
Expanding the Anti-Phl p 7 Antibody Toolkit: An Anti-Idiotype Nanobody Inhibitor

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Development of Anti-IgD Nanobodies
2.3. Expression and Purification of aδNb072
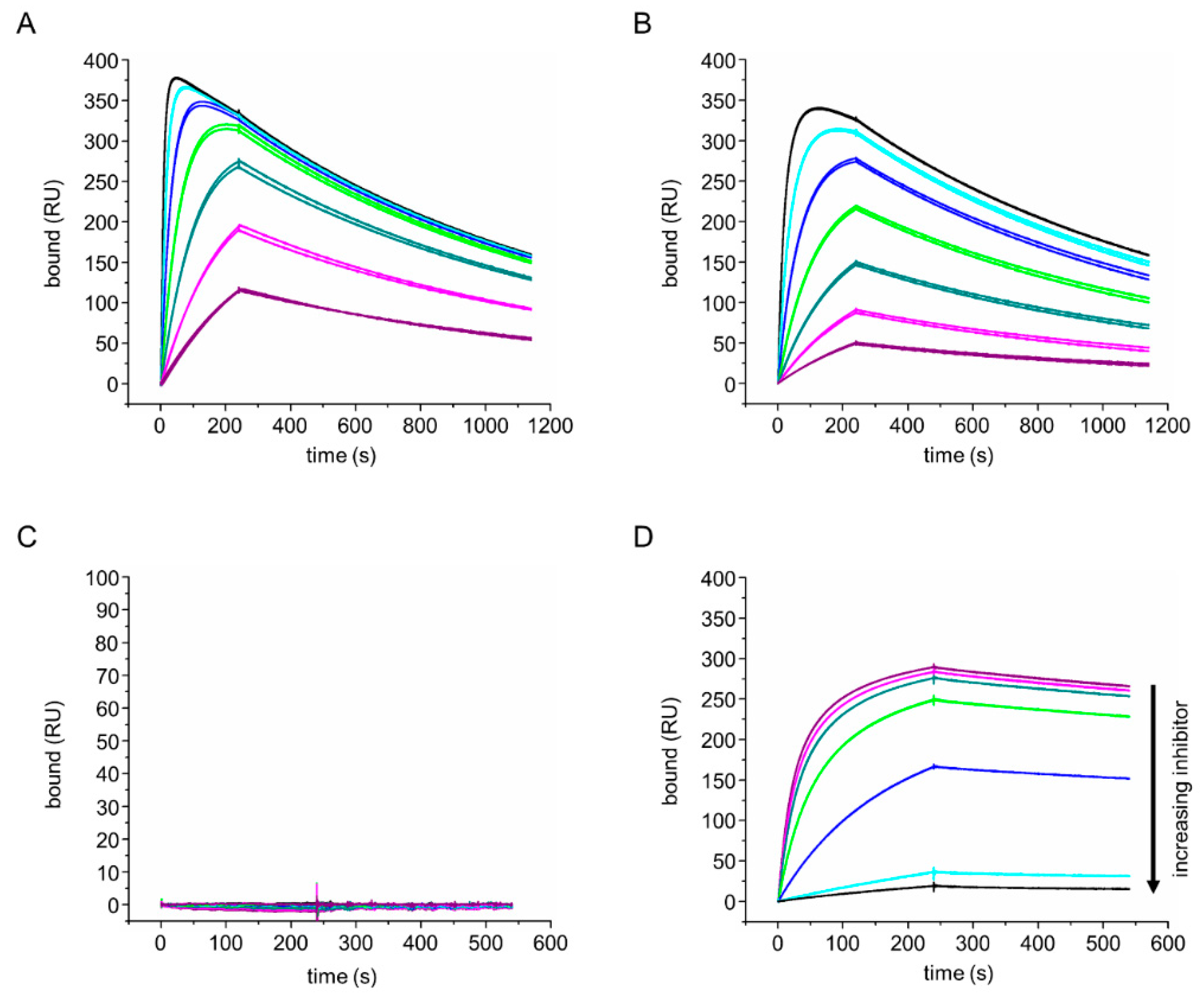
2.4. Surface Plasmon Resonance
2.5. aδNb072/HAPPID1 Fab Complex Purification and Crystallization
2.6. X-ray Structure Determination and Refinement
3. Results
3.1. Production of an Anti-Phl p 7 IgD Antibody
3.2. Identification and Characterization of an Anti-Paratope Nb
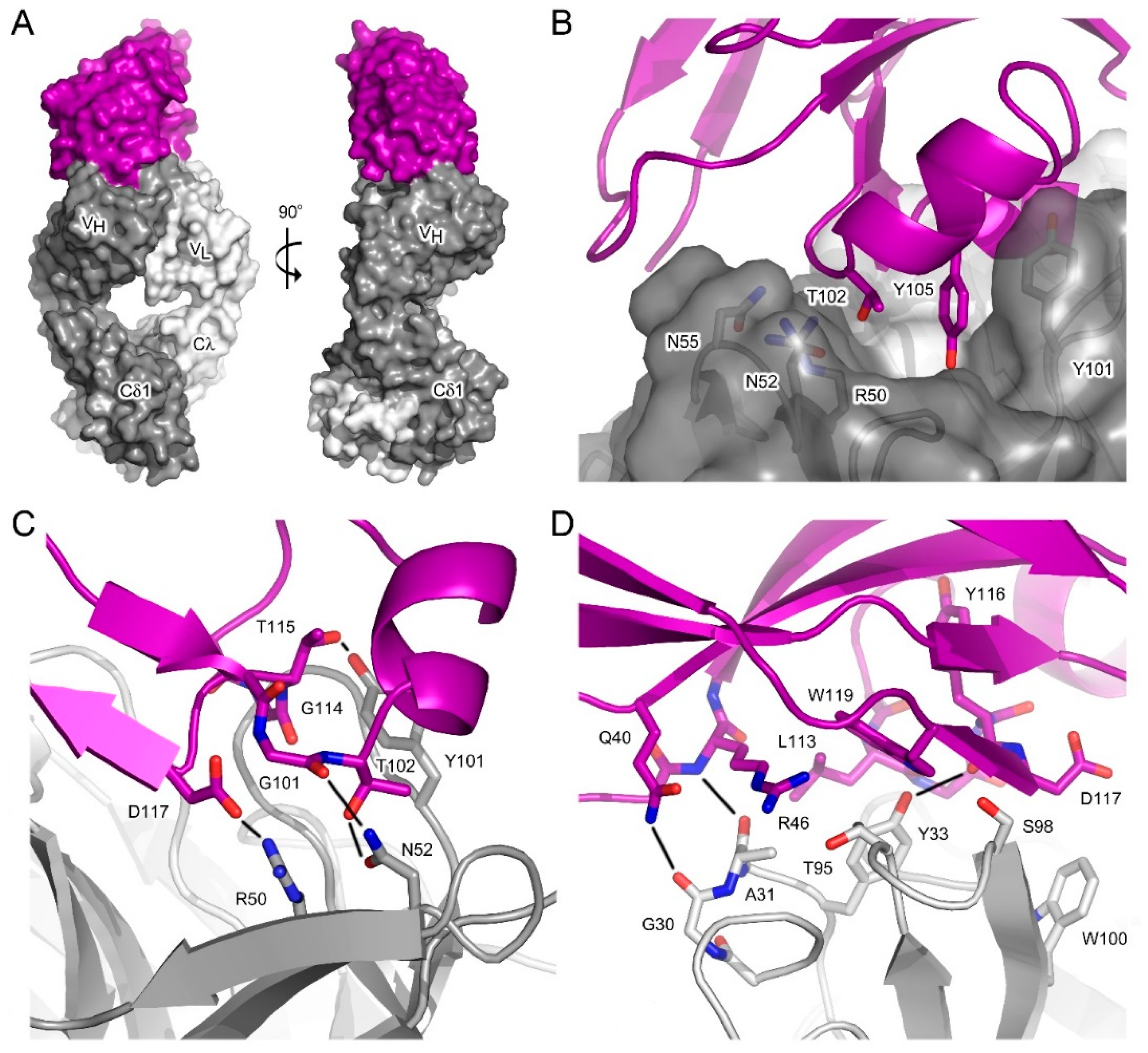
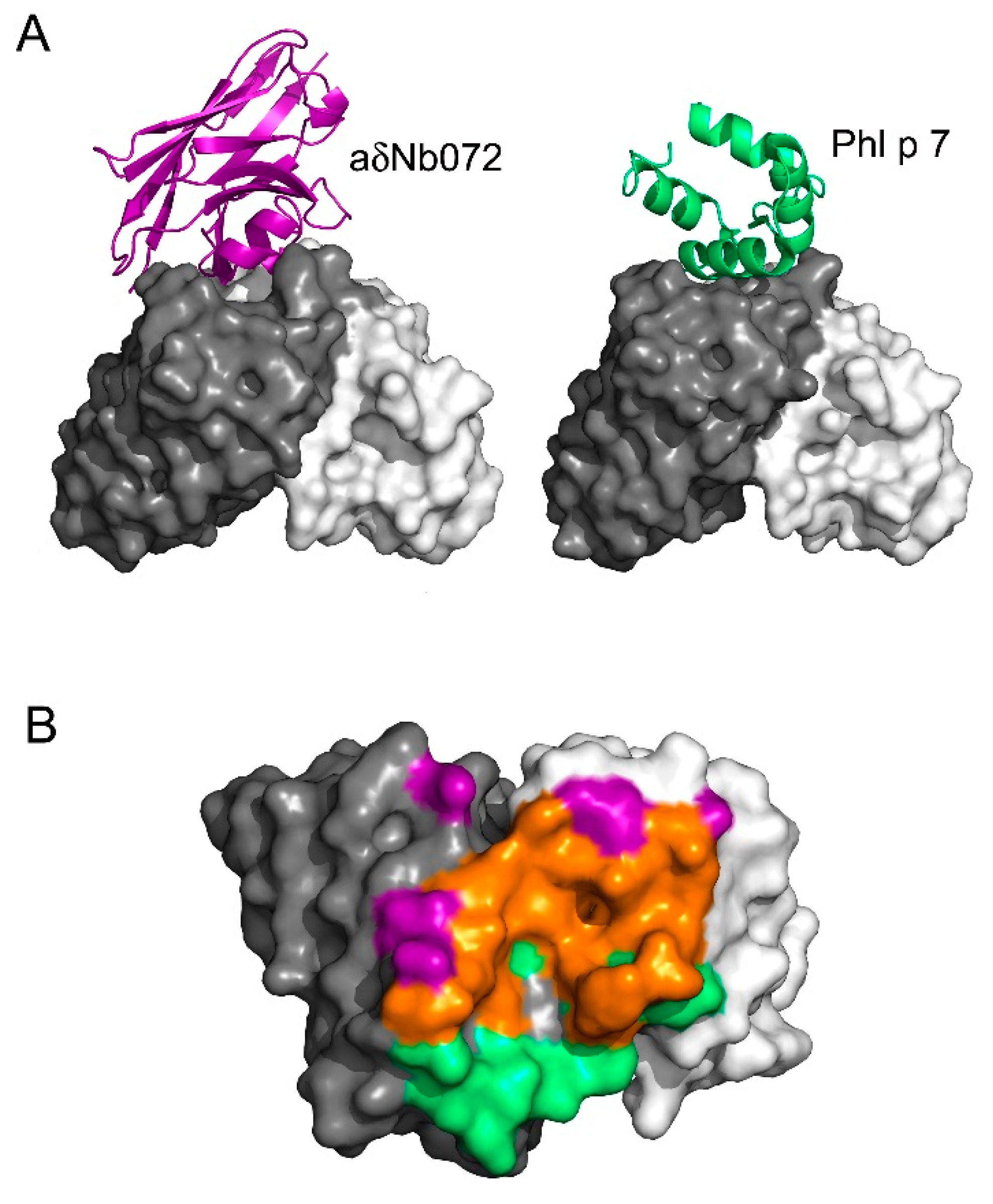
3.3. Interface between aδNb072 and the HAPPID1 Fab
3.4. aδNb072 Is an Orthosteric Inhibitor of Phl p 7
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballow, M. Primary Immunodeficiency Disorders: Antibody Deficiency. J. Allergy Clin. Immunol. 2002, 109, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Pisetsky, D.S. Pathogenesis of Autoimmune Disease. Nat. Rev. Nephrol. 2023, 19, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J.; Sutton, B.J.; Beavil, A.J.; Beavil, R.L.; McCloskey, N.; Coker, H.A.; Fear, D.; Smurthwaite, L. The Biology of IgE and the Basis of Allergic Disease. Annu. Rev. Immunol. 2003, 21, 579–628. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.W.; Cavacini, L. Structure and Function of Immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef]
- Lu, L.L.; Suscovich, T.J.; Fortune, S.M.; Alter, G. Beyond Binding: Antibody Effector Functions in Infectious Diseases. Nat. Rev. Immunol. 2018, 18, 46–61. [Google Scholar] [CrossRef]
- Sela-Culang, I.; Kunik, V.; Ofran, Y. The Structural Basis of Antibody-Antigen Recognition. Front. Immunol. 2013, 4, 302. [Google Scholar] [CrossRef]
- Niederberger, V.; Hayek, B.; Vrtala, S.; Laffer, S.; Twardosz, A.; Vangelista, L.; Sperr, W.R.; Valent, P.; Rumpold, H.; Kraft, D.; et al. Calcium-dependent Immunoglobulin E Recognition of the Apo- and Calcium-bound Form of a Cross-reactive Two EF-hand Timothy Grass Pollen Allergen, Phl p 7. FASEB J. 1999, 13, 843–856. [Google Scholar] [CrossRef]
- James, L.K.; Bowen, H.; Calvert, R.A.; Dodev, T.S.; Shamji, M.H.; Beavil, A.J.; McDonnell, J.M.; Durham, S.R.; Gould, H.J. Allergen Specificity of IgG4-Expressing B Cells in Patients with Grass Pollen Allergy Undergoing Immunotherapy. J. Allergy Clin. Immunol. 2012, 130, 663–670. [Google Scholar] [CrossRef]
- Dodev, T.S.; Karagiannis, P.; Gilbert, A.E.; Josephs, D.H.; Bowen, H.; James, L.K.; Bax, H.J.; Beavil, R.; Pang, M.O.; Gould, H.J.; et al. A Tool Kit for Rapid Cloning and Expression of Recombinant Antibodies. Sci. Rep. 2014, 4, 5885. [Google Scholar] [CrossRef]
- Dodev, T.S.; Bowen, H.; Shamji, M.H.; Bax, H.J.; Beavil, A.J.; McDonnell, J.M.; Durham, S.R.; Sutton, B.J.; Gould, H.J.; James, L.K. Inhibition of Allergen-Dependent IgE Activity by Antibodies of the Same Specificity but Different Class. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 720–724. [Google Scholar] [CrossRef]
- Mitropoulou, A.N.; Bowen, H.; Dodev, T.S.; Davies, A.M.; Bax, H.J.; Beavil, R.L.; Beavil, A.J.; Gould, H.J.; James, L.K.; Sutton, B.J. Structure of a Patient-Derived Antibody in Complex with Allergen Reveals Simultaneous Conventional and Superantigen-like Recognition. Proc. Natl. Acad. Sci. USA 2018, 115, E8707–E8716. [Google Scholar] [CrossRef] [PubMed]
- Bucaite, G.; Kang-Pettinger, T.; Moreira, J.; Gould, H.J.; James, L.K.; Sutton, B.J.; McDonnell, J.M. Interplay between Affinity and Valency in Effector Cell Degranulation: A Model System with Polcalcin Allergens and Human Patient–Derived IgE Antibodies. J. Immunol. 2019, 203, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Rowe, D.S.; Fahey, J.L. A New Class of Human Immunoglobulins. II. Normal Serum IgD. J. Exp. Med. 1965, 121, 185–199. [Google Scholar] [CrossRef]
- Gutzeit, C.; Chen, K.; Cerutti, A. The Enigmatic Function of IgD: Some Answers at Last. Eur. J. Immunol. 2018, 48, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Ohshima, Y. The Dual Aspects of IgD in the Development of Tolerance and the Pathogenesis of Allergic Diseases. Allergol. Int. 2022, 72, 227–233. [Google Scholar] [CrossRef]
- Davies, A.M.; Beavil, R.L.; Barbolov, M.; Sandhar, B.S.; Gould, H.J.; Beavil, A.J.; Sutton, B.J.; McDonnell, J.M. Crystal Structures of the Human IgD Fab Reveal Insights into CH1 Domain Diversity. Mol. Immunol. 2023, 159, 28–37. [Google Scholar] [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally Occurring Antibodies Devoid of Light Chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef]
- Ghahroudi, M.A.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. Selection and Identification of Single Domain Antibody Fragments from Camel Heavy-Chain Antibodies. FEBS Lett. 1997, 414, 521–526. [Google Scholar] [CrossRef]
- Muyldermans, S. Applications of Nanobodies. Annu. Rev. Anim. Biosci. 2021, 9, 401–421. [Google Scholar] [CrossRef]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide Tag Forming a Rapid Covalent Bond to a Protein, through Engineering a Bacterial Adhesin. Proc. Natl. Acad. Sci. USA 2012, 109, E690–E697. [Google Scholar] [CrossRef]
- Vester, S.K.; Rahikainen, R.; Khairil Anuar, I.N.A.; Hills, R.A.; Tan, T.K.; Howarth, M. SpySwitch Enables pH- or Heat-Responsive Capture and Release for Plug-and-Display Nanoassembly. Nat. Commun. 2022, 13, 3714. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W. Protein Production by Auto-Induction in High-Density Shaking Cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Pardon, E.; Laeremans, T.; Triest, S.; Rasmussen, S.G.F.; Wohlkönig, A.; Ruf, A.; Muyldermans, S.; Hol, W.G.J.; Kobilka, B.K.; Steyaert, J. A General Protocol for the Generation of Nanobodies for Structural Biology. Nat. Protoc. 2014, 9, 674–693. [Google Scholar] [CrossRef] [PubMed]
- Schoonaert, L.; Rué, L.; Roucourt, B.; Timmers, M.; Little, S.; Chávez-Gutiérrez, L.; Dewilde, M.; Joyce, P.; Curnock, A.; Weber, P.; et al. Identification and Characterization of Nanobodies Targeting the EphA4 Receptor. J. Biol. Chem. 2017, 292, 11452–11465. [Google Scholar] [CrossRef]
- Muyldermans, S. A Guide to: Generation and Design of Nanobodies. FEBS J. 2021, 288, 2084–2102. [Google Scholar] [CrossRef]
- Myszka, D.G. Improving Biosensor Analysis. J. Mol. Recognit. 1999, 12, 279–284. [Google Scholar] [CrossRef]
- Hulme, E.C.; Trevethick, M.A. Ligand Binding Assays at Equilibrium: Validation and Interpretation. Br. J. Pharmacol. 2010, 161, 1219–1237. [Google Scholar] [CrossRef]
- Winter, G.; Waterman, D.G.; Parkhurst, J.M.; Brewster, A.S.; Gildea, R.J.; Gerstel, M.; Fuentes-Montero, L.; Vollmar, M.; Michels-Clark, T.; Young, I.D.; et al. DIALS: Implementation and Evaluation of a New Integration Package. Acta Cryst. D 2018, D74, 85–97. [Google Scholar] [CrossRef]
- Agirre, J.; Atanasova, M.; Bagdonas, H.; Ballard, C.B.; Baslé, A.; Beilsten-Edmands, J.; Borges, R.J.; Brown, D.G.; Burgos-Mármol, J.J.; Berrisford, J.M.; et al. The CCP4 Suite: Integrative Software for Macromolecular Crystallography. Acta Cryst. D 2023, D79, 449–461. [Google Scholar] [CrossRef]
- Evans, P.R.; Murshudov, G.N. How Good Are My Data and What Is the Resolution? Acta Cryst. D 2013, D69, 1204–1214. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser Crystallographic Software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Sosa, B.A.; Demircioglu, F.E.; Chen, J.Z.; Ingram, J.; Ploegh, H.L.; Schwartz, T.U. How Lamina-Associated Polypeptide 1 (LAP1) Activates Torsin. eLife 2014, 3, e03239. [Google Scholar] [CrossRef] [PubMed]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkóczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.-W.; Jain, S.; McCoy, A.J.; et al. Macromolecular Structure Determination Using X-Rays, Neutrons and Electrons: Recent Developments in Phenix. Acta Cryst. D 2019, D75, 861–877. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Cryst. D 2010, D66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- McDonnell, J.M.; Dhaliwal, B.; Sutton, B.J.; Gould, H.J. IgE, IgE Receptors and Anti-IgE Biologics: Protein Structures and Mechanisms of Action. Annu. Rev. Immunol. 2023, 41, 255–275. [Google Scholar] [CrossRef]
- MacCallum, R.M.; Martin, A.C.R.; Thornton, J.M. Antibody-Antigen Interactions: Contact Analysis and Binding Site Topography. J. Mol. Biol. 1996, 262, 732–745. [Google Scholar] [CrossRef]
- De Genst, E.; Silence, K.; Decanniere, K.; Conrath, K.; Loris, R.; Kinne, J.; Muyldermans, S.; Wyns, L. Molecular Basis for the Preferential Cleft Recognition by Dromedary Heavy-Chain Antibodies. Proc. Natl. Acad. Sci. USA 2006, 103, 4586–4591. [Google Scholar] [CrossRef]
- Zavrtanik, U.; Lukan, J.; Loris, R.; Lah, J.; Hadži, S. Structural Basis of Epitope Recognition by Heavy-Chain Camelid Antibodies. J. Mol. Biol. 2018, 430, 4369–4386. [Google Scholar] [CrossRef]
- Selzer, T.; Schreiber, G. New Insights into the Mechanism of Protein-Protein Association. Proteins Struct. Funct. Genet. 2001, 45, 190–198. [Google Scholar] [CrossRef]
- Janda, A.; Bowen, A.; Greenspan, N.S.; Casadevall, A. Ig Constant Region Effects on Variable Region Structure and Function. Front. Microbiol. 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Kohler, H.; Pashov, A.; Kieber-Emmons, T. The Promise of Anti-Idiotype Revisited. Front. Immunol. 2019, 10, 808. [Google Scholar] [CrossRef] [PubMed]
- Sutton, B.J.; Gould, H.J. The Human IgE Network. Nature 1993, 366, 421–428. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| aδNb072/HAPPID1 Fab | |
|---|---|
| Data Processing | |
| Space group | P 21 21 21 |
| a, b, c (Å) | 65.74, 71.04, 122.17 |
| Resolution (Å) a | 61.09–2.10 (2.16–2.10) |
| Completeness (%) a | 100.0 (100.0) |
| Multiplicitya | 26.4 (26.7) |
| Mean (I)/σ (I)a | 9.5 (1.2) |
| CC1/2 a | 0.998 (0.611) |
| Rpim (%) a | 6.2 (65.5) |
| Wilson B factor (Å2) | 35.23 |
| Refinement | |
| Rwork/Rfree (%) b | 21.42/25.92 |
| No. of reflections | 34 098 |
| RMSD | |
| Bond lengths (Å) | 0.002 |
| Bond angles (°) | 0.555 |
| Coordinate error (Å) | 0.26 |
| No. of atoms | |
| Protein | 4 281 c |
| Solvent | 148 |
| Other | 49 d |
| Average B factor (Å2) | |
| Protein | 49.15 |
| Solvent | 42.97 |
| Other | 52.98 |
| Ramachandran plot | |
| Favored (%) | 97.51 |
| Allowed (%) | 2.49 |
| HAPPID1 Fab | HAPPIE1 Fab | |
|---|---|---|
| KD ± SD (M) | 1.40 (±0.01) × 10−9 | 3.64 (±0.08) × 10−9 |
| kon ± SD (M−1 s−1) | 5.86 (±0.01) × 105 | 2.28 (±0.01) × 105 |
| koff ± SD (s−1) | 8.22 (±0.08) × 10−4 | 8.29 (±0.20) × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vester, S.K.; Davies, A.M.; Beavil, R.L.; Sandhar, B.S.; Beavil, A.J.; Gould, H.J.; Sutton, B.J.; McDonnell, J.M. Expanding the Anti-Phl p 7 Antibody Toolkit: An Anti-Idiotype Nanobody Inhibitor. Antibodies 2023, 12, 75. https://doi.org/10.3390/antib12040075
Vester SK, Davies AM, Beavil RL, Sandhar BS, Beavil AJ, Gould HJ, Sutton BJ, McDonnell JM. Expanding the Anti-Phl p 7 Antibody Toolkit: An Anti-Idiotype Nanobody Inhibitor. Antibodies. 2023; 12(4):75. https://doi.org/10.3390/antib12040075
Chicago/Turabian StyleVester, Susan K., Anna M. Davies, Rebecca L. Beavil, Balraj S. Sandhar, Andrew J. Beavil, Hannah J. Gould, Brian J. Sutton, and James M. McDonnell. 2023. "Expanding the Anti-Phl p 7 Antibody Toolkit: An Anti-Idiotype Nanobody Inhibitor" Antibodies 12, no. 4: 75. https://doi.org/10.3390/antib12040075
APA StyleVester, S. K., Davies, A. M., Beavil, R. L., Sandhar, B. S., Beavil, A. J., Gould, H. J., Sutton, B. J., & McDonnell, J. M. (2023). Expanding the Anti-Phl p 7 Antibody Toolkit: An Anti-Idiotype Nanobody Inhibitor. Antibodies, 12(4), 75. https://doi.org/10.3390/antib12040075