
An Antibody Targeting Fibroblast Activation Protein Simultaneously Fused to Interleukin-2 and Tumor Necrosis Factor Selectively Localizes to Neoplastic Lesions

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Cloning, Expression, and Protein Purification
2.3. Biochemical Protein Characterization
2.4. Affinity Measurements
2.5. In Vitro Biological Activities
2.6. Flow Cytometry
2.7. Immunofluorescence Studies
2.8. Mice and Tumor Models
2.9. Immunofluorescence-Based Biodistribution
2.10. Mice Therapy Studies
2.11. Statistical Analysis
2.12. Ethical Statement
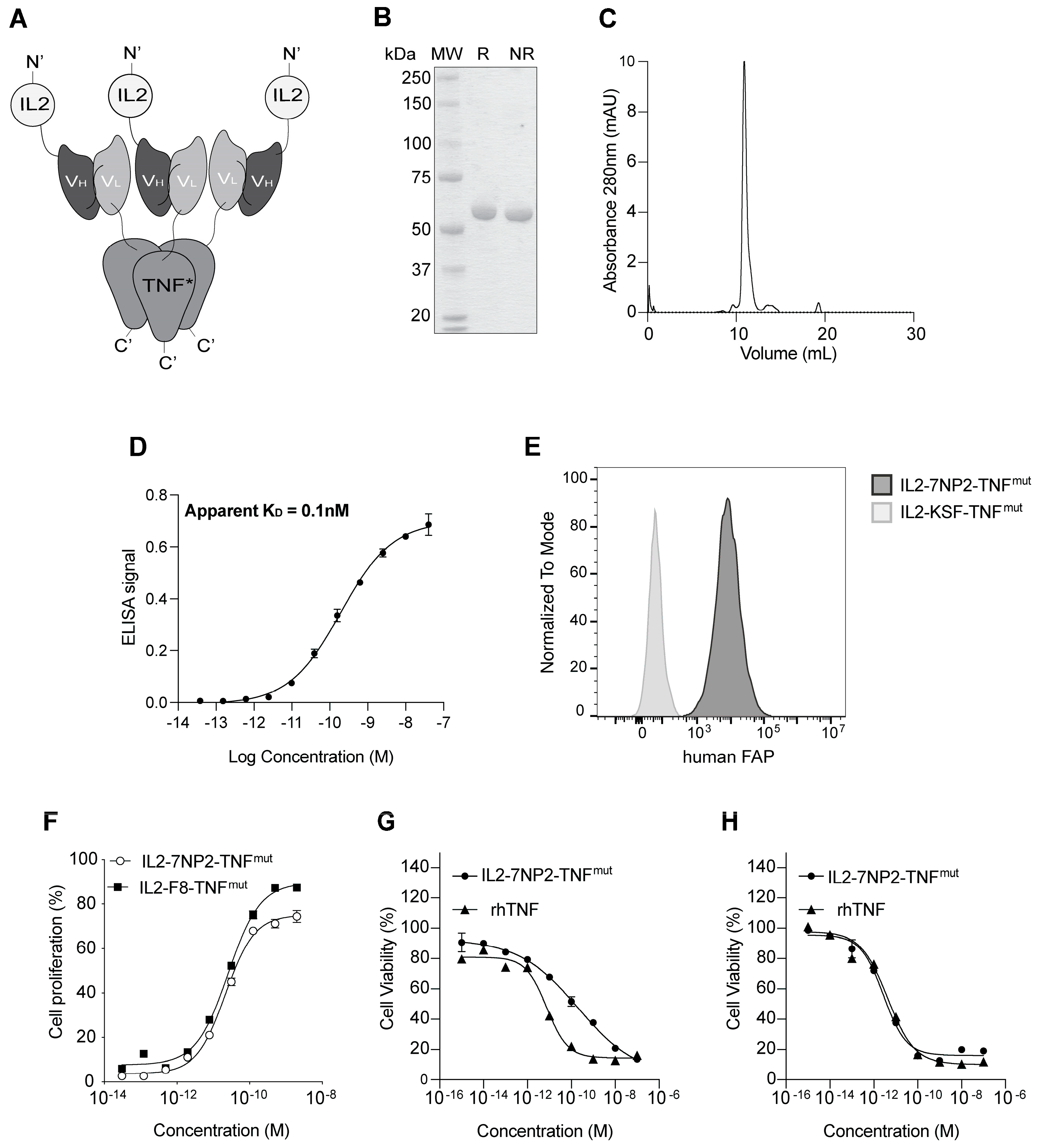
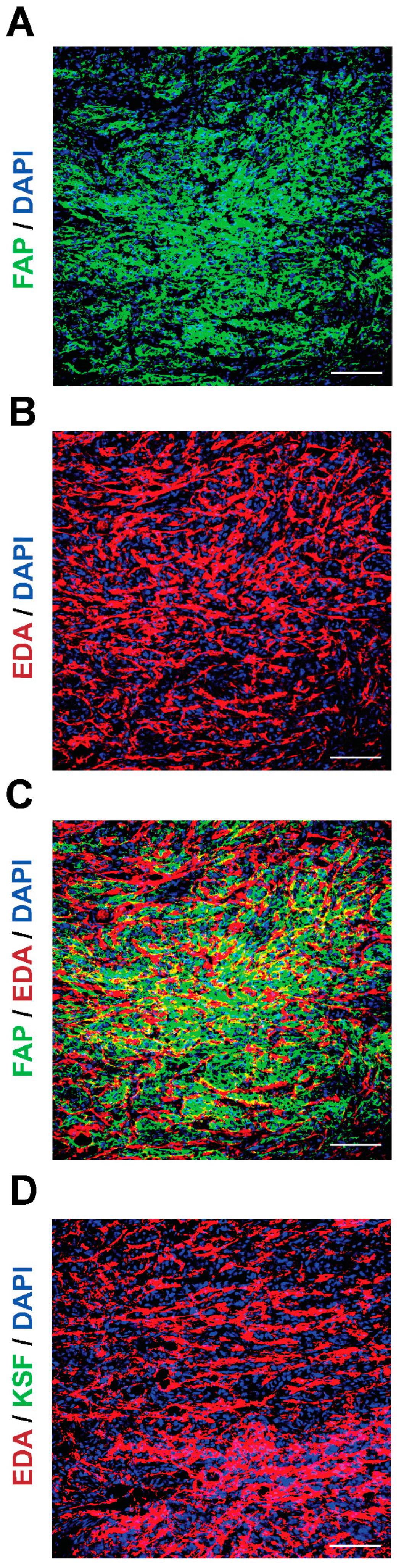
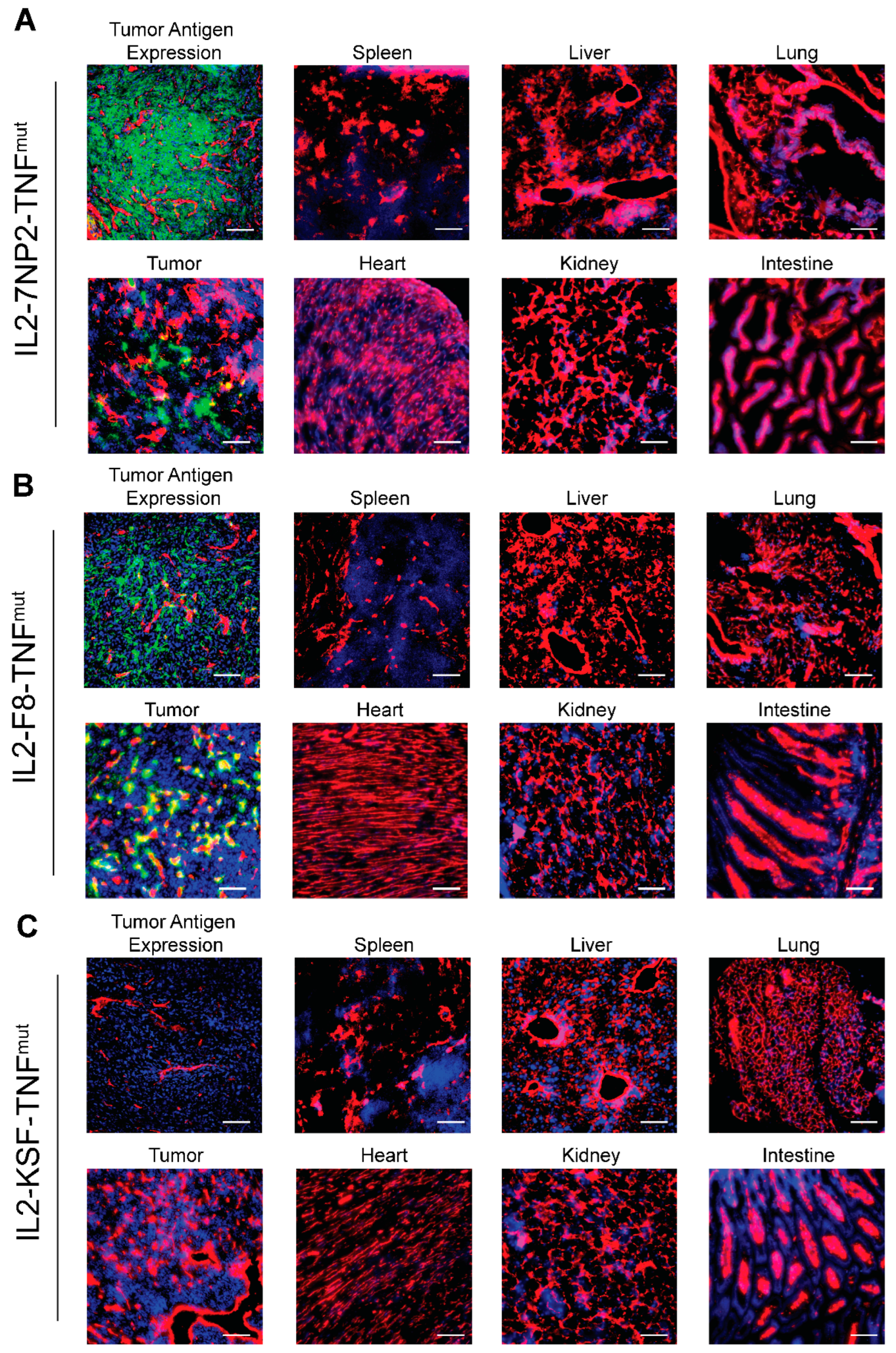
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neri, D. Antibody–Cytokine Fusions: Versatile Products for the Modulation of Anticancer Immunity. Cancer Immunol. Res. 2019, 7, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.A. IL-2: The First Effective Immunotherapy for Human Cancer. J. Immunol. 2014, 192, 5451–5458. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Kerner, Z.J.; Hong, H.; Sun, J. Targeted Cancer Therapy with Tumor Necrosis Factor-Alpha. Biochem. Insights 2008, 1, 15–21. [Google Scholar] [CrossRef]
- Conlon, K.C.; Miljkovic, M.D.; Waldmann, T.A. Cytokines in the Treatment of Cancer. J. Interferon Cytokine Res. 2019, 39, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Price, G.; Brenner, M.; Prentice, H.; Hoffbrand, A.; Newland, A. Cytotoxic effects of tumour necrosis factor and gamma-interferon on acute myeloid leukaemia blasts. Br. J. Cancer 1987, 55, 287–290. [Google Scholar] [CrossRef]
- Fyfe, G.; Fisher, R.I.; Rosenberg, S.A.; Sznol, M.; Parkinson, D.R.; Louie, A.C. Results of treatment of 255 patients with metastatic renal cell carcinoma who received high-dose recombinant interleukin-2 therapy. J. Clin. Oncol. 1995, 13, 688–696. [Google Scholar] [CrossRef]
- Atkins, M.B.; Lotze, M.T.; Dutcher, J.P.; Fisher, R.I.; Weiss, G.; Margolin, K.; Abrams, J.; Sznol, M.; Parkinson, D.; Hawkins, M.; et al. High-Dose Recombinant Interleukin 2 Therapy for Patients With Metastatic Melanoma: Analysis of 270 Patients Treated Between 1985 and 1993. J. Clin. Oncol. 1999, 17, 2105–2116. [Google Scholar] [CrossRef]
- Dutcher, J.P.; Schwartzentruber, D.J.; Kaufman, H.L.; Agarwala, S.S.; Tarhini, A.; Lowder, J.N.; Atkins, M.B. High dose interleukin-2 (Aldesleukin)—Expert consensus on best management practices-2014. J. Immunother. Cancer 2014, 2, 26. [Google Scholar] [CrossRef]
- List, T.; Neri, D. Immunocytokines: A review of molecules in clinical development for cancer therapy. Clin. Pharmacol. Adv. Appl. 2013, 5 (Suppl. S1), 29–45. [Google Scholar] [CrossRef]
- Gutbrodt, K.L.; Neri, D. Immunocytokines. Antibodies 2012, 1, 70–87. [Google Scholar] [CrossRef]
- Runbeck, E.; Crescioli, S.; Karagiannis, S.; Papa, S. Utilizing Immunocytokines for Cancer Therapy. Antibodies 2021, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wu, Y.; Bi, J.; Huang, Y.; Cheng, Y.; Li, Y.; Wu, Y.; Cao, G.; Tian, Z. The use of supercytokines, immunocytokines, engager cytokines, and other synthetic cytokines in immunotherapy. Cell. Mol. Immunol. 2022, 19, 192–209. [Google Scholar] [CrossRef] [PubMed]
- Gout, D.Y.; Groen, L.S.; van Egmond, M. The present and future of immunocytokines for cancer treatment. Cell. Mol. Life Sci. 2022, 79, 509. [Google Scholar] [CrossRef] [PubMed]
- Bootz, F.; Neri, D. Immunocytokines: A novel class of products for the treatment of chronic inflammation and autoimmune conditions. Drug Discov. Today 2016, 21, 180–189. [Google Scholar] [CrossRef]
- Hutmacher, C.; Neri, D. Antibody-cytokine fusion proteins: Biopharmaceuticals with immunomodulatory properties for cancer therapy. Adv. Drug Deliv. Rev. 2018, 141, 67–91. [Google Scholar] [CrossRef]
- Sondel, P.M.; Gillies, S.D. Immunocytokines for Cancer Immunotherapy. In Handbook of Cancer Vaccines; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004; pp. 341–358. [Google Scholar] [CrossRef]
- Tak, W.M.; Mary, E.S.; Bradley, D.J. (Eds.) Tumor Immunology. In Primer to the Immune Response, 2nd ed.; Elsevier Science: Amsterdam, The Netherlands, 2014; pp. 423–455. [Google Scholar] [CrossRef]
- Murphy, K.; Weaver, C. Janeway’s Immunology, 9th ed.; Garland Science/Taylor & Francis Group, LLC.: New York, NY, USA, 2017. [Google Scholar]
- Balza, E.; Carnemolla, B.; Mortara, L.; Castellani, P.; Soncini, D.; Accolla, R.S.; Borsi, L. Therapy-induced antitumor vaccination in neuroblastomas by the combined targeting of IL-2 and TNFα. Int. J. Cancer 2009, 127, 101–110. [Google Scholar] [CrossRef]
- Danielli, R.; Patuzzo, R.; Di Giacomo, A.M.; Gallino, G.; Maurichi, A.; Di Florio, A.; Cutaia, O.; Lazzeri, A.; Fazio, C.; Miracco, C.; et al. Intralesional administration of L19-IL2/L19-TNF in stage III or stage IVM1a melanoma patients: Results of a phase II study. Cancer Immunol. Immunother. 2015, 64, 999–1009. [Google Scholar] [CrossRef]
- Schwager, K.; Hemmerle, T.; Aebischer, D.; Neri, D. The Immunocytokine L19–IL2 Eradicates Cancer When Used in Combination with CTLA-4 Blockade or with L19-TNF. J. Investig. Dermatol. 2013, 133, 751–758. [Google Scholar] [CrossRef]
- Gillies, S.D.; Lan, Y.; Brunkhorst, B.; Wong, W.-K.; Li, Y.; Lo, K.-M. Bi-functional cytokine fusion proteins for gene therapy and antibody-targeted treatment of cancer. Cancer Immunol. Immunother. 2002, 51, 449–460. [Google Scholar] [CrossRef]
- Kermer, V.; Hornig, N.; Harder, M.; Bondarieva, A.; Kontermann, R.E.; Müller, D. Combining Antibody-Directed Presentation of IL-15 and 4-1BBL in a Trifunctional Fusion Protein for Cancer Immunotherapy. Mol. Cancer Ther. 2014, 13, 112–121. [Google Scholar] [CrossRef]
- Jahn, T.; Zuther, M.; Friedrichs, B.; Heuser, C.; Guhlke, S.; Abken, H.; Hombach, A.A. An Il12-Il2-Antibody Fusion Protein Targeting Hodgkin’s Lymphoma Cells Potentiates Activation Of Nk And T Cells For An Anti-Tumor Attack. PLoS ONE 2012, 7, e44482. [Google Scholar] [CrossRef] [PubMed]
- De Luca, R.; Kachel, P.; Kropivsek, K.; Snijder, B.; Manz, M.G.; Neri, D. A novel dual-cytokine–antibody fusion protein for the treatment of CD38-positive malignancies. Protein Eng. Des. Sel. 2018, 31, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Schanzer, J.M.; Fichtner, I.; Baeuerle, P.A.; Kufer, P. Antitumor Activity of a Dual Cytokine/Single-chain Antibody Fusion Protein for Simultaneous Delivery of GM-CSF and IL-2 to Ep-CAM Expressing Tumor Cells. J. Immunother. 2006, 29, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Helguera, G.; Rodríguez, J.A.; Penichet, M.L. Cytokines fused to antibodies and their combinations as therapeutic agents against different peritoneal HER2/neu expressing tumors. Mol. Cancer Ther. 2006, 5, 1029–1040. [Google Scholar] [CrossRef]
- Halin, C.; Gafner, V.; Villani, M.E.; Borsi, L.; Berndt, A.; Kosmehl, H.; Zardi, L.; Neri, D. Synergistic Therapeutic Effects of a Tumor Targeting Antibody Fragment, Fused to Interleukin 12 and to Tumor Necrosis Factor alpha. Cancer Res. 2003, 63, 3202–3210. [Google Scholar]
- Borsi, L.; Balza, E.; Carnemolla, B.; Sassi, F.; Castellani, P.; Berndt, A.; Kosmehl, H.; Birò, A.; Siri, A.; Orecchia, P.; et al. Selective targeted delivery of TNFα to tumor blood vessels. Blood 2003, 102, 4384–4392. [Google Scholar] [CrossRef]
- Hemmerle, T.; Probst, P.; Giovannoni, L.; Green, A.J.; Meyer, T.; Neri, D. The antibody-based targeted delivery of TNF in combination with doxorubicin eradicates sarcomas in mice and confers protective immunity. Br. J. Cancer 2013, 109, 1206–1213. [Google Scholar] [CrossRef]
- Puca, E.; Probst, P.; Stringhini, M.; Murer, P.; Pellegrini, G.; Cazzamalli, S.; Hutmacher, C.; Gouyou, B.; Wulhfard, S.; Matasci, M.; et al. The antibody-based delivery of interleukin-12 to solid tumors boosts NK and CD8 + T cell activity and synergizes with immune checkpoint inhibitors. Int. J. Cancer 2019, 146, 2518–2530. [Google Scholar] [CrossRef]
- Ziffels, B.; Stringhini, M.; Probst, P.; Fugmann, T.; Sturm, T.; Neri, D. Antibody-Based Delivery of Cytokine Payloads to Carbonic Anhydrase IX Leads to Cancer Cures in Immunocompetent Tumor-Bearing Mice. Mol. Cancer Ther. 2019, 18, 1544–1554. [Google Scholar] [CrossRef]
- De Luca, R.; Soltermann, A.; Pretto, F.; Pemberton-Ross, C.; Pellegrini, G.; Wulhfard, S.; Neri, D. Potency-matched Dual Cytokine–Antibody Fusion Proteins for Cancer Therapy. Mol. Cancer Ther. 2017, 16, 2442–2451. [Google Scholar] [CrossRef]
- De Luca, R.; Neri, D. Potentiation of PD-L1 blockade with a potency-matched dual cytokine–antibody fusion protein leads to cancer eradication in BALB/c-derived tumors but not in other mouse strains. Cancer Immunol. Immunother. 2018, 67, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- Sedo, A.; Busek, P.; Mateu, R.; Zubal, M.; Kotackova, L. Targeting fibroblast activation protein in cancer ndash Prospects and caveats. Front. Biosci. 2018, 23, 1933–1968. [Google Scholar] [CrossRef]
- Hamson, E.J.; Keane, F.M.; Tholen, S.; Schilling, O.; Gorrell, M.D. Understanding fibroblast activation protein (FAP): Substrates, activities, expression and targeting for cancer therapy. Proteom. Clin. Appl. 2014, 8, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Wang, S.; Wang, H.; Tang, W. Fibroblast activation protein-α in tumor cells promotes colorectal cancer angiogenesis via the Akt and ERK signaling pathways. Mol. Med. Rep. 2017, 17, 2593–2599. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-F.; Zhang, L.-H.; Shan, L.-H.; Sun, W.-G.; Chai, C.-C.; Wu, H.-M.; Ibla, J.C.; Wang, L.-F.; Liu, J.-R. Effects of the fibroblast activation protein on the invasion and migration of gastric cancer. Exp. Mol. Pathol. 2013, 95, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Martin, T.A.; Ye, L.; Jiang, W.G. FAP-α (Fibroblast activation protein-α) is involved in the control of human breast cancer cell line growth and motility via the FAK pathway. BMC Cell Biol. 2014, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.A.; Schubert, R.D.; Peter, R.U.; Kraut, N.; Park, J.E.; Rettig, W.J.; Garin-Chesa, P. Fibroblast Activation Protein: Differential Expression and Serine Protease Activity in Reactive Stromal Fibroblasts of Melanocytic Skin Tumors. J. Investig. Dermatol. 2003, 120, 182–188. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, L.; Xia, Z.-W.; Yang, S.-C.; Li, W.-H.; Liu, B.; Yu, Z.-Q.; Gong, P.-F.; Yang, Y.-L.; Sun, W.-Z.; et al. The role of fibroblast activation protein in progression and development of osteosarcoma cells. Clin. Exp. Med. 2020, 20, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Puré, E.; Blomberg, R. Pro-tumorigenic roles of fibroblast activation protein in cancer: Back to the basics. Oncogene 2018, 37, 4343–4357. [Google Scholar] [CrossRef]
- Pang, Y.; Zhao, L.; Luo, Z.; Hao, B.; Wu, H.; Lin, Q.; Sun, L.; Chen, H. Comparison of 68Ga-FAPI and 18F-FDG Uptake in Gastric, Duodenal, and Colorectal Cancers. Radiology 2021, 298, 393–402. [Google Scholar] [CrossRef]
- Röhrich, M.; Naumann, P.; Giesel, F.L.; Choyke, P.L.; Staudinger, F.; Wefers, A.; Liew, D.P.; Kratochwil, C.; Rathke, H.; Liermann, J.; et al. Impact of 68Ga-FAPI PET/CT Imaging on the Therapeutic Management of Primary and Recurrent Pancreatic Ductal Adenocarcinomas. J. Nucl. Med. 2020, 62, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Nadal, L.; Peissert, F.; Elsayed, A.; Weiss, T.; Look, T.; Weller, M.; Piro, G.; Carbone, C.; Tortora, G.; Matasci, M.; et al. Generation and in vivo validation of an IL-12 fusion protein based on a novel anti-human FAP monoclonal antibody. J. Immunother. Cancer 2022, 10, e005282. [Google Scholar] [CrossRef] [PubMed]
- Millul, J.; Bassi, G.; Mock, J.; Elsayed, A.; Pellegrino, C.; Zana, A.; Plaza, S.D.; Nadal, L.; Gloger, A.; Schmidt, E.; et al. An ultra-high-affinity small organic ligand of fibroblast activation protein for tumor-targeting applications. Proc. Natl. Acad. Sci. USA 2021, 118, e2101852118. [Google Scholar] [CrossRef] [PubMed]
- De Luca, R.; Gouyou, B.; Ongaro, T.; Villa, A.; Ziffels, B.; Sannino, A.; Buttinoni, G.; Galeazzi, S.; Mazzacuva, M.; Neri, D. A Novel Fully-Human Potency-Matched Dual Cytokine-Antibody Fusion Protein Targets Carbonic Anhydrase IX in Renal Cell Carcinomas. Front. Oncol. 2019, 9, 1228. [Google Scholar] [CrossRef]
- Villa, A.; Trachsel, E.; Kaspar, M.; Schliemann, C.; Sommavilla, R.; Rybak, J.-N.; Rösli, C.; Borsi, L.; Neri, D. A high-affinity human monoclonal antibody specific to the alternatively spliced EDA domain of fibronectin efficiently targets tumor neo-vasculaturein vivo. Int. J. Cancer 2008, 122, 2405–2413. [Google Scholar] [CrossRef]
- Folli, S.; Épèlegrin, A.; Chalandon, Y.; Yao, X.; Buchegger, F.; Lienard, D.; Lejeune, F.; Mach, J.-P. Tumor-necrosis factor can enhance radio-antibody uptake in human colon carcinoma xenografts by increasing vascular permeability. Int. J. Cancer 1993, 53, 829–836. [Google Scholar] [CrossRef]
- Khawli, L.A.; Miller, G.K.; Epstein, A.L. Effect of seven new vasoactive immunoconjugates on the enhancement of monoclonal antibody uptake in tumors. Cancer 1994, 73, 824–831. [Google Scholar] [CrossRef]
- Watanabe, N.; Niitsu, Y.; Umeno, H.; Kuriyama, H.; Neda, H.; Yamauchi, N.; Maeda, M.; Urushizaki, I. Toxic effect of tumor necrosis factor on tumor vasculature in mice. Cancer Res 1988, 48, 2179–2183. [Google Scholar]
- Renard, N.; Liénard, D.; Lespagnard, L.; Eggermont, A.; Heimann, R.; Lejeune, F. Early endothelium activation and polymorphonuclear cell invasion precede specific necrosis of human melanoma and sarcoma treated by intravascular high-dose tumour necrosis factor alpha (rTNFα). Int. J. Cancer 1994, 57, 656–663. [Google Scholar] [CrossRef]
- Gregorc, V.; Cavina, R.; Novello, S.; Grossi, F.; Lazzari, C.; Capelletto, E.; Genova, C.; Salini, G.; Lambiase, A.; Santoro, A. NGR-hTNF and Doxorubicin as Second-Line Treatment of Patients with Small Cell Lung Cancer. Oncologist 2018, 23, 1133–e112. [Google Scholar] [CrossRef]
- Landgraf, W.; Sandow, J. Recombinant Human Insulins—Clinical Efficacy and Safety in Diabetes Therapy. Eur. Endocrinol. 2016, 12, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Loktev, A.; Lindner, T.; Mier, W.; Debus, J.; Altmann, A.; Jäger, D.; Giesel, F.; Kratochwil, C.; Barthe, P.; Roumestand, C.; et al. A Tumor-Imaging Method Targeting Cancer-Associated Fibroblasts. J. Nucl. Med. 2018, 59, 1423–1429. [Google Scholar] [CrossRef] [PubMed]
- Loktev, A.; Lindner, T.; Burger, E.-M.; Altmann, A.; Giesel, F.; Kratochwil, C.; Debus, J.; Marme, F.; Jäger, D.; Mier, W.; et al. Development of Fibroblast Activation Protein-Targeted Radiotracers with Improved Tumor Retention. J. Nucl. Med. 2019, 60, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Pang, Y.; Yao, L.; Zhao, L.; Fan, C.; Ke, J.; Guo, P.; Hao, B.; Fu, H.; Xie, C.; et al. Imaging fibroblast activation protein in liver cancer: A single-center post hoc retrospective analysis to compare [68Ga]Ga-FAPI-04 PET/CT versus MRI and [18F]-FDG PET/CT. Eur. J. Nucl. Med. 2020, 48, 1604–1617. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prodi, E.; Comacchio, C.; Gilardoni, E.; Di Nitto, C.; Puca, E.; Neri, D.; De Luca, R. An Antibody Targeting Fibroblast Activation Protein Simultaneously Fused to Interleukin-2 and Tumor Necrosis Factor Selectively Localizes to Neoplastic Lesions. Antibodies 2023, 12, 29. https://doi.org/10.3390/antib12020029
Prodi E, Comacchio C, Gilardoni E, Di Nitto C, Puca E, Neri D, De Luca R. An Antibody Targeting Fibroblast Activation Protein Simultaneously Fused to Interleukin-2 and Tumor Necrosis Factor Selectively Localizes to Neoplastic Lesions. Antibodies. 2023; 12(2):29. https://doi.org/10.3390/antib12020029
Chicago/Turabian StyleProdi, Eleonora, Claudia Comacchio, Ettore Gilardoni, Cesare Di Nitto, Emanuele Puca, Dario Neri, and Roberto De Luca. 2023. "An Antibody Targeting Fibroblast Activation Protein Simultaneously Fused to Interleukin-2 and Tumor Necrosis Factor Selectively Localizes to Neoplastic Lesions" Antibodies 12, no. 2: 29. https://doi.org/10.3390/antib12020029
APA StyleProdi, E., Comacchio, C., Gilardoni, E., Di Nitto, C., Puca, E., Neri, D., & De Luca, R. (2023). An Antibody Targeting Fibroblast Activation Protein Simultaneously Fused to Interleukin-2 and Tumor Necrosis Factor Selectively Localizes to Neoplastic Lesions. Antibodies, 12(2), 29. https://doi.org/10.3390/antib12020029