
Humanized VHH-hFc Fusion Proteins Targeting the L-HN Fragment of Tetanus Toxin Provided Protection In Vivo
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents, Cell Lines, and Animals
2.2. Immunization of the Camel
2.3. Construction of the Phage-Display Nanobody Library
2.4. Biopanning and Isolation of the TL-HN-Specific VHHs
2.5. Construction, Expression, and Purification of VHH-hFc Fusion Protein
2.6. Cross-Reaction and Binding Activity of the VHH-hFc Fusion Proteins
2.7. KD Analysis and Cross-Competitive Binding Assay
2.8. Neutralizing Activity
2.9. Humanization Modification and Activity Evaluation of VHH-hFc Fusion Proteins
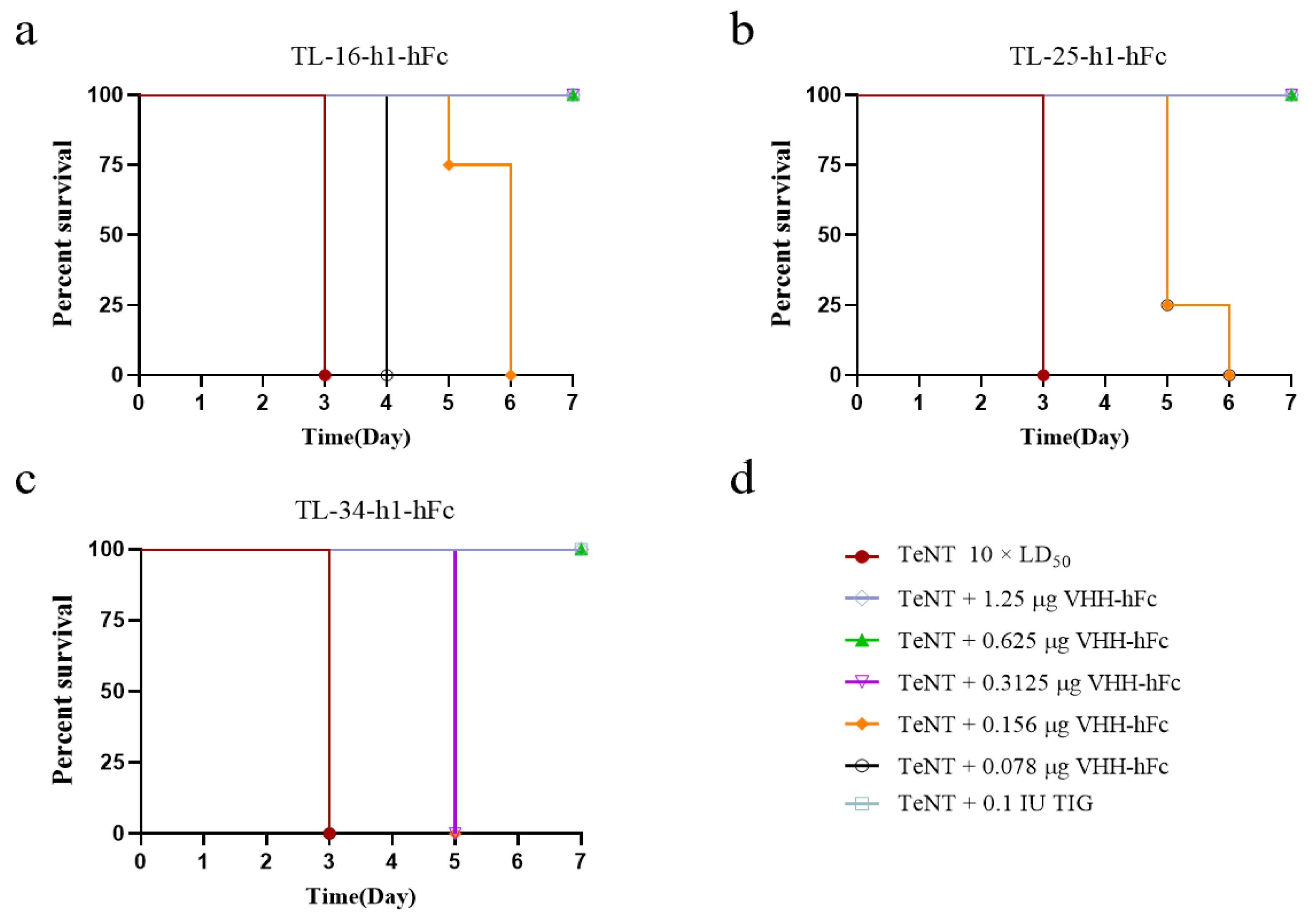
2.10. Preventive and Therapeutic Effects of the Humanized VHH-hFc Fusion Proteins in a Mouse Model
2.11. Statistical Analysis
3. Results
3.1. Construction and Biopanning of the Phage-Display VHH Library
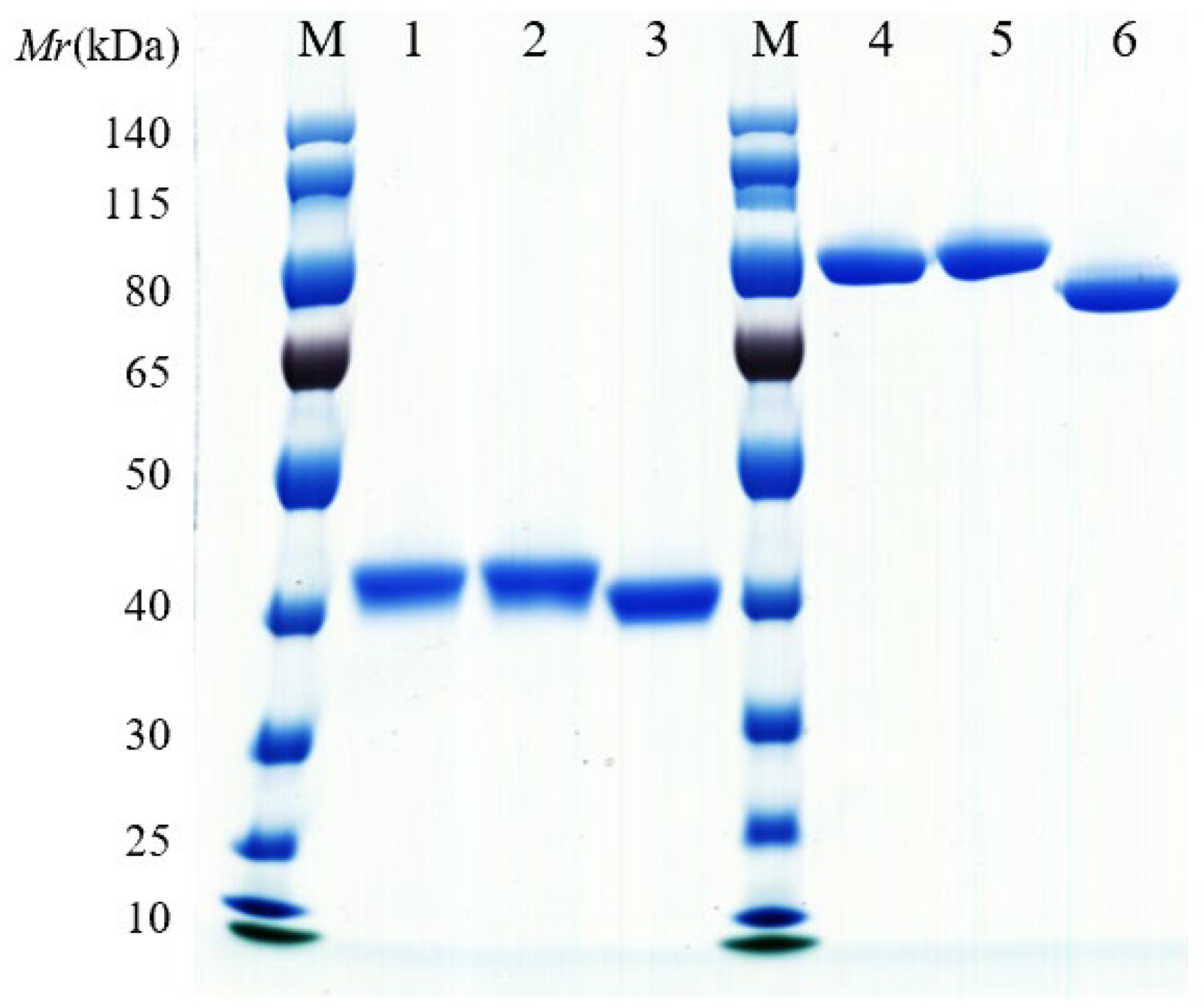
3.2. Expression and Purification of VHH-hFc Fusion Proteins
3.3. Preliminary Screening of VHH-hFc Fusion Proteins with Effective Neutralizing Activity In Vivo
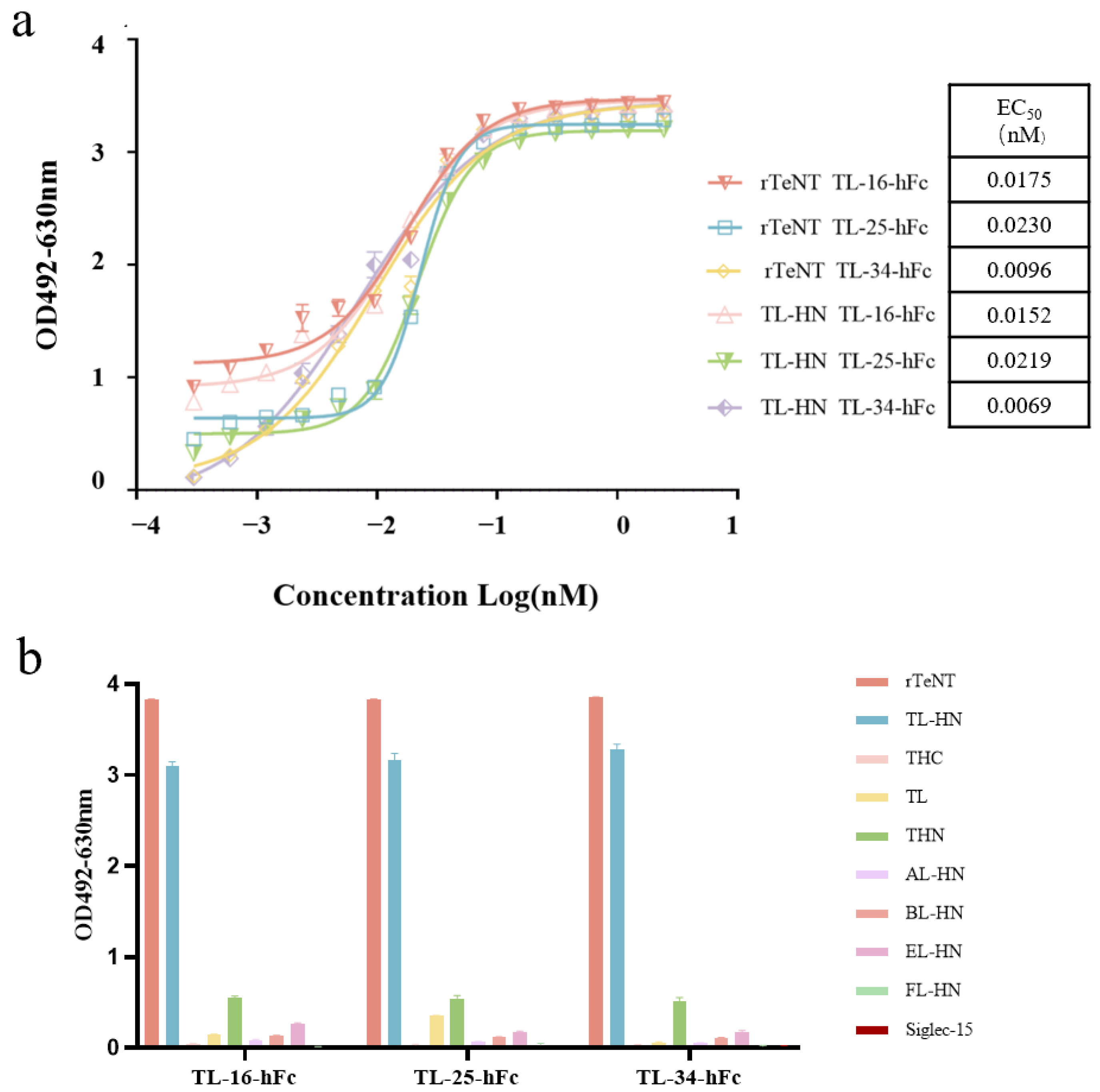
3.4. Binding Activity and Cross-Reactivity of the VHH-hFc Fusion Proteins
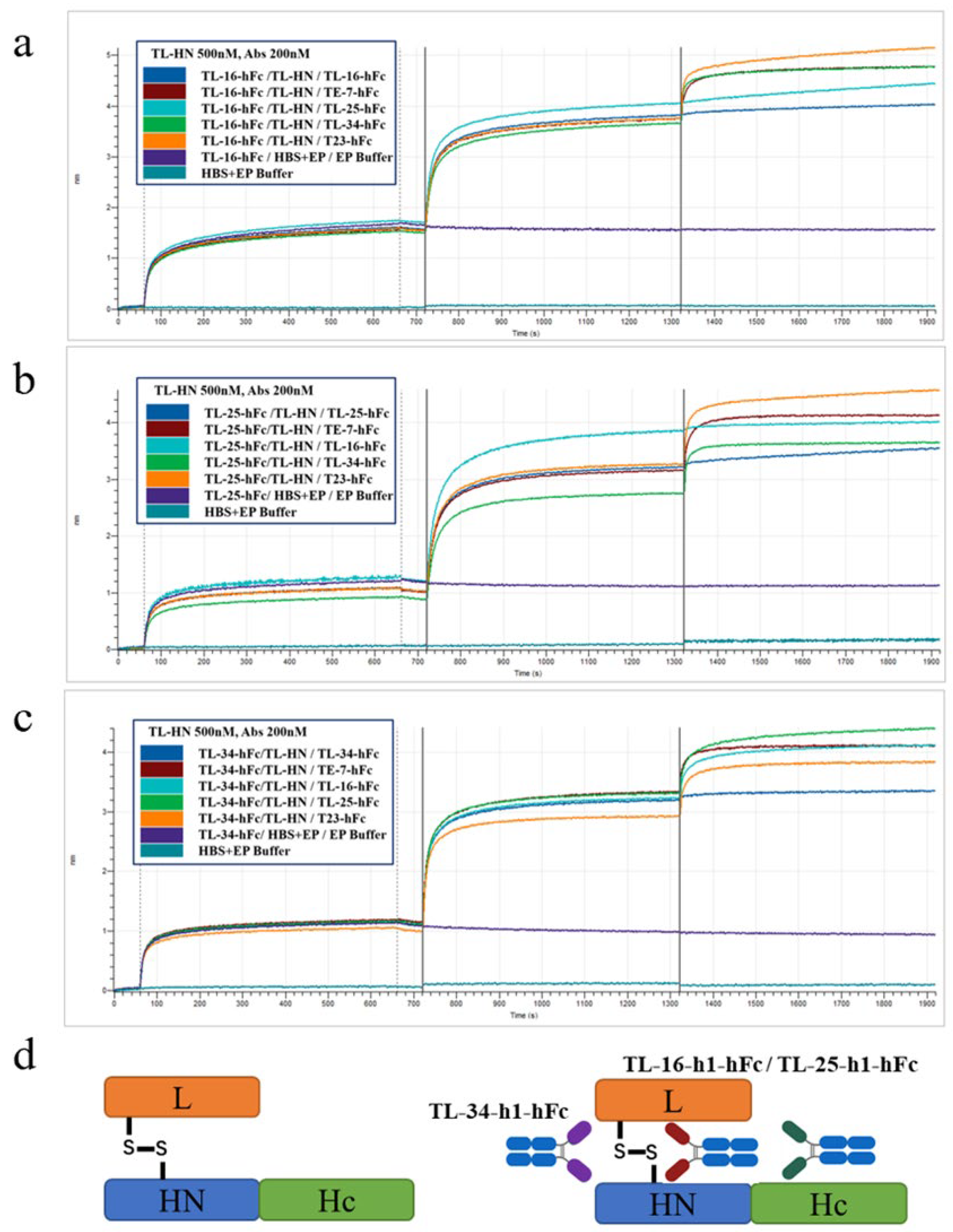
3.5. Affinity and Competitive Binding Activity of the VHH-hFc Fusion Proteins
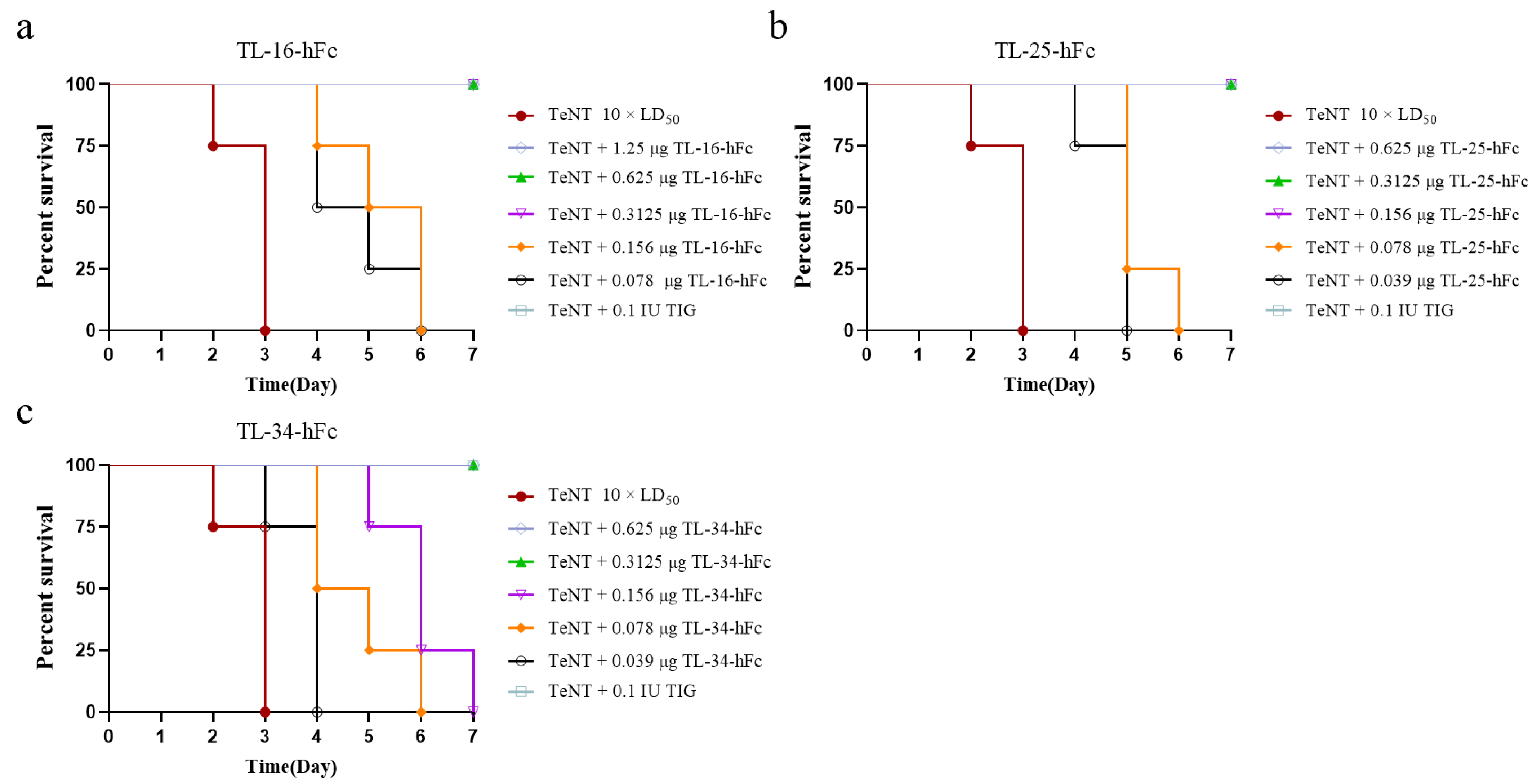
3.6. Neutralizing Activity of VHH-hFc Fusion Proteins at Different Doses
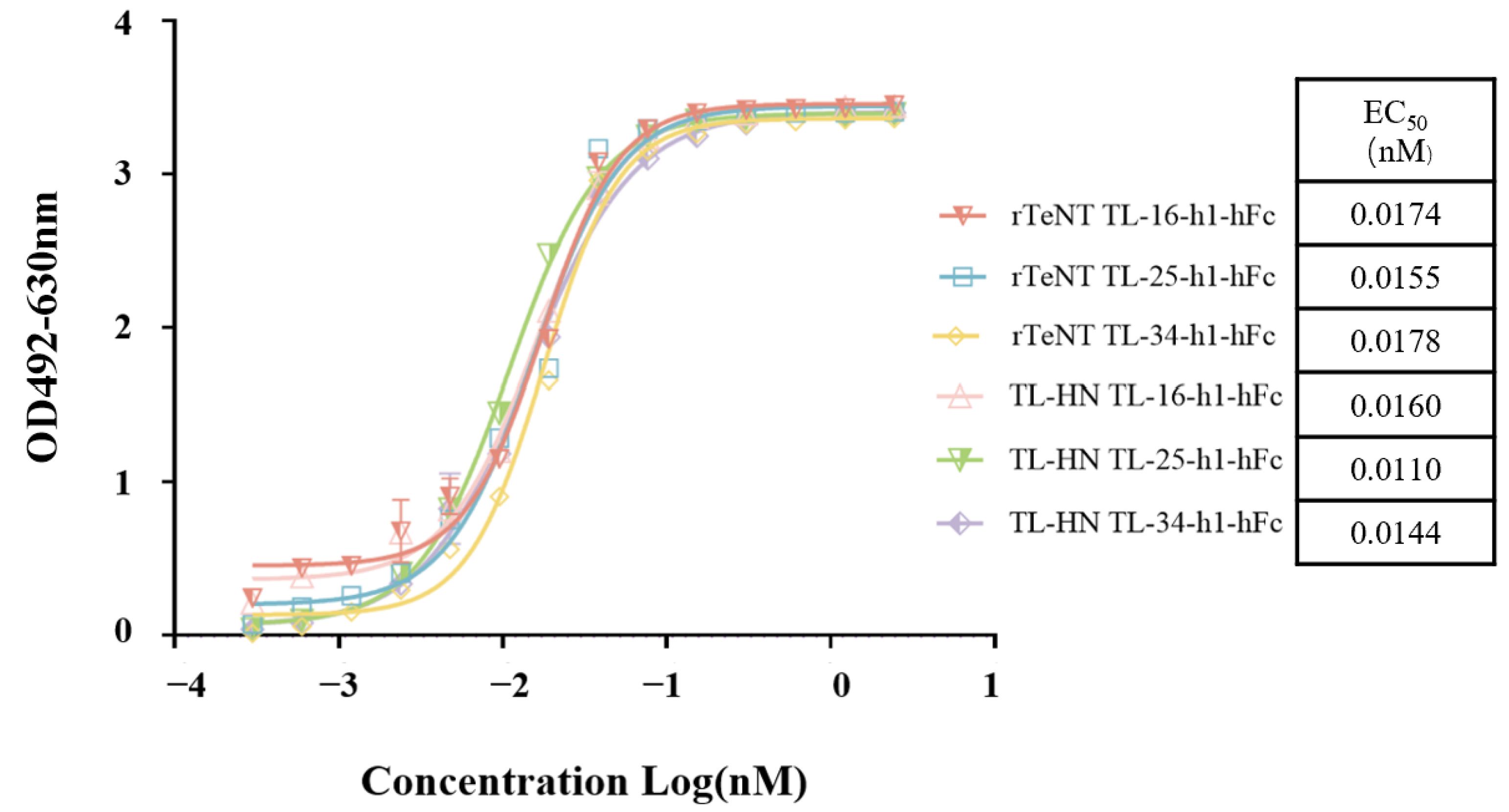
3.7. Humanization and Characterization of the Humanized VHH-hFc Fusion Proteins
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cook, T.M.; Protheroe, R.T.; Handel, J.M. Tetanus: A review of the literature. Br. J. Anaesth. 2001, 87, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Yen, L.M.; Thwaites, C.L. Tetanus. Lancet 2019, 393, 1657–1668. [Google Scholar] [CrossRef]
- Megighian, A.; Pirazzini, M.; Fabris, F.; Rossetto, O.; Montecucco, C. Tetanus and tetanus neurotoxin: From peripheral uptake to central nervous tissue targets. J. Neurochem. 2021, 158, 1244–1253. [Google Scholar] [CrossRef]
- Finkelstein, P.; Teisch, L.; Allen, C.J.; Ruiz, G. Tetanus: A Potential Public Health Threat in Times of Disaster. Prehosp. Disaster Med. 2017, 32, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Goonetilleke, A.; Harris, J.B. Clostridial neurotoxins. J. Neurol. Neurosurg. Psychiatry 2004, 75 (Suppl. 3), iii35–iii39. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Papini, E.; Genna, G.; Montecucco, C. An intact interchain disulfide bond is required for the neurotoxicity of tetanus toxin. Infect. Immun. 1990, 58, 4136–4141. [Google Scholar] [CrossRef]
- Fotinou, C.; Emsley, P.; Black, I.; Ando, H.; Ishida, H.; Kiso, M.; Sinha, K.A.; Fairweather, N.F.; Isaacs, N.W. The crystal structure of tetanus toxin Hc fragment complexed with a synthetic GT1b analogue suggests cross-linking between ganglioside receptors and the toxin. J. Biol. Chem. 2001, 276, 32274–32281. [Google Scholar] [CrossRef]
- Chen, C.; Baldwin, M.R.; Barbieri, J.T. Molecular basis for tetanus toxin coreceptor interactions. Biochemistry 2008, 47, 7179–7186. [Google Scholar] [CrossRef]
- Rummel, A.; Bade, S.; Alves, J.; Bigalke, H.; Binz, T. Two carbohydrate binding sites in the H(CC)-domain of tetanus neurotoxin are required for toxicity. J. Mol. Biol. 2003, 326, 835–847. [Google Scholar] [CrossRef]
- Schiavo, G.; Benfenati, F.; Poulain, B.; Rossetto, O.; Polverino de Laureto, P.; DasGupta, B.R.; Montecucco, C. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 1992, 359, 832–835. [Google Scholar] [CrossRef]
- Harper, C.B.; Martin, S.; Nguyen, T.H.; Daniels, S.J.; Lavidis, N.A.; Popoff, M.R.; Hadzic, G.; Mariana, A.; Chau, N.; McCluskey, A.; et al. Dynamin inhibition blocks botulinum neurotoxin type A endocytosis in neurons and delays botulism. J. Biol. Chem. 2011, 286, 35966–35976. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Masuyer, G.; Stenmark, P. Botulinum and Tetanus Neurotoxins. Annu. Rev. Biochem. 2019, 88, 811–837. [Google Scholar] [CrossRef]
- Winner, B.M.; Bodt, S.M.L.; McNutt, P.M. Special Delivery: Potential Mechanisms of Botulinum Neurotoxin Uptake and Trafficking within Motor Nerve Terminals. Int. J. Mol. Sci. 2020, 21, 8715. [Google Scholar] [CrossRef]
- Edupuganti, O.P.; Ovsepian, S.V.; Wang, J.; Zurawski, T.H.; Schmidt, J.J.; Smith, L.; Lawrence, G.W.; Dolly, J.O. Targeted delivery into motor nerve terminals of inhibitors for SNARE-cleaving proteases via liposomes coupled to an atoxic botulinum neurotoxin. FEBS J. 2012, 279, 2555–2567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yu, R.; Chi, X.; Chen, Z.; Hao, M.; Du, P.; Fan, P.; Liu, Y.; Dong, Y.; Fang, T.; et al. Tetanus vaccine-induced human neutralizing antibodies provide full protection against neurotoxin challenge in mice. Int. Immunopharmacol. 2021, 91, 107297. [Google Scholar] [CrossRef]
- Thwaites, C.L.; Beeching, N.J.; Newton, C.R. Maternal and neonatal tetanus. Lancet 2015, 385, 362–370. [Google Scholar] [CrossRef]
- Alhaji, M.A.; Mustapha, M.G.; Ashir, G.M.; Akuhwa, R.T.; Bello, M.A.; Farouk, A.G. Recurrent generalized tetanus: A case report. Trop. Dr. 2011, 41, 127–128. [Google Scholar] [CrossRef]
- Rhee, P.; Nunley, M.K.; Demetriades, D.; Velmahos, G.; Doucet, J.J. Tetanus and trauma: A review and recommendations. J. Trauma. 2005, 58, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Saylor, C.; Dadachova, E.; Casadevall, A. Monoclonal antibody-based therapies for microbial diseases. Vaccine 2009, 27 (Suppl. 6), G38–G46. [Google Scholar] [CrossRef]
- Alonso-Gutiérrez, A.; Díaz-Ramos, P.; Sulleiro-Avendaño, E.; de Miguel-Marañón, M.; Padilla-Gallego, M.E.; Sancho-López, A.; Ruiz-Antúnez, S.; Prieto-Yerro, C. Contribution of the spanish agency for medicines and healthcare products to the European committee for the evaluation of medicinal products for human use. Rev. Clin. Esp. 2015, 215, 230–235. [Google Scholar] [CrossRef]
- Ahmad, A.; Qaisar, I.; Naeem, M.; Mazhar, A.U.; Ashfaq, M. Intrathecal anti-tetanus human immunoglobulin in the treatment of neonatal tetanus. J. Coll. Physicians Surg. Pak. 2011, 21, 539–541. [Google Scholar] [PubMed]
- Gibson, R.B.; Banzhaf, E.J. The Quantitative Changes in the Proteins in the Blood Plasma of Horses in the Course of Immunization. J. Exp. Med. 1910, 12, 411–434. [Google Scholar] [CrossRef] [PubMed]
- de Smit, H.; Ackerschott, B.; Tierney, R.; Stickings, P.; Harmsen, M.M. A novel single-domain antibody multimer that potently neutralizes tetanus neurotoxin. Vaccine X 2021, 8, 100099. [Google Scholar] [CrossRef]
- Jin, B.K.; Odongo, S.; Radwanska, M.; Magez, S. NANOBODIES®: A Review of Diagnostic and Therapeutic Applications. Int. J. Mol. Sci. 2023, 24, 5994. [Google Scholar] [CrossRef]
- Zavrtanik, U.; Lukan, J.; Loris, R.; Lah, J.; Hadži, S. Structural Basis of Epitope Recognition by Heavy-Chain Camelid Antibodies. J. Mol. Biol. 2018, 430, 4369–4386. [Google Scholar] [CrossRef] [PubMed]
- Steeland, S.; Vandenbroucke, R.E.; Libert, C. Nanobodies as therapeutics: Big opportunities for small antibodies. Drug Discov. Today 2016, 21, 1076–1113. [Google Scholar] [CrossRef]
- Ye, G.; Gallant, J.; Zheng, J.; Massey, C.; Shi, K.; Tai, W.; Odle, A.; Vickers, M.; Shang, J.; Wan, Y.; et al. The development of Nanosota-1 as anti-SARS-CoV-2 nanobody drug candidates. eLife 2021, 10, e64815. [Google Scholar] [CrossRef]
- Liu, X.Y.; Wei, D.K.; Li, Z.Y.; Lu, J.S.; Xie, X.M.; Yu, Y.Z.; Pang, X.B. Immunogenicity and immunoprotection of the functional TL-HN fragment derived from tetanus toxin. Vaccine 2023, 41, 6834–6841. [Google Scholar] [CrossRef]
- Liu, F.J.; Shi, D.Y.; Li, Z.Y.; Lu, J.S.; Wang, R.; Pang, X.B.; Yang, Z.X.; Yu, Y.Z. Evaluation of a recombinant tetanus toxin subunit vaccine. Toxicon 2020, 187, 75–81. [Google Scholar] [CrossRef]
- Liu, F.J.; Shi, D.Y.; Mao, Y.Y.; Xiong, X.H.; Lu, J.S.; Pang, X.B.; Dong, X.J.; Yang, Z.X.; Yu, Y.Z. Immunological characterisation and immunoprotective efficacy of functional domain antigens of botulinum neurotoxin serotype A. Vaccine 2020, 38, 2978–2983. [Google Scholar] [CrossRef]
- Li, Z.; Lu, J.S.; Liu, S.; Wang, R.; Xu, Q.; Yu, Y.Z.; Yang, Z.X. Recombinant L-HN Fusion Antigen Derived from the L and HN Domains of Botulinum Neurotoxin B Stimulates a Protective Antibody Response Against Active Neurotoxin. Neurotox. Res. 2021, 39, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, J.; Tan, X.; Wang, R.; Xu, Q.; Yu, Y.; Yang, Z. Functional EL-HN Fragment as a Potent Candidate Vaccine for the Prevention of Botulinum Neurotoxin Serotype E. Toxins 2022, 14, 135. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, B.; Lu, J.; Liu, X.; Tan, X.; Wang, R.; Du, P.; Yu, S.; Xu, Q.; Pang, X.; et al. Biological and Immunological Characterization of a Functional L-HN Derivative of Botulinum Neurotoxin Serotype F. Toxins 2023, 15, 200. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Ji, C.; Xu, J.; Wang, D.; Fang, T.; Jing, Y.; Kwang-Fu Shen, C.; Chen, W. The Immunogenicity of the C Fragment of Tetanus Neurotoxin in Production of Tetanus Antitoxin. Biomed. Res. Int. 2018, 2018, 6057348. [Google Scholar] [CrossRef]
- Cheng, K.; Lu, J.; Guo, J.; Wang, R.; Chen, L.; Wang, X.; Jiang, Y.; Li, Y.; Xu, C.; Kang, Q.; et al. Characterization of neutralizing chimeric heavy-chain antibodies against tetanus toxin. Hum. Vaccin. Immunother. 2024, 20, 2366641. [Google Scholar] [CrossRef]
- Pirazzini, M.; Grinzato, A.; Corti, D.; Barbieri, S.; Leka, O.; Vallese, F.; Tonellato, M.; Silacci-Fregni, C.; Piccoli, L.; Kandiah, E.; et al. Exceptionally potent human monoclonal antibodies are effective for prophylaxis and treatment of tetanus in mice. J. Clin. Investig. 2021, 131, e151676. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Yu, J.; Lin, S.; Liu, T.; Zan, L.; Li, N.; Hong, P.; Wang, X.; Jia, Z.; et al. Structural basis of tetanus toxin neutralization by native human monoclonal antibodies. Cell Rep. 2021, 35, 109070. [Google Scholar] [CrossRef]
- Wang, H.; Yu, R.; Fang, T.; Yu, T.; Chi, X.; Zhang, X.; Liu, S.; Fu, L.; Yu, C.; Chen, W. Tetanus Neurotoxin Neutralizing Antibodies Screened from a Human Immune scFv Antibody Phage Display Library. Toxins 2016, 8, 266. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Cui, J.; Liu, D.; Zhang, W.; Xue, C.; Xiong, X.; Liu, G.; Chen, H. Preparation and characterization of a neutralizing murine monoclonal antibody against tetanus toxin. J. Immunol. Methods 2023, 513, 113427. [Google Scholar] [CrossRef]
- Lang, A.B.; Cryz, S.J., Jr.; Schürch, U.; Ganss, M.T.; Bruderer, U. Immunotherapy with human monoclonal antibodies. Fragment A specificity of polyclonal and monoclonal antibodies is crucial for full protection against tetanus toxin. J. Immunol. 1993, 151, 466–472. [Google Scholar] [CrossRef]
- Volk, W.A.; Bizzini, B.; Snyder, R.M.; Bernhard, E.; Wagner, R.R. Neutralization of tetanus toxin by distinct monoclonal antibodies binding to multiple epitopes on the toxin molecule. Infect. Immun. 1984, 45, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Kamei, M.; Sugimoto, N.; Ma, Y.; Hashizume, S. Characteristics of toxin-neutralization by anti-tetanus human monoclonal antibodies directed against the three functional domains [A], [B] and [C] of the tetanus toxin molecule and a reliable method for evaluating the protective effects of monoclonal antibodies. Eur. J. Epidemiol. 1992, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aliprandini, E.; Takata, D.Y.; Lepique, A.; Kalil, J.; Boscardin, S.B.; Moro, A.M. An oligoclonal combination of human monoclonal antibodies able to neutralize tetanus toxin in vivo. Toxicon X 2019, 2, 100006. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Montal, M. Molecular dissection of botulinum neurotoxin reveals interdomain chaperone function. Toxicon 2013, 75, 101–107. [Google Scholar] [CrossRef]
- Lukić, I.; Marinković, E.; Filipović, A.; Krnjaja, O.; Kosanović, D.; Inić-Kanada, A.; Stojanović, M. Key protection factors against tetanus: Anti-tetanus toxin antibody affinity and its ability to prevent tetanus toxin—Ganglioside interaction. Toxicon 2015, 103, 135–144. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Panning Round | Input | Output | Output/Input Ratio | Enrichment Factor |
|---|---|---|---|---|
| First | 5.0 × 1011 | 4.2 × 107 | 8.4 × 10−5 | - |
| Second | 1.0 × 1011 | 1.95 × 107 | 1.95 × 10−4 | 2.3 |
| Group a | Dose of TeNT b | Surviving/Total Mice c | Survival (%) | Group a | Dose of TeNT b | Surviving/Total Mice c | Survival (%) |
|---|---|---|---|---|---|---|---|
| TL-1-hFc | 10 × LD50 | 0/4 | 0 | TL-19-hFc | 10 × LD50 | 0/4 | 0 |
| TL-2-hFc | 10 × LD50 | 0/4 | 0 | TL-20-hFc | 10 × LD50 | 0/4 | 0 |
| TL-3-hFc | 10 × LD50 | 0/4 | 0 | TL-21-hFc | 10 × LD50 | 2/4 | 50 |
| TL-4-hFc | 10 × LD50 | 0/4 | 0 | TL-22-hFc | 10 × LD50 | 0/4 | 0 |
| TL-5-hFc | 10 × LD50 | 0/4 | 0 | TL-23-hFc | 10 × LD50 | 0/4 | 0 |
| TL-6-hFc | 10 × LD50 | 0/4 | 0 | TL-24-hFc | 10 × LD50 | 0/4 | 0 |
| TL-7-hFc | 10 × LD50 | 0/4 | 0 | TL-25-hFc | 10 × LD50 | 4/4 | 100 |
| TL-8-hFc | 10 × LD50 | 0/4 | 0 | TL-26-hFc | 10 × LD50 | 0/4 | 0 |
| TL-9-hFc | 10 × LD50 | 0/4 | 0 | TL-27-hFc | 10 × LD50 | 0/4 | 0 |
| TL-10-hFc | 10 × LD50 | 0/4 | 0 | TL-28-hFc | 10 × LD50 | 0/4 | 0 |
| TL-11-hFc | 10 × LD50 | 0/4 | 0 | TL-29-hFc | 10 × LD50 | 0/4 | 0 |
| TL-12-hFc | 10 × LD50 | 2/4 | 50 | TL-30-hFc | 10 × LD50 | 3/4 | 75 |
| TL-13-hFc | 10 × LD50 | 0/4 | 0 | TL-31-hFc | 10 × LD50 | 1/4 | 25 |
| TL-14-hFc | 10 × LD50 | 0/4 | 0 | TL-32-hFc | 10 × LD50 | 2/4 | 50 |
| TL-15-hFc | 10 × LD50 | 0/4 | 0 | TL-33-hFc | 10 × LD50 | 0/4 | 0 |
| TL-16-hFc | 10 × LD50 | 4/4 | 100 | TL-34-hFc | 10 × LD50 | 4/4 | 100 |
| TL-17-hFc | 10 × LD50 | 0/4 | 0 | TL-35-hFc | 10 × LD50 | 0/4 | 0 |
| TL-18-hFc | 10 × LD50 | 0/4 | 0 | TL-36-hFc | 10 × LD50 | 0/4 | 0 |
| Dilution buffer | 10 × LD50 | 0/4 | 0 |
| Protein | Kon (104 Ms−1) | Kdis (10−4 s−1) | KD (nM) | χ2 | R2 |
|---|---|---|---|---|---|
| TL-16-hFc | 2.38 | 2.39 | 10.1 | 0.579 | 0.99 |
| TL-25-hFc | 2.59 | 4.44 | 17.1 | 0.0198 | 0.99 |
| TL-34-hFc | 4.66 | 1.68 | 3.61 | 0.6906 | 0.99 |
| Protein | Kon (104 Ms−1) | Kdis (10−4 s−1) | KD (nM) | χ2 | R2 |
|---|---|---|---|---|---|
| TL-16-h1-hFc | 2.239 | 0.2036 | 0.9094 | 0.2084 | 0.99 |
| TL-25-h1-hFc | 1.89 | 0.109 | 0.573 | 0.5898 | 0.99 |
| TL-34-h1-hFc | 3.98 | 0.252 | 0.633 | 0.6744 | 0.99 |
| Group | Dose of Protein a (μg) | Dose of TeNT b | Surviving/Total Mice c | Survival (%) |
|---|---|---|---|---|
| TL-16-h1-hFc + TL-25-h1-hFc | 0.156 + 0.156 | 10 × LD50 | 4/4 | 100 |
| 0.078 + 0.078 | 10 × LD50 | 4/4 | 100 | |
| 0.039 + 0.039 | 10 × LD50 | 3/4 | 75 | |
| 0.0195 + 0.0195 | 10 × LD50 | 0/4 | 0 | |
| TL-16-h1-hFc + TL-34-h1-hFc | 0.156 + 0.312 | 10 × LD50 | 4/4 | 100 |
| 0.078 + 0.156 | 10 × LD50 | 4/4 | 100 | |
| 0.039 + 0.078 | 10 × LD50 | 4/4 | 100 | |
| 0.0195 + 0.039 | 10 × LD50 | 2/4 | 50 | |
| TL-25-h1-hFc + TL-34-h1-hFc | 0.156 + 0.312 | 10 × LD50 | 4/4 | 100 |
| 0.078 + 0.156 | 10 × LD50 | 4/4 | 100 | |
| 0.039 + 0.078 | 10 × LD50 | 4/4 | 100 | |
| 0.0195 + 0.039 | 10 × LD50 | 3/4 | 75 | |
| TIG | 0.1 IU | 10 × LD50 | 4/4 | 100 |
| Dilution buffer | - | 10 × LD50 | 0/4 | 0 |
| Antibody a | Dose of Protein b | Dose of TeNT c | Number of Survivors/Total Mice per Group | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 12 h d | 24 h | 48 h | 3 d | 5 d | 7 d | 9 d | 12 d | 14 d | |||
| TL-16-h1-hFc | 25 μg/kg | 10 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 2/4 ** | 0/4 ** | 0/4 ** | 0/4 ** | 0/4 ** | 0/4 ** |
| 125 μg/kg | 10 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 3/4 ** | |
| TL-25-h1-hFc | 25 μg/kg | 10 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 3/4 ** | 1/4 ** | 1/4 ** | 0/4 ** | 0/4 ** |
| 125 μg/kg | 10 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 3/4 ** | |
| TL-34-h1-hFc | 50 μg/kg | 10 × LD50 | 4/4 ** | 4/4 ** | 3/4 ** | 2/4 ** | 0/4 ** | 0/4 ** | 0/4 ** | 0/4 * | 0/4 ** |
| 250 μg/kg | 10 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 2/4 ** | 0/4 ** | 0/4 ** | |
| TIG | 0.1 IU | 10 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** |
| B-h3 | 250 μg/kg | 10 × LD50 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 |
| PBS | - | 10 × LD50 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 |
| Antibody a | Dose of Protein b | Dose of TeNT c | Number of Survivors/Total Mice per Group | ||||
|---|---|---|---|---|---|---|---|
| 1 h d | 3 h | 6 h | 12 h | 24 h | |||
| TL-16-h1-hFc | 25 μg/kg | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** |
| 125 μg/kg | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | |
| TL-25-h1-hFc | 25 μg/kg | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** |
| 125 μg/kg | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | |
| TL-34-h1-hFc | 50 μg/kg | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** |
| 250 μg/kg | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | |
| TIG | 0.1 IU | 5 × LD50 | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** | 4/4 ** |
| B-h3 | 250 μg/kg | 5 × LD50 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 |
| PBS | - | 5 × LD50 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Cheng, K.; Guo, J.; Jiang, Y.; Kang, Q.; Wang, R.; Du, P.; Gao, C.; Yu, Y.; Yang, Z.; et al. Humanized VHH-hFc Fusion Proteins Targeting the L-HN Fragment of Tetanus Toxin Provided Protection In Vivo. Antibodies 2025, 14, 48. https://doi.org/10.3390/antib14020048
Li Y, Cheng K, Guo J, Jiang Y, Kang Q, Wang R, Du P, Gao C, Yu Y, Yang Z, et al. Humanized VHH-hFc Fusion Proteins Targeting the L-HN Fragment of Tetanus Toxin Provided Protection In Vivo. Antibodies. 2025; 14(2):48. https://doi.org/10.3390/antib14020048
Chicago/Turabian StyleLi, Yating, Kexuan Cheng, Jiazheng Guo, Yujia Jiang, Qinglin Kang, Rong Wang, Peng Du, Chen Gao, Yunzhou Yu, Zhixin Yang, and et al. 2025. "Humanized VHH-hFc Fusion Proteins Targeting the L-HN Fragment of Tetanus Toxin Provided Protection In Vivo" Antibodies 14, no. 2: 48. https://doi.org/10.3390/antib14020048
APA StyleLi, Y., Cheng, K., Guo, J., Jiang, Y., Kang, Q., Wang, R., Du, P., Gao, C., Yu, Y., Yang, Z., Wang, W., & Lu, J. (2025). Humanized VHH-hFc Fusion Proteins Targeting the L-HN Fragment of Tetanus Toxin Provided Protection In Vivo. Antibodies, 14(2), 48. https://doi.org/10.3390/antib14020048