open 2014, 3, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Tamaskovic, R.; Simon, M.; Stefan, N.; Schwill, M.; Pluckthun, A. Designed ankyrin repeat proteins (DARPins) from research to therapy. Methods Enzym. 2012, 503, 101–134. [Google Scholar]
- Wetzel, S.K.; Ewald, C.; Settanni, G.; Jurt, S.; Pluckthun, A.; Zerbe, O. Residue-resolved stability of full-consensus ankyrin repeat proteins probed by NMR. J. Mol. Biol. 2010, 402, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, S.K.; Settanni, G.; Kenig, M.; Binz, H.K.; Pluckthun, A. Folding and unfolding mechanism of highly stable full-consensus ankyrin repeat proteins. J. Mol. Biol. 2008, 376, 241–257. [Google Scholar] [CrossRef]
- Schweizer, A.; Roschitzki-Voser, H.; Amstutz, P.; Briand, C.; Gulotti-Georgieva, M.; Prenosil, E.; Binz, H.K.; Capitani, G.; Baici, A.; Pluckthun, A.; et al. Inhibition of caspase-2 by a designed ankyrin repeat protein: Specificity, structure, and inhibition mechanism. Structure 2007, 15, 625–636. [Google Scholar] [CrossRef]
- Zahnd, C.; Wyler, E.; Schwenk, J.M.; Steiner, D.; Lawrence, M.C.; McKern, N.M.; Pecorari, F.; Ward, C.W.; Joos, T.O.; Pluckthun, A. A designed ankyrin repeat protein evolved to picomolar affinity to Her2. J. Mol. Biol. 2007, 369, 1015–1028. [Google Scholar] [CrossRef]
- Zahnd, C.; Pecorari, F.; Straumann, N.; Wyler, E.; Pluckthun, A. Selection and characterization of Her2 binding-designed ankyrin repeat proteins. J. Biol. Chem. 2006, 281, 35167–35175. [Google Scholar] [CrossRef]
- Amstutz, P.; Koch, H.; Binz, H.K.; Deuber, S.A.; Pluckthun, A. Rapid selection of specific MAP kinase-binders from designed ankyrin repeat protein libraries. Protein Eng. Des. Sel. Peds 2006, 19, 219–229. [Google Scholar] [CrossRef][Green Version]
- Amstutz, P.; Binz, H.K.; Parizek, P.; Stumpp, M.T.; Kohl, A.; Grutter, M.G.; Forrer, P.; Pluckthun, A. Intracellular kinase inhibitors selected from combinatorial libraries of designed ankyrin repeat proteins. J. Biol. Chem. 2005, 280, 24715–24722. [Google Scholar] [CrossRef]
- Dreier, B.; Mikheeva, G.; Belousova, N.; Parizek, P.; Boczek, E.; Jelesarov, I.; Forrer, P.; Pluckthun, A.; Krasnykh, V. Her2-specific multivalent adapters confer designed tropism to adenovirus for gene targeting. J. Mol. Biol. 2011, 405, 410–426. [Google Scholar] [CrossRef] [PubMed]
- Veesler, D.; Dreier, B.; Blangy, S.; Lichiere, J.; Tremblay, D.; Moineau, S.; Spinelli, S.; Tegoni, M.; Pluckthun, A.; Campanacci, V.; et al. Crystal structure and function of a DARPin neutralizing inhibitor of lactococcal phage TP901–1: Comparison of DARPin and camelid VHH binding mode. J. Bio.l Chem. 2009, 284, 30718–30726. [Google Scholar] [CrossRef] [PubMed]
- Milovnik, P.; Ferrari, D.; Sarkar, C.A.; Pluckthun, A. Selection and characterization of DARPins specific for the neurotensin receptor 1. Protein Eng. Des. Sel. Peds 2009, 22, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Scholz, O.; Henssler, E.M.; Bail, J.; Schubert, P.; Bogdanska-Urbaniak, J.; Sopp, S.; Reich, M.; Wisshak, S.; Kostner, M.; Bertram, R.; et al. Activity reversal of Tet repressor caused by single amino acid exchanges. Mol. Microbiol. 2004, 53, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Yau, K.Y.; Dubuc, G.; Li, S.; Hirama, T.; Mackenzie, C.R.; Jermutus, L.; Hall, J.C.; Tanha, J. Affinity maturation of a V(H)H by mutational hotspot randomization. J. Immunol. Methods 2005, 297, 213–224. [Google Scholar] [CrossRef]
- Perruchini, C.; Pecorari, F.; Bourgeois, J.P.; Duyckaerts, C.; Rougeon, F.; Lafaye, P. Llama VHH antibody fragments against GFAP: Better diffusion in fixed tissues than classical monoclonal antibodies. Acta Neuropathol. 2009, 118, 685–695. [Google Scholar] [CrossRef]
- WHO. Ebola Virus Disease (Fact Sheets). Archived 30th May 2019. Available online: https://www.who.int/en/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 4 February 2020).
- Kunamneni, A.; Clarke, E.C.; Ye, C.; Bradfute, S.B.; Durvasula, R. Generation and Selection of a Panel of Pan-Filovirus Single-Chain Antibodies using Cell-Free Ribosome Display. Am. J. Trop. Med. Hyg. 2019, 101, 198–206. [Google Scholar] [CrossRef]
- Malone, R.W.; Homan, J.; Callahan, M.V.; Glasspool-Malone, J.; Damodaran, L.; Schneider Ade, B.; Zimler, R.; Talton, J.; Cobb, R.R.; Ruzic, I.; et al. Zika Virus: Medical Countermeasure Development Challenges. Plos Negl. Trop. Dis. 2016, 10, e0004530. [Google Scholar] [CrossRef]
- WHO. Zika Virus (Fact Sheets). Archived 20th July 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/zika-virus (accessed on 4 February 2020).
- Kunamneni, A.; Ye, C.; Bradfute, S.B.; Durvasula, R. Ribosome display for the rapid generation of high-affinity Zika-neutralizing single-chain antibodies. PLoS ONE 2018, 13, e0205743. [Google Scholar] [CrossRef]
- WHO. Tuberculosis. Archived 24th March, 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 12 May 2020).
- Getahun, H.; Harrington, M.; O’Brien, R.; Nunn, P. Diagnosis of smear-negative pulmonary tuberculosis in people with HIV infection or AIDS in resource-constrained settings: Informing urgent policy changes. Lancet 2007, 369, 2042–2049. [Google Scholar] [CrossRef]
- Ahangarzadeh, S.; Bandehpour, M.; Kazemi, B. Selection of single-chain variable fragments specific for Mycobacterium tuberculosis ESAT-6 antigen using ribosome display. Iran J. Basic Med. Sci. 2017, 20, 327–333. [Google Scholar] [PubMed]
- WHO. Q&A on Coronaviruses (COVID-19). Archived 17th April, 2020. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/question-and-answers-hub/q-a-detail/q-a-coronaviruses (accessed on 12 May 2020).
- (WHO), W.H.O. Coronavirus disease (COVID-19) Pandemic. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 12 May 2020).
- Liu, Z.-X.; Yi, G.-H.; Qi, Y.-P.; Liu, Y.-L.; Yan, J.-P.; Qian, J.; Du, E.-Q.; Ling, W.-F. Identification of single-chain antibody fragments specific against SARS-associated coronavirus from phage-displayed antibody library. Biochem. Biophys. Res. Commun. 2005, 329, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.M.; Feng, D.X.; Lou, J.; Zhou, Y.; Fung, K.P.; Waye, M.M.Y.; Tsui, S.K.W.; Chan, P.K.S.; Marks, J.D.; Pang, S.F.; et al. Development of human single-chain antibodies against SARS-associated coronavirus. Intervirology 2008, 51. [Google Scholar] [CrossRef]
- Zhu, Z.; Chakraborti, S.; He, Y.; Roberts, A.; Sheahan, T.; Xiao, X.; Hensley, L.E.; Prabakaran, P.; Rockx, B.; Sidorov, I.A.; et al. Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc. Natl. Acad. Sci. USA 2007, 104, 12123–12128. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, Y.; Huang, J.; Jiang, S.; Du, L. Advances in MERS-CoV Vaccines and Therapeutics Based on the Receptor-Binding Domain. Viruses 2019, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Hillyer, C.; Du, L. Neutralizing Antibodies against SARS-CoV-2 and Other Human Coronaviruses. Trends Immunol. 2020, 41, 355–359. [Google Scholar] [CrossRef]
- Schaffitzel, C.; Hanes, J.; Jermutus, L.; Pluckthun, A. Ribosome display: An in vitro method for selection and evolution of antibodies from libraries. J. Immunol. Methods 1999, 231, 119–135. [Google Scholar] [CrossRef]
- Huang, S.; Feng, L.; An, G.; Zhang, X.; Zhao, Z.; Han, R.; Lei, F.; Zhang, Y.; Luo, A.; Jing, X.; et al. Ribosome display and selection of single-chain variable fragments effectively inhibit growth and progression of microspheres in vitro and in vivo. Cancer Sci. 2018, 109, 1503–1512. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC), About HIV/AIDS. Archived 2nd December, 2019. Available online: https://www.cdc.gov/hiv/basics/whatishiv.html (accessed on 17 May 2020).
- WHO. HIV/AIDS. Archived 17th April, 2020. Available online: https://www.who.int/health-topics/hiv-aids/#tab=tab_1 (accessed on 17 May 2020).
- Burton, D.R.; Weiss, R.A. AIDS/HIV. A boost for HIV vaccine design. Science 2010, 329, 770–773. [Google Scholar] [CrossRef]
- Scheid, J.F.; Mouquet, H.; Ueberheide, B.; Diskin, R.; Klein, F.; Oliveira, T.; Pietzsch, J.; Fenyö, D.; Abadir, A.; Velinzon, K.; et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011, 333, 1633–1637. [Google Scholar]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.J.; Purcell, A.H.; Thomson, S.V. Pierce’s disease of grapevines: Isolation of the causal bacterium. Science 1978, 199, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Morano, L.D.; Bextine, B.R.; Garcia, D.A.; Maddox, S.V.; Gunawan, S.; Vitovsky, N.J.; Black, M.C. Initial genetic analysis of Xylella fastidiosa in Texas. Curr. Microbiol. 2008, 56, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Lacava, P.T.; Miller, T.A. Detection of the bacterium, Xylella fastidiosa, in saliva of glassy-winged sharpshooter, Homalodisca vitripennis. J. Insect Sci. 2008, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Myers, A.L.; Sutton, T.B.; Abad, J.A.; Kennedy, G.G. Pierce’s Disease of Grapevines: Identification of the Primary Vectors in North Carolina. Phytopathology 2007, 97, 1440–1450. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.C.; Blua, M.J.; Bextine, B. Impact of duration versus frequency of probing by Homalodisca vitripennis (Hemiptera: Cicadellidae) on inoculation of Xylella fastidiosa. J. Econ. Entomol. 2008, 101, 1122–1126. [Google Scholar] [CrossRef]
- Hendson, M.; Purcell, A.H.; Chen, D.; Smart, C.; Guilhabert, M.; Kirkpatrick, B. Genetic diversity of Pierce’s disease strains and other pathotypes of Xylella fastidiosa. Appl. Environ. Microbiol. 2001, 67, 895–903. [Google Scholar] [CrossRef]
- Simpson, A.J.; Reinach, F.C.; Arruda, P.; Abreu, F.A.; Acencio, M.; Alvarenga, R.; Alves, L.M.; Araya, J.E.; Baia, G.S.; Baptista, C.S.; et al. The genome sequence of the plant pathogen Xylella fastidiosa. The Xylella fastidiosa Consortium of the Organization for Nucleotide Sequencing and Analysis. Nature 2000, 406, 151–159. [Google Scholar] [CrossRef]
- Retchless, A.; Labroussaa, F.; Shapiro, L.; Stenger, D.; Lindow, S.; Almeida, R. Genomic Insights into Xylella Fastidiosa Interactions with Plant and Insect Hosts; Springer: Berlin, Germany, 2014; pp. 177–202. [Google Scholar]
- Caserta, R.; Takita, M.A.; Targon, M.L.; Rosselli-Murai, L.K.; de Souza, A.P.; Peroni, L.; Stach-Machado, D.R.; Andrade, A.; Labate, C.A.; Kitajima, E.W.; et al. Expression of Xylella fastidiosa fimbrial and afimbrial proteins during biofilm formation. Appl. Environ. Microbiol. 2010, 76, 4250–4259. [Google Scholar] [CrossRef]
- Fjellbirkeland, A.; Bemanian, V.; McDonald, I.R.; Murrell, J.C.; Jensen, H.B. Molecular analysis of an outer membrane protein, MopB, of Methylococcus capsulatus (Bath) and structural comparisons with proteins of the OmpA family. Arch. Microbiol. 2000, 173, 346–351. [Google Scholar] [CrossRef]
- Voegel, T.M.; Warren, J.G.; Matsumoto, A.; Igo, M.M.; Kirkpatrick, B.C. Localization and characterization of Xylella fastidiosa haemagglutinin adhesins. Microbiology 2010, 156 Pt 7, 2172–2179. [Google Scholar] [CrossRef]
- Pierce, B.K.; Voegel, T.; Kirkpatrick, B.C. The Xylella fastidiosa PD1063 protein is secreted in association with outer membrane vesicles. PLoS ONE 2014, 9, e113504. [Google Scholar] [CrossRef] [PubMed]
- Bishop, R.E. Structural biology of membrane-intrinsic beta-barrel enzymes: Sentinels of the bacterial outer membrane. Biochim. Et Biophys. Acta 2008, 1778, 1881–1896. [Google Scholar] [CrossRef] [PubMed]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Kostakioti, M.; Newman, C.L.; Thanassi, D.G.; Stathopoulos, C. Mechanisms of protein export across the bacterial outer membrane. J. Bacteriol. 2005, 187, 4306–4314. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Wu, C.H.; Lin, J.W.; Weng, S.F.; Tseng, Y.H. Mutation of the gene encoding a major outer-membrane protein in Xanthomonas campestris pv. campestris causes pleiotropic effects, including loss of pathogenicity. Microbiology 2010, 156 Pt 9, 2842–2854. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, N.; Wakebe, H.; Yoshihara, E.; Nakae, T.; Nishino, T. Role of protein F in maintaining structural integrity of the Pseudomonas aeruginosa outer membrane. J. Bacteriol. 1989, 171, 983–990. [Google Scholar] [CrossRef]
- Woodruff, W.A.; Hancock, R.E. Pseudomonas aeruginosa outer membrane protein F: Structural role and relationship to the Escherichia coli OmpA protein. J. Bacteriol. 1989, 171, 3304–3309. [Google Scholar] [CrossRef]
- Wang, Y. The function of OmpA in Escherichia coli. Biochem. Biophys. Res. Commun. 2002, 292, 396–401. [Google Scholar] [CrossRef]
- Khan, N.A.; Shin, S.; Chung, J.W.; Kim, K.J.; Elliott, S.; Wang, Y.; Kim, K.S. Outer membrane protein A and cytotoxic necrotizing factor-1 use diverse signaling mechanisms for Escherichia coli K1 invasion of human brain microvascular endothelial cells. Microb. Pathog. 2003, 35, 35–42. [Google Scholar] [CrossRef]
- Prasadarao, N.V.; Wass, C.A.; Weiser, J.N.; Stins, M.F.; Huang, S.H.; Kim, K.S. Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infect. Immun. 1996, 64, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Rashed, A.; Almeida, R.P. Disrupting the transmission of a vector-borne plant pathogen. Appl. Environ. Microbiol. 2012, 78, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Lampe, D.J.; Lauzon, C.R.; Miller, T. Development of symbiotic control of Pierce’s Disease. 2016. Available online: https://biopesticide.ucr.edu/abstracts/assets/Lampe_abstract.pdf (accessed on 12 April 2020).
- Miller, T. Symbiotic Control in agriculture and medicine. Comp. Biochem. Physiol. A Mol. Integr. Physiol. Comp. Biochem. Physiol. Pt A 2007, 146. [Google Scholar]
- Azizi, A.; Arora, A.; Markiv, A.; Lampe, D.J.; Miller, T.A.; Kang, A.S. Ribosome display of combinatorial antibody libraries derived from mice immunized with heat-killed Xylella fastidiosa and the selection of MopB-specific single-chain antibodies. Appl. Environ. Microbiol. 2012, 78, 2638–2647. [Google Scholar] [CrossRef] [PubMed]
- Feil, H.; Feil, W.S.; Lindow, S.E. Contribution of Fimbrial and Afimbrial Adhesins of Xylella fastidiosa to Attachment to Surfaces and Virulence to Grape. Phytopathology 2007, 97, 318–324. [Google Scholar] [CrossRef]
- De La Fuente, L.; Burr, T.J.; Hoch, H.C. Autoaggregation of Xylella fastidiosa cells is influenced by type I and type IV pili. Appl. Environ. Microbiol. 2008, 74, 5579–5582. [Google Scholar] [CrossRef]
- Hoch, H.C. Continued assessment of Xylella fastidiosa fimbrial adhesins as important virulence factors in Pierce’s disease: Influence of xylem sap, p 87–91. In Proceedings of the 2010 Pierce’s Disease Research Symposium; Esser, T., Ed.; California Department of Food and Agriculture: Sacramento, CA, USA, 2010. [Google Scholar]
- Li, Y.; Hao, G.; Galvani, C.D.; Meng, Y.; De La Fuente, L.; Hoch, H.C.; Burr, T.J. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology 2007, 153 Pt 3, 719–726. [Google Scholar] [CrossRef]
- Markiv, A.; Anani, B.; Durvasula, R.V.; Kang, A.S. Module based antibody engineering: A novel synthetic REDantibody. J. Immunol. Methods 2011, 364, 40–49. [Google Scholar] [CrossRef]
- Markiv, A.; Beatson, R.; Burchell, J.; Durvasula, R.V.; Kang, A.S. Expression of recombinant multi-coloured fluorescent antibodies in gor -/trxB- E. coli cytoplasm. BMC Biotechnol. 2011, 11, 117. [Google Scholar] [CrossRef]
- Fang, W.; Vega-Rodriguez, J.; Ghosh, A.K.; Jacobs-Lorena, M.; Kang, A.; St Leger, R.J. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 2011, 331, 1074–1077. [Google Scholar] [CrossRef]
- Bukhari, T.; Takken, W.; Koenraadt, C.J. Development of Metarhizium anisopliae and Beauveria bassiana formulations for control of malaria mosquito larvae. Parasites Vectors 2011, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.A. Paratransgenesis as a potential tool for pest control: Review of applied arthropod symbiosis. J. Appl. Entomol. 2011, 135, 474–478. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunamneni, A.; Ogaugwu, C.; Bradfute, S.; Durvasula, R. Ribosome Display Technology: Applications in Disease Diagnosis and Control. Antibodies 2020, 9, 28. https://doi.org/10.3390/antib9030028
Kunamneni A, Ogaugwu C, Bradfute S, Durvasula R. Ribosome Display Technology: Applications in Disease Diagnosis and Control. Antibodies. 2020; 9(3):28. https://doi.org/10.3390/antib9030028
Chicago/Turabian StyleKunamneni, Adinarayana, Christian Ogaugwu, Steven Bradfute, and Ravi Durvasula. 2020. "Ribosome Display Technology: Applications in Disease Diagnosis and Control" Antibodies 9, no. 3: 28. https://doi.org/10.3390/antib9030028
APA StyleKunamneni, A., Ogaugwu, C., Bradfute, S., & Durvasula, R. (2020). Ribosome Display Technology: Applications in Disease Diagnosis and Control. Antibodies, 9(3), 28. https://doi.org/10.3390/antib9030028