
A Review of the Currently Available Antibody Therapy for the Treatment of Coronavirus Disease 2019 (COVID-19)
Abstract
1. Introduction
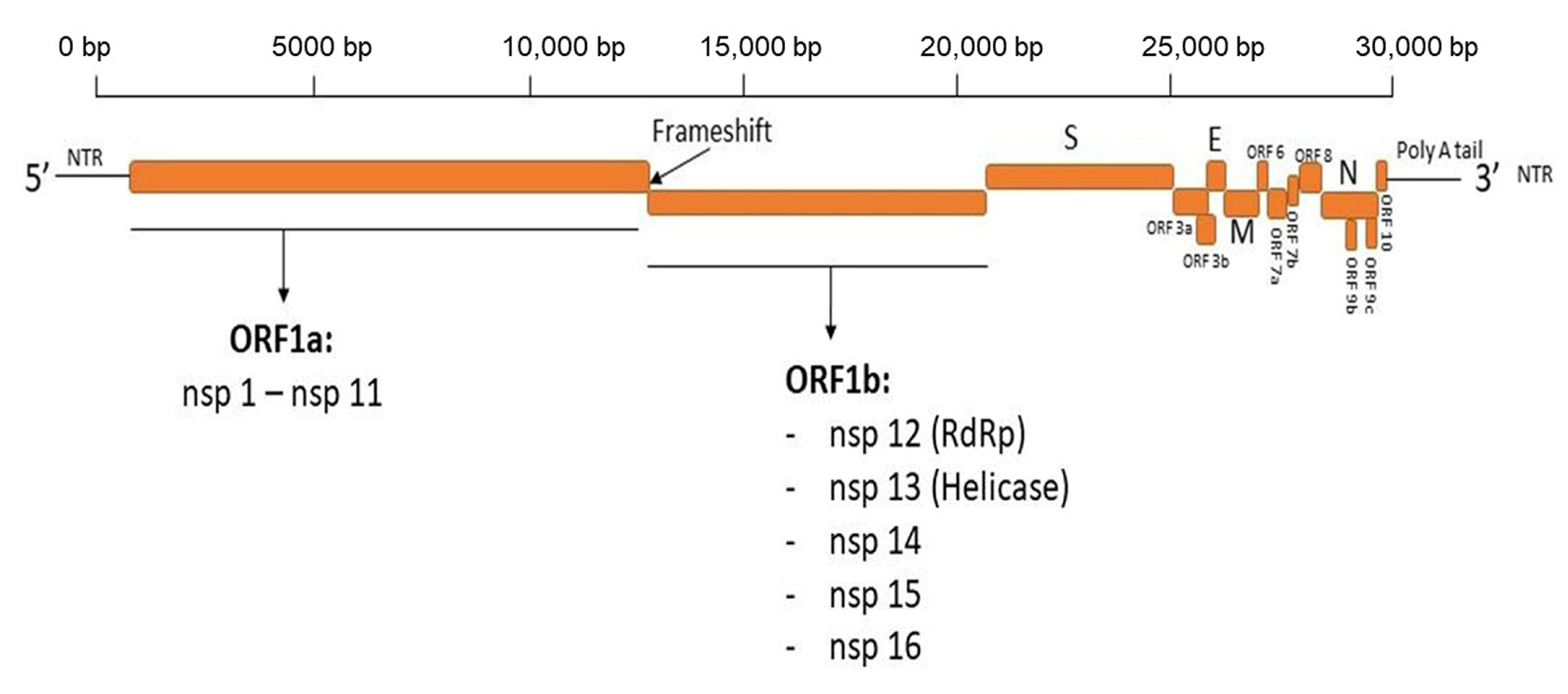
2. SARS-CoV-2
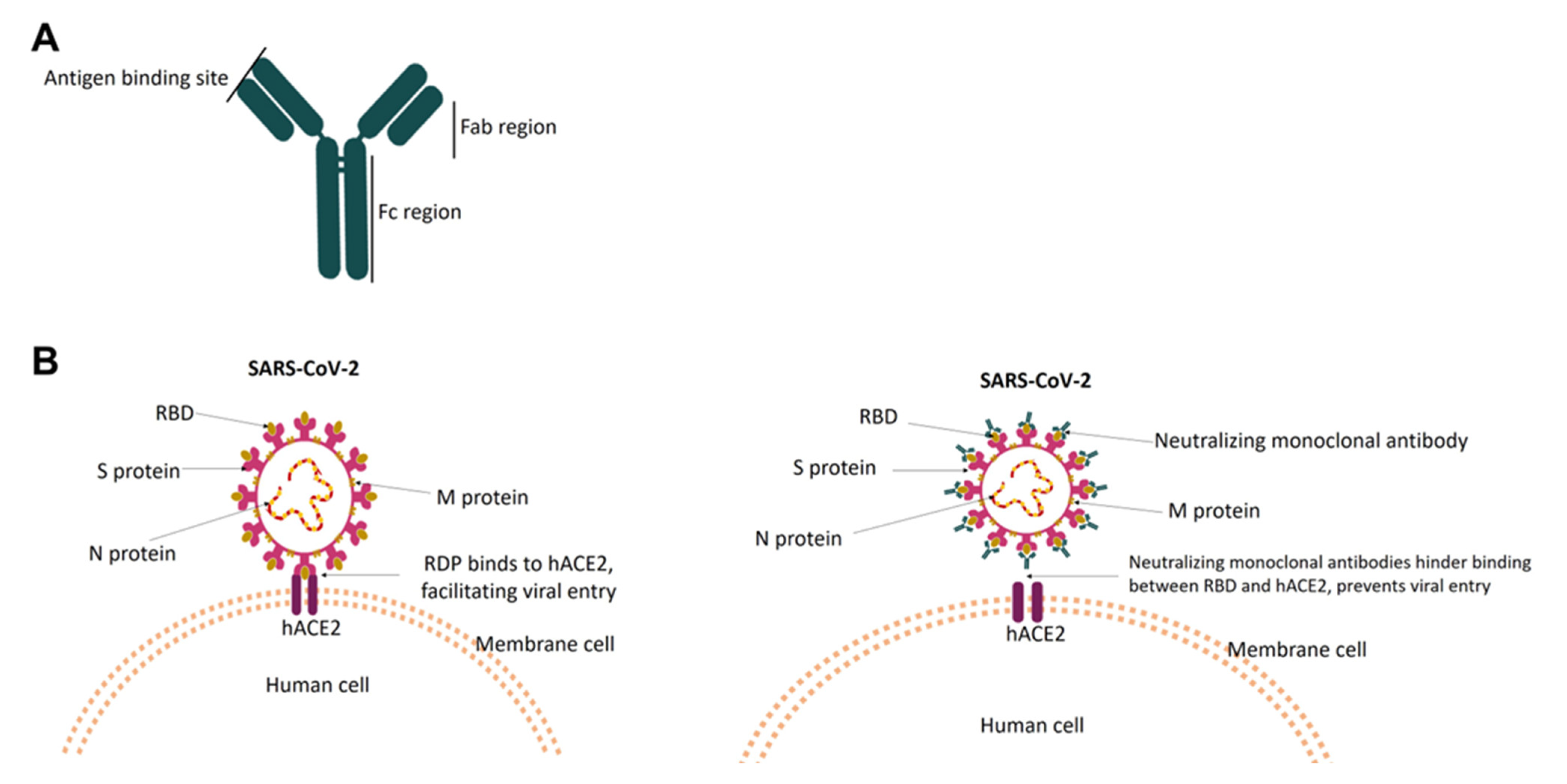
3. Neutralizing Monoclonal Antibodies against SARS-CoV-2
4. Currently Available Monoclonal Antibodies for COVID-19 Treatment
4.1. Evusheld (Combination of Tixagevimab and Cilgavimab)
4.2. Bebtelovimab
4.3. Bispecific Antibodies
4.4. Antibodies That Alleviate the Harmful Effect of an Over-Stimulated Host Immune Response
5. Challenges to the Use of Antibody Therapies for COVID-19
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kahn, J.S.; McIntosh, K. History and recent advances in coronavirus discovery. J. Pediatr. Infect. Dis. 2005, 24, S223–S227. [Google Scholar] [CrossRef]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and sources of endemic human coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [PubMed]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.; Demmler-Harrison, G.J.; Kaplan, S.L.; Steinbach, W.J.; Hotez, P.J. Feigin and Cherry’s Textbook of Pediatric Infectious Diseases E-Book: 2-Volume Set; Elsevier Health Sciences: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Brian, D.; Baric, R. Coronavirus genome structure, and replication. Curr. Top. Microbiol. Immunol. 2005, 287, 1–30. [Google Scholar] [PubMed]
- Nichols, G.L.; Gillingham, E.; Macintyre, H.; Vardoulakis, S.; Hajat, S.; Sarran, C.; Amankwaah, D.; Phalkey, R. Coronavirus seasonality, respiratory infections, and weather. BMC Infect. Dis. 2021, 21, 1–15. [Google Scholar] [CrossRef]
- UK Research and Innovation. Coronavirus Seasonality: Is the Spread Likely to Vary? Available online: https://coronavirusexplained.ukri.org/en/article/und0009/ (accessed on 10 November 2022).
- Audi, A.; AlIbrahim, M.; Kaddoura, M.; Hijazi, G.; Yassine, H.M.; Zaraket, H. Seasonality of respiratory viral infections: Will COVID-19 follow suit? Front. Public Health 2020, 8, 576. [Google Scholar] [CrossRef]
- Peiris, J.S.; Guan, Y.; Yuen, K. Severe acute respiratory syndrome. Nat. Med. 2004, 10, S88–S97. [Google Scholar] [CrossRef]
- Cowling, B.J.; Park, M.; Fang, V.J.; Wu, P.; Leung, G.M.; Wu, J.T. Preliminary epidemiological assessment of MERS-CoV outbreak in South Korea, May to June 2015. Euro Surveil. 2015, 20, 21163. [Google Scholar] [CrossRef]
- Singh, D.; Yi, S.V. On the origin and evolution of SARS-CoV-2. Exp. Mol. Med. 2021, 53, 537–547. [Google Scholar] [CrossRef]
- Vos, L.; Bruyndonckx, R.; Zuithoff, N.; Little, P.; Oosterheert, J.; Broekhuizen, B.; Lammens, C.; Loens, K.; Viveen, M.; Butler, C. Lower respiratory tract infection in the community: Associations between viral etiology and illness course. Clin. Microbiol. Infect. 2021, 27, 96–104. [Google Scholar] [CrossRef]
- Zhu, Z.; Lian, X.; Su, X.; Wu, W.; Marraro, G.A.; Zeng, Y. From SARS and MERS to COVID-19: A summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respir. Res. 2020, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Boudjelal, M.; Nehdi, A.; Islam, I. Why do SARS-COV vaccines not exist? The pharma scientific intelligence and business model must be revisited! Expert Opin. Drug Discov. 2020, 15, 1233–1235. [Google Scholar] [CrossRef] [PubMed]
- Shanmugaraj, B.; Siriwattananon, K.; Wangkanont, K.; Phoolcharoen, W. Perspectives on monoclonal antibody therapy as a potential therapeutic intervention for Coronavirus disease-19 (COVID-19). Asian Pac. J. Allergy Immunol. 2020, 38, 10–18. [Google Scholar]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Khailany, R.A.; Safdar, M.; Ozaslan, M. Genomic characterization of a novel SARS-CoV-2. Gene Rep. 2020, 19, 100682. [Google Scholar] [CrossRef]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: Structural genomics approach. Biochim. Biophys. Acta-Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef]
- Yoshimoto, F.K. The proteins of severe acute respiratory syndrome coronavirus-2 (SARS CoV-2 or n-COV19) cause COVID-19. Protein J. 2020, 39, 198–216. [Google Scholar] [CrossRef]
- Wu, C.-r.; Yin, W.-c.; Jiang, Y.; Xu, H.E. Structure genomics of SARS-CoV-2 and its Omicron variant: Drug design templates for COVID-19. Acta Pharmacol. Sin. 2022, 43, 3021–3033. [Google Scholar] [CrossRef]
- Thomas, S. Mapping the nonstructural transmembrane proteins of severe acute respiratory syndrome coronavirus 2. J. Comput. Biol. 2021, 28, 909–921. [Google Scholar] [CrossRef]
- Gupta, M.K.; Vemula, S.; Donde, R.; Gouda, G.; Behera, L.; Vadde, R. In-silico approaches to detect inhibitors of the human severe acute respiratory syndrome coronavirus envelope protein ion channel. J. Biomol. Struct. Dyn. 2021, 39, 2617–2627. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary identification of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef] [PubMed]
- Gil, C.; Ginex, T.; Maestro, I.; Nozal, V.; Barrado-Gil, L.; Cuesta-Geijo, M.Á.; Urquiza, J.; Ramírez, D.; Alonso, C.; Campillo, N.E. COVID-19: Drug targets and potential treatments. J. Med. Chem. 2020, 63, 12359–12386. [Google Scholar] [CrossRef] [PubMed]
- NIH. Ritonavir-Boosted Nirmatrelvir (Paxlovid). Available online: https://www.covid19treatmentguidelines.nih.gov/therapies/antiviral-therapy/ritonavir-boosted-nirmatrelvir--paxlovid-/ (accessed on 18 November 2022).
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.-H. An overview of severe acute respiratory syndrome–coronavirus (SARS-CoV) 3CL protease inhibitors: Peptidomimetics and small molecule chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef]
- GILEAD. Veklury® (Remdesivir) Retains Antiviral Activity Against Omicron, Delta and Other Emergent SARS-CoV-2 Variants in Multiple In Vitro Studies. Available online: https://www.gilead.com/news-and-press/press-room/press-releases/2022/2/veklury-remdesivir-retains-antiviral-activity-against-omicron-delta-and-other-emergent-sarscov2-variants-in-multiple-in-vitro-studies (accessed on 18 November 2022).
- Eastman, R.T.; Roth, J.S.; Brimacombe, K.R.; Simeonov, A.; Shen, M.; Patnaik, S.; Hall, M.D. Remdesivir: A review of its discovery and development leading to emergency use authorization for treatment of COVID-19. ACS Cent. Sci. 2020, 6, 672–683. [Google Scholar] [CrossRef]
- Westendorf, K.; Žentelis, S.; Wang, L.; Foster, D.; Vaillancourt, P.; Wiggin, M.; Lovett, E.; van der Lee, R.; Hendle, J.; Pustilnik, A. LY-CoV1404 (bebtelovimab) potently neutralizes SARS-CoV-2 variants. Cell Rep. 2022, 39, 110812. [Google Scholar] [CrossRef] [PubMed]
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes New Monoclonal Antibody for Treatment of COVID-19 that Retains Activity Against Omicron Variant. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-new-monoclonal-antibody-treatment-covid-19-retains (accessed on 23 November 2022).
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Dougan, M.; Nirula, A.; Azizad, M.; Mocherla, B.; Gottlieb, R.L.; Chen, P.; Hebert, C.; Perry, R.; Boscia, J.; Heller, B. Bamlanivimab plus etesevimab in mild or moderate Covid-19. N. Engl. J. Med. 2021, 385, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Monoclonal Antibodies for Treatment of COVID-19. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-monoclonal-antibodies-treatment-covid-19 (accessed on 23 November 2022).
- Phan, A.T.; Gukasyan, J.; Arabian, S.; Wang, S.; Neeki, M.M. Emergent inpatient administration of casirivimab and imdevimab antibody cocktail for the treatment of COVID-19 pneumonia. Cureus 2021, 13, e15280. [Google Scholar] [CrossRef]
- Gupta, A.; Gonzalez-Rojas, Y.; Juarez, E.; Casal, M.C.; Moya, J.; Falci, D.R.; Sarkis, E.; Solis, J.; Zheng, H.; Scott, N. Effect of sotrovimab on hospitalization or death among high-risk patients with mild to moderate COVID-19: A randomized clinical trial. JAMA 2022, 327, 1236–1246. [Google Scholar] [CrossRef]
- Martin-Blondel, G.; Marcelin, A.-G.; Soulié, C.; Kaisaridi, S.; Lusivika-Nzinga, C.; Dorival, C.; Nailler, L.; Boston, A.; Melenotte, C.; Cabié, A. Sotrovimab to prevent severe COVID-19 in high-risk patients infected with Omicron BA. 2. J. Infect. 2022, 85, e104–e108. [Google Scholar] [CrossRef]
- Abramowicz, M.; Zucotti, G.; Pflomm, M. Tixagevimab and Cilgavimab (Evusheld) for Pre-Exposure Prophylaxis of COVID-19. JAMA 2022, 327, 384–385. [Google Scholar]
- Focosi, D.; Casadevall, A. A Critical Analysis of the Use of Cilgavimab plus Tixagevimab Monoclonal Antibody Cocktail (Evusheld™) for COVID-19 Prophylaxis and Treatment. Viruses 2022, 14, 1999. [Google Scholar] [CrossRef] [PubMed]
- CDC. SARS-CoV-2 Variant Classifications and Definitions. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/variant-classifications.html (accessed on 24 November 2022).
- Meredith, L.W.; Hamilton, W.L.; Warne, B.; Houldcroft, C.J.; Hosmillo, M.; Jahun, A.S.; Curran, M.D.; Parmar, S.; Caller, L.G.; Caddy, S.L. Rapid implementation of SARS-CoV-2 sequencing to investigate cases of health-care associated COVID-19: A prospective genomic surveillance study. Lancet Infect. Dis. 2020, 20, 1263–1271. [Google Scholar] [CrossRef]
- GISAID. Tracking of hCoV-19 Variants. Available online: https://gisaid.org/hcov19-variants/ (accessed on 25 November 2022).
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.-W.; Sahi, V.; Figueroa, A. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Cosar, B.; Karagulleoglu, Z.Y.; Unal, S.; Ince, A.T.; Uncuoglu, D.B.; Tuncer, G.; Kilinc, B.R.; Ozkan, Y.E.; Ozkoc, H.C.; Demir, I.N. SARS-CoV-2 mutations and their viral variants. Cytokine Growth Factor Rev. 2021, 63, 10–22. [Google Scholar] [CrossRef]
- Tuekprakhon, A.; Nutalai, R.; Dijokaite-Guraliuc, A.; Zhou, D.; Ginn, H.M.; Selvaraj, M.; Liu, C.; Mentzer, A.J.; Supasa, P.; Duyvesteyn, H.M. Antibody escape of SARS-CoV-2 Omicron BA. 4 and BA. 5 from vaccine and BA. 1 serum. Cell 2022, 185, 2422–2433. [Google Scholar] [CrossRef]
- Syed, A.M.; Ciling, A.; Taha, T.Y.; Chen, I.P.; Khalid, M.M.; Sreekumar, B.; Chen, P.-Y.; Kumar, G.R.; Suryawanshi, R.; Silva, I. Omicron mutations enhance infectivity and reduce antibody neutralization of SARS-CoV-2 virus-like particles. Proc. Natl. Acad. Sci. USA 2022, 119, e2200592119. [Google Scholar] [CrossRef]
- Mileto, D.; Micheli, V.; Fenizia, C.; Cutrera, M.; Gagliardi, G.; Mancon, A.; Bracchitta, F.; De Silvestri, A.; Rizzardini, G.; Lombardi, A. Reduced neutralization of SARS-CoV-2 Omicron variant by BNT162b2 vaccinees’ sera: A preliminary evaluation. Emerg. Microbes Infect. 2022, 11, 790–792. [Google Scholar] [CrossRef]
- Wang, W.B.; Liang, Y.; Jin, Y.Q.; Zhang, J.; Su, J.G.; Li, Q.M. E484K mutation in SARS-CoV-2 RBD enhances binding affinity with hACE2 but reduces interactions with neutralizing antibodies and nanobodies: Binding free energy calculation studies. J. Mol. Graph. Model. 2021, 109, 108035. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Rangarajan, E.S.; Izard, T.; Farzan, M.; Choe, H. The D614G mutation in the SARS-CoV-2 spike protein reduces S1 shedding and increases infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef] [PubMed]
- Volz, E.; Hill, V.; McCrone, J.T.; Price, A.; Jorgensen, D.; O’Toole, Á.; Southgate, J.; Johnson, R.; Jackson, B.; Nascimento, F.F. Evaluating the effects of SARS-CoV-2 spike mutation D614G on transmissibility and pathogenicity. Cell 2021, 184, 64–75.e11. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.C.; Rosen, L.E.; Shepherd, J.G.; Spreafico, R.; da Silva Filipe, A.; Wojcechowskyj, J.A.; Davis, C.; Piccoli, L.; Pascall, D.J.; Dillen, J. Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. Cell 2021, 184, 1171–1187.e20. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Lan, J.; Ju, X.; Gong, M.; Long, Q.; Zhu, Z.; Yu, Y.; Wu, J.; Zhong, J.; Zhang, R. Mutation Y453F in the spike protein of SARS-CoV-2 enhances interaction with the mink ACE2 receptor for host adaption. PLoS Pathog. 2021, 17, e1010053. [Google Scholar] [CrossRef]
- Bayarri-Olmos, R.; Rosbjerg, A.; Johnsen, L.B.; Helgstrand, C.; Bak-Thomsen, T.; Garred, P.; Skjoedt, M.-O. The SARS-CoV-2 Y453F mink variant displays a pronounced increase in ACE-2 affinity but does not challenge antibody neutralization. J. Biol. Chem. 2021, 296, 100536. [Google Scholar] [CrossRef]
- Klinakis, A.; Cournia, Z.; Rampias, T. N-terminal domain mutations of the spike protein are structurally implicated in epitope recognition in emerging SARS-CoV-2 strains. Comput. Struct. Biotechnol. J. 2021, 19, 5556–5567. [Google Scholar] [CrossRef]
- Kemp, S.A.; Meng, B.; Ferriera, I.A.; Datir, R.; Harvey, W.T.; Papa, G.; Lytras, S.; Collier, D.A.; Mohamed, A.; Gallo, G. Recurrent emergence and transmission of a SARS-CoV-2 spike deletion H69/V70. BioRxiv 2021. [Google Scholar] [CrossRef]
- Tian, F.; Tong, B.; Sun, L.; Shi, S.; Zheng, B.; Wang, Z.; Dong, X.; Zheng, P. N501Y mutation of spike protein in SARS-CoV-2 strengthens its binding to receptor ACE2. eLife 2021, 10, e69091. [Google Scholar] [CrossRef]
- Jangra, S.; Ye, C.; Rathnasinghe, R.; Stadlbauer, D.; Krammer, F.; Simon, V.; Martinez-Sobrido, L.; Garcia-Sastre, A.; Schotsaert, M.; Group, P.S. The E484K mutation in the SARS-CoV-2 spike protein reduces but does not abolish neutralizing activity of human convalescent and post-vaccination sera. MedRxiv 2021. [Google Scholar] [CrossRef]
- Yang, W.-T.; Huang, W.-H.; Liao, T.-L.; Hsiao, T.-H.; Chuang, H.-N.; Liu, P.-Y. SARS-CoV-2 E484K Mutation Narrative Review: Epidemiology, Immune Escape, Clinical Implications, and Future Considerations. Infect. Drug Resist. 2022, 15, 373. [Google Scholar] [CrossRef] [PubMed]
- Fratev, F. N501Y and K417N mutations in the spike protein of SARS-CoV-2 alter the interactions with Both hACE2 and human-derived antibody: A free energy of perturbation retrospective study. J. Chem. Inf. Model. 2021, 61, 6079–6084. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, Y.; Schmidt, F.; Zhang, F.; DaSilva, J.; Poston, D.; Lorenzi, J.C.; Muecksch, F.; Rutkowska, M.; Hoffmann, H.-H.; Michailidis, E. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. eLife 2020, 9, e61312. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Sharma, A.R.; Dhama, K.; Agoramoorthy, G.; Chakraborty, C. Omicron variant (B. 1.1. 529) of SARS-CoV-2: Understanding mutations in the genome, S-glycoprotein, and antibody-binding regions. GeroScience 2022, 44, 619–637. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, T.; Fang, Y.; Liu, J.; Ye, Q.; Ding, L. SARS-CoV-2 spike L452R mutation increases Omicron variant fusogenicity and infectivity as well as host glycolysis. Signal Transduct. Target. Ther. 2022, 7, 1–3. [Google Scholar] [CrossRef]
- Saito, A.; Irie, T.; Suzuki, R.; Maemura, T.; Nasser, H.; Uriu, K.; Kosugi, Y.; Shirakawa, K.; Sadamasu, K.; Kimura, I. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature 2022, 602, 300–306. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, Y.; Iketani, S.; Nair, M.S.; Li, Z.; Mohri, H.; Wang, M.; Yu, J.; Bowen, A.D.; Chang, J.Y. Antibody evasion by SARS-CoV-2 Omicron subvariants BA. 2.12. 1, BA. 4 and BA. 5. Nature 2022, 608, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Evans, J.P.; Zheng, Y.-M.; Carlin, C.; Saif, L.J.; Oltz, E.M.; Xu, K.; Gumina, R.J.; Liu, S.-L. Evasion of neutralizing antibody responses by the SARS-CoV-2 BA. 2.75 variant. Cell Host Microbe 2022, 30, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Hachmann, N.P.; Miller, J.; Collier, A.-r.Y.; Barouch, D.H. Neutralization escape by SARS-CoV-2 Omicron subvariant BA. 4.6. N. Engl. J. Med. 2022, 387, 1904–1906. [Google Scholar] [CrossRef]
- Jian, F.; Yu, Y.; Song, W.; Yisimayi, A.; Yu, L.; Gao, Y.; Zhang, N.; Wang, Y.; Shao, F.; Hao, X. Further humoral immunity evasion of emerging SARS-CoV-2 BA. 4 and BA. 5 subvariants. Lancet Infect. Dis. 2022, 22, 1535–1537. [Google Scholar] [CrossRef]
- WHO. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 25 November 2022).
- Katella, K. Omicron, Delta, Alpha, and More: What To Know About the Coronavirus Variants. Available online: https://www.yalemedicine.org/news/covid-19-variants-of-concern-omicron (accessed on 25 November 2022).
- CDC. Variant Proportions. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/index.html (accessed on 25 November 2022).
- WHO. TAG-VE statement on Omicron sublineages BQ.1 and XBB. Available online: https://www.who.int/news/item/27-10-2022-tag-ve-statement-on-omicron-sublineages-bq.1-and-xbb (accessed on 26 November 2022).
- Hwang, Y.-C.; Lu, R.-M.; Su, S.-C.; Chiang, P.-Y.; Ko, S.-H.; Ke, F.-Y.; Liang, K.-H.; Hsieh, T.-Y.; Wu, H.-C. Monoclonal antibodies for COVID-19 therapy and SARS-CoV-2 detection. J. Biomed. Sci. 2022, 29, 1–50. [Google Scholar] [CrossRef]
- Olivera-Ugarte, S.-M.; Bolduc, M.; Laliberté-Gagné, M.-È.; Blanchette, L.-J.; Garneau, C.; Fillion, M.; Savard, P.; Dubuc, I.; Flamand, L.; Farnòs, O. A nanoparticle-based COVID-19 vaccine candidate elicits broad neutralizing antibodies and protects against SARS-CoV-2 infection. Nanomed. Nanotechnol. Biol. Med. 2022, 44, 102584. [Google Scholar] [CrossRef] [PubMed]
- Nathan, R.; Shawa, I.; De La Torre, I.; Pustizzi, J.M.; Haustrup, N.; Patel, D.R.; Huhn, G. A narrative review of the clinical practicalities of bamlanivimab and etesevimab antibody therapies for SARS-CoV-2. Infect. Dis. Ther. 2021, 10, 1933–1947. [Google Scholar] [CrossRef]
- Lau, E.H.; Tsang, O.T.; Hui, D.S.; Kwan, M.Y.; Chan, W.-h.; Chiu, S.S.; Ko, R.L.; Chan, K.H.; Cheng, S.; Perera, R.A. Neutralizing antibody titres in SARS-CoV-2 infections. Nat. Commun. 2021, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 2020, 369, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.C.; Adams, A.C.; Hufford, M.M.; De La Torre, I.; Winthrop, K.; Gottlieb, R.L. Neutralizing monoclonal antibodies for treatment of COVID-19. Nat. Rev. Immunol. 2021, 21, 382–393. [Google Scholar] [CrossRef]
- Walls, A.C.; Xiong, X.; Park, Y.-J.; Tortorici, M.A.; Snijder, J.; Quispe, J.; Cameroni, E.; Gopal, R.; Dai, M.; Lanzavecchia, A. Unexpected receptor functional mimicry elucidates activation of coronavirus fusion. Cell 2019, 176, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, A.; Chen, X.; Zhang, Y.; Yu, F.; Yue, S.; Li, A.; Zhang, J.; Pan, Z.; Yang, Y. A potent human monoclonal antibody with pan-neutralizing activities directly dislocates S trimer of SARS-CoV-2 through binding both up and down forms of RBD. Signal Transduct. Target. Ther. 2022, 7, 1–13. [Google Scholar] [CrossRef]
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Monoclonal Antibodies for Treatment of COVID-19. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-monoclonal-antibodies-treatment-covid-19-0 (accessed on 26 November 2022).
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Monoclonal Antibody for Treatment of COVID-19. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-monoclonal-antibody-treatment-covid-19 (accessed on 26 November 2022).
- FDA. FDA Updates on Bebtelovimab. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-updates-bebtelovimab (accessed on 26 November 2022).
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes New Long-Acting Monoclonal Antibodies for Pre-exposure Prevention of COVID-19 in Certain Individuals. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-new-long-acting-monoclonal-antibodies-pre-exposure (accessed on 25 November 2022).
- U.S. Department of Health and Human Service. FDA Authorizes Revisions to Evusheld Dosing. Available online: https://aspr.hhs.gov/COVID-19/Therapeutics/updates/Pages/important-update-29June2022.aspx (accessed on 27 November 2022).
- FDA. FDA updates Sotrovimab Emergency Use Authorization. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-updates-sotrovimab-emergency-use-authorization (accessed on 25 November 2022).
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Additional Monoclonal Antibody for Treatment of COVID-19. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-additional-monoclonal-antibody-treatment-covid-19 (accessed on 27 November 2022).
- Antibody society. Anti-SARS-CoV-2 mAbs in Clinical Studies or Approved for Use. Available online: https://www.antibodysociety.org/covid-19-biologics-tracker/ (accessed on 12 December 2022).
- U.S. Department of Health and Human Service. Pause in the Distribution of Bamlanivimab/etesevimab. Available online: https://aspr.hhs.gov/COVID-19/Therapeutics/updates/Pages/important-update-25June2021.aspx (accessed on 26 November 2022).
- U.S. Department of Health and Human Service. FDA Authorizes Shelf-Life Extension for REGEN-COV from 24 months to 30 Months. Available online: https://aspr.hhs.gov/COVID-19/Therapeutics/updates/Pages/important-update-27June2022.aspx (accessed on 27 November 2022).
- Global times. Sinopharm’s F61 Nasal Spray Receives Clinical Trial Approval for People at High Risk of Exposure to COVID-19. Available online: https://www.globaltimes.cn/page/202211/1280675 (accessed on 12 December 2022).
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Drug for Treatment of COVID-19. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-drug-treatment-covid-19 (accessed on 12 December 2022).
- WHO. WHO Prequalifies First Monoclonal Antibody-tocilizumab-to Treat COVID-19. Available online: https://www.who.int/news/item/11-02-2022-who-prequalifies-first-monoclonal-antibody---tocilizumab-to-treat-covid-19 (accessed on 12 December 2022).
- U.S. National Library of Medicine. Sarilumab COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04327388 (accessed on 13 December 2022).
- U.S. National Library of Medicine. Study of Adalimumab or Placebo in Patients with Mild to Moderate COVID-19 (COMBAAT) (COMBAAT). Available online: https://clinicaltrials.gov/ct2/show/NCT04705844 (accessed on 13 December 2022).
- U.S. National Library of Medicine. Study of Efficacy and Safety of Canakinumab Treatment for CRS in Participants with COVID-19-induced Pneumonia (CAN-COVID). Available online: https://clinicaltrials.gov/ct2/show/NCT04362813 (accessed on 13 December 2022).
- U.S. National Library of Medicine. Efficacy and Safety Study of IV Ravulizumab in Patients with COVID-19 Severe Pneumonia. Available online: https://clinicaltrials.gov/ct2/show/NCT04369469 (accessed on 13 December 2022).
- Drugbank. Ravulizumab Recruiting Phase 4 Trials for Coronavirus Disease 2019 (COVID-19) Treatment. Available online: https://go.drugbank.com/drugs/DB11580/clinical_trials?conditions=DBCOND0129755&phase=4&purpose=treatment&status=recruiting (accessed on 13 December 2022).
- U.S. National Library of Medicine. Phase 3 Study to Evaluate Efficacy and Safety of Lenzilumab in Patients with COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04351152 (accessed on 13 December 2022).
- Gottlieb, R.L.; Nirula, A.; Chen, P.; Boscia, J.; Heller, B.; Morris, J.; Huhn, G.; Cardona, J.; Mocherla, B.; Stosor, V. Effect of bamlanivimab as monotherapy or in combination with etesevimab on viral load in patients with mild to moderate COVID-19: A randomized clinical trial. JAMA 2021, 325, 632–644. [Google Scholar] [CrossRef]
- Xiang, H.R.; He, B.; Li, Y.; Cheng, X.; Zhang, Q.Z.; Peng, W.X. Bamlanivimab plus etesevimab treatment have a better outcome against COVID-19: A meta-analysis. J. Med. Virol. 2022, 94, 1893–1905. [Google Scholar] [CrossRef]
- Lilly, E. Lilly’s Bamlanivimab and Etesevimab Together Reduced Hospitalizations and Death in Phase 3 Trial for Early COVID-19. Available online: https://investor.lilly.com/news-releases/news-release-details/lillys-bamlanivimab-and-etesevimab-together-reduced (accessed on 27 November 2022).
- Doggrell, S.A. Do we need bamlanivimab? Is etesevimab a key to treating COVID-19? Expert Opin. Biol. Ther. 2021, 21, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Chiba, S.; Halfmann, P.; Nagai, H. Efficacy of antibodies and antiviral drugs against Covid-19 omicron variant. N. Engl. J. Med. 2022, 386, 995–998. [Google Scholar] [CrossRef]
- Chen, B.; Haste, N.; Binkin, N.; Law, N.; Horton, L.; Yam, N.; Chen, V.; Abeles, S.R. Real World Effectiveness of Tixagevimab/cilgavimab (Evusheld) in the Omicron Era. medRxiv 2022. [Google Scholar] [CrossRef]
- Young-Xu, Y.; Epstein, L.; Marconi, V.C.; Davey, V.; Zwain, G.; Smith, J.; Korves, C.; Cunningham, F.; Bonomo, R.; Ginde, A.A. Tixagevimab/Cilgavimab for Prevention of COVID-19 during the Omicron Surge: Retrospective Analysis of National VA Electronic Data. medRxiv 2022. [Google Scholar] [CrossRef]
- Astrazeneca. Evusheld Long-Acting Antibody Combination Approved for Prevention and Treatment of COVID-19 in Japan. Available online: https://www.astrazeneca.com/media-centre/press-releases/2022/evusheld-approved-for-covid-19-in-japan.html (accessed on 27 November 2022).
- Loo, Y.-M.; McTamney, P.M.; Arends, R.H.; Gasser, R.A.; Abram, M.E.; Aksyuk, A.; Diallo, S.; Flores, D.J.; Kelly, E.J.; Ren, K. The SARS-CoV-2 monoclonal antibody combiantion, AZD7442, is protective in nonhuman primates and has an extended half-life in humans. Sci. Transl. Med. 2021, 14, eabl8124. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Ustianowski, A.; De Wit, S.; Launay, O.; Avila, M.; Templeton, A.; Yuan, Y.; Seegobin, S.; Ellery, A.; Levinson, D.J. Intramuscular AZD7442 (tixagevimab–cilgavimab) for prevention of COVID-19. N. Engl. J. Med. 2022, 386, 2188–2200. [Google Scholar] [CrossRef]
- FDA. FDA Releases Important Information about Risk of COVID-19 due to Certain Variants Not Neutralized by Evusheld. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-releases-important-information-about-risk-covid-19-due-certain-variants-not-neutralized-evusheld (accessed on 27 November 2022).
- Benotmane, I.; Velay, A.; Gautier-Vargas, G.; Olagne, J.; Thaunat, O.; Fafi-Kremer, S.; Caillard, S. Pre-exposure prophylaxis with 300 mg Evusheld elicits limited neutralizing activity against the Omicron variant. Kidney Int. 2022, 102, 442–444. [Google Scholar] [CrossRef]
- Hentzien, M.; Autran, B.; Piroth, L.; Yazdanpanah, Y.; Calmy, A. A monoclonal antibody stands out against omicron subvariants: A call to action for a wider access to bebtelovimab. Lancet Infect. Dis. 2022, 22, 1278. [Google Scholar] [CrossRef]
- Yamasoba, D.; Kosugi, Y.; Kimura, I.; Fujita, S.; Uriu, K.; Ito, J.; Sato, K. Neutralisation sensitivity of SARS-CoV-2 omicron subvariants to therapeutic monoclonal antibodies. Lancet Infect. Dis. 2022, 22, 942–943. [Google Scholar] [CrossRef]
- Orders, M. An EUA for Bebtelovimab for Treatment of COVID-19. Med. Lett. Drugs Ther. 2022, 64, 41–42. [Google Scholar]
- FDA. Fact Sheet for Health Care Providers. Emergency Use Authorization for Bebtelovimab. Available online: Chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://www.fda.gov/media/156152/download (accessed on 25 November 2022).
- De Gasparo, R.; Pedotti, M.; Simonelli, L.; Nickl, P.; Muecksch, F.; Cassaniti, I.; Percivalle, E.; Lorenzi, J.C.; Mazzola, F.; Magrì, D. Bispecific IgG neutralizes SARS-CoV-2 variants and prevents escape in mice. Nature 2021, 593, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.R.; Tomasovic, L.; Kuzmina, N.A.; Ronk, A.J.; Byrne, P.O.; Johnson, R.; Storm, N.; Olmedillas, E.; Hou, Y.J.; Schäfer, A. IgG-like bispecific antibodies with potent and synergistic neutralization against circulating SARS-CoV-2 variants of concern. Nat. Commun. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Montazersaheb, S.; Hosseiniyan Khatibi, S.M.; Hejazi, M.S.; Tarhriz, V.; Farjami, A.; Ghasemian Sorbeni, F.; Farahzadi, R.; Ghasemnejad, T. COVID-19 infection: An overview on cytokine storm and related interventions. Virol. J. 2022, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vardhana, S.A.; Wolchok, J.D. The many faces of the anti-COVID immune response. J. Exp. Med. 2020, 217, e20200678. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Dhimolea, E. Canakinumab. mAbs 2010, 2, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Katia, F.; Myriam, D.P.; Ucciferri, C.; Auricchio, A.; Di Nicola, M.; Marchioni, M.; Eleonora, C.; Emanuela, S.; Cipollone, F.; Vecchiet, J. Efficacy of canakinumab in mild or severe COVID-19 pneumonia. Immun. Inflamm. Dis. 2021, 9, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Chanchlani, N.; Lin, S.; Chee, D.; Hamilton, B.; Nice, R.; Arkir, Z.; Bewshea, C.; Cipriano, B.; Derikx, L.A.; Dunlop, A. Adalimumab and infliximab impair SARS-CoV-2 antibody responses: Results from a therapeutic drug monitoring study in 11,422 Biologic-Treated patients. J. Crohns. Colitis. 2022, 16, 389–397. [Google Scholar] [CrossRef]
- Jang, D.-i.; Lee, A.-H.; Shin, H.-Y.; Song, H.-R.; Park, J.-H.; Kang, T.-B.; Lee, S.-R.; Yang, S.-H. The role of tumor necrosis factor alpha (TNF-α) in autoimmune disease and current TNF-α inhibitors in therapeutics. Internat. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: What treatments are being investigated? BMJ 2020, 368, m1252. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Bohn, M.-F.; Ljungars, A. The challenges with developing therapeutic monoclonal antibodies for pandemic application. Expert Opin. Drug Discov. 2022, 17, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, L.B.; Tedla, N.; Bull, R.A. Broadly-neutralizing antibodies against emerging SARS-CoV-2 variants. Front. Immunol. 2021, 12, 752003. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, X.; Zhou, H.; Zhu, H.; Jiang, S.; Wang, P. Broadly neutralizing antibodies to SARS-CoV-2 and other human coronaviruses. Nat. Rev. Immunol. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Liao, H.; Li, Z. Clinical Development and Regulatory Pathways of Monoclonal Antibodies in Japan. Ther. Innov. Regul. Sci. 2022, 56, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Larkin, H.D. Expiration Dates Extended on COVID-19 Monoclonal Antibody Therapies. JAMA 2022, 328, 11. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Service. Shelf-Life Extension of Evusheld under Emergency Use Authorization. Available online: https://aspr.hhs.gov/COVID-19/Therapeutics/updates/Pages/important-update-28June2022.aspx (accessed on 13 December 2022).



{kind=link}
{kind=link}
| Therapeutics | Type | Target Protein (SARS-CoV-2) | References |
|---|---|---|---|
| Nirmatrelvir with Ritonavir (Paxlovid) | Antiviral drug | Viral protease (Mpro) | [25,26] |
| Remdesivir (Veklury) | Antiviral drug | RdRp | [27,28] |
| Bebtelovimab | Monoclonal antibody | Spike protein (RBD) | [29,30] |
| Molnupiravir (Lagevrio) | Antiviral drug | RdRp | [31] |
| Bamlavinivmab with etesevimab | Monoclonal antibody | Surface spike glycoprotein | [32] |
| Casirivimab with imdevimab | Monoclonal antibody | Spike protein | [33,34] |
| Sotrovimab | Monoclonal antibody | Spike protein | [35,36] |
| Tixagevimab with cilgavimab | Monoclonal antibody | Spike protein | [37,38] |
| Mutations | Type of Mutation and Region | Impact | References |
|---|---|---|---|
| D614G | Amino acid substitution within receptor-binding motif (RBM) | - Increases infectivity. - Increases transmissibility. | [50,51] |
| N439K | Amino acid substitution within receptor-binding motif (RBM) | - Enhances the binding affinity for the ACE2 receptor. - Reduces the neutralizing activity of some monoclonal antibodies (mAbs) and polyclonal antibodies. | [52] |
| Y453F | Amino acid substitution within receptor-binding motif (RBM) | - Increases ACE2-binding affinity. - Enhances transmission capacity. | [53,54] |
| Δ69–70 | Amino acid deletion in the N-terminal domain (NTD) of the spike protein | - Affects the network of NTD loops. - Increases transmission capacity. | [55,56] |
| N501Y | Amino acid substitution within the RBD | - Strengthens S protein binding to receptor ACE2. - Facilitates immune escape (antibody). | [42,57] |
| E484K | Amino acid substitution within the RBD | Reduces the neutralizing activity of antibodies. | [58,59] |
| K417N | Amino acid substitution in the spike protein | - Increases the interaction with hACE2. - May abolished the antibody effect. | [60] |
| K444 Q/R/N | Amino acid substitution within the RBD | Reduces the neutralizing activity of antibodies. | [61] |
| V445E | Amino acid substitution within the RBD | Reduces the neutralizing activity of antibodies. | [42,61] |
| K150 T/Q/R/E | Amino acid substitution in NTD of the spike protein | Reduces the neutralizing activity of antibodies. | [61,62] |
| N148S | Amino acid substitution in NTD of the spike protein | Reduces the neutralizing activity of antibodies. | [61] |
| L452R | Amino acid substitution within the RBD | - Increases infectivity. - Increases viral fusogenicity. - Facilitates escape antibodies. | [63] |
| P681R | Amino acid substitution in the spike protein | - Enhances viral fusogenicity. - Increases viral pathogenicity. | [64] |
| F486V | Amino acid substitution within the RBD | - Facilitates escape from certain class 1 and 2 antibodies. | [65] |
| N460K | Amino acid substitution in the spike protein | - Enhances S processing. - Enhances the resistance to neutralizing antibodies. | [66] |
| R346T | Amino acid substitution in the spike protein | - Increases viral prevalence. - Increases the ability to evade neutralizing antibodies. | [67,68] |
| No | Antibodies Name | Current Status | References |
|---|---|---|---|
| 1 | Bebtelovimab | Remain authorized in the U.S. until further notice by the FDA. | [30,83] |
| 2 | Tixagevimab with cilgavimab | Remain authorized with the recommendation of repeat dosing every six months with a dose of 300 mg of tixagevimab and 300 mg of Cilgavimab. | [84,85] |
| 3 | Sotrovimab | Since 5 April 2022, no longer authorized in any U.S. region; approved in Australia, the UK, and the EU. | [86,87,88] |
| 4 | Bamlanivimab with etesevimab | Pausing all distribution. | [81,89] |
| 5 | Casirivimab with imdevimab | Currently not authorized in any U.S. region; however, it is recommended to be retained for future SARS-CoV-2 variants that may be susceptible. | [33,88,90] |
| 6 | Amubarvimab/romlusevimab | Approved in China. | [88] |
| 7 | Regdanvimab (CT-P59) | Approved in the Republic of Korea and the EU. | [88] |
| 8 | Ronapreve | Approved in Japan, the UK, the EU, and Australia. | [88] |
| 9 | F61 | Approved for clinical trials in China. | [91] |
| 10 | Tocilizumab | Authorized for emergency use in June 2021. | [92,93] |
| 11 | Sarilumab | Clinical trial phase 3. | [94] |
| 12 | Adalimumab | Clinical trial phase 3. | [95] |
| 13 | Canakinumab | Clinical trial phase 3. | [96] |
| 14 | Ravulizumab | Completed clinical phase 3; recruiting phase 4 trials. | [97,98] |
| 15 | Lenzilumab | Clinical trial phase 3. | [99] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widyasari, K.; Kim, J. A Review of the Currently Available Antibody Therapy for the Treatment of Coronavirus Disease 2019 (COVID-19). Antibodies 2023, 12, 5. https://doi.org/10.3390/antib12010005
Widyasari K, Kim J. A Review of the Currently Available Antibody Therapy for the Treatment of Coronavirus Disease 2019 (COVID-19). Antibodies. 2023; 12(1):5. https://doi.org/10.3390/antib12010005
Chicago/Turabian StyleWidyasari, Kristin, and Jinnam Kim. 2023. "A Review of the Currently Available Antibody Therapy for the Treatment of Coronavirus Disease 2019 (COVID-19)" Antibodies 12, no. 1: 5. https://doi.org/10.3390/antib12010005
APA StyleWidyasari, K., & Kim, J. (2023). A Review of the Currently Available Antibody Therapy for the Treatment of Coronavirus Disease 2019 (COVID-19). Antibodies, 12(1), 5. https://doi.org/10.3390/antib12010005