
Yeast Surface Display Platform for Rapid Selection of an Antibody Library via Sequential Counter Antigen Flow Cytometry


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Antibodies
2.2. Preparation of Hapten Protein Conjugates
Preparation of Biotin-Labeled Ovalbumin (OVA)
2.3. Construction of Plasmids for Yeast Surface Display (YSD) of Antibody Fragments
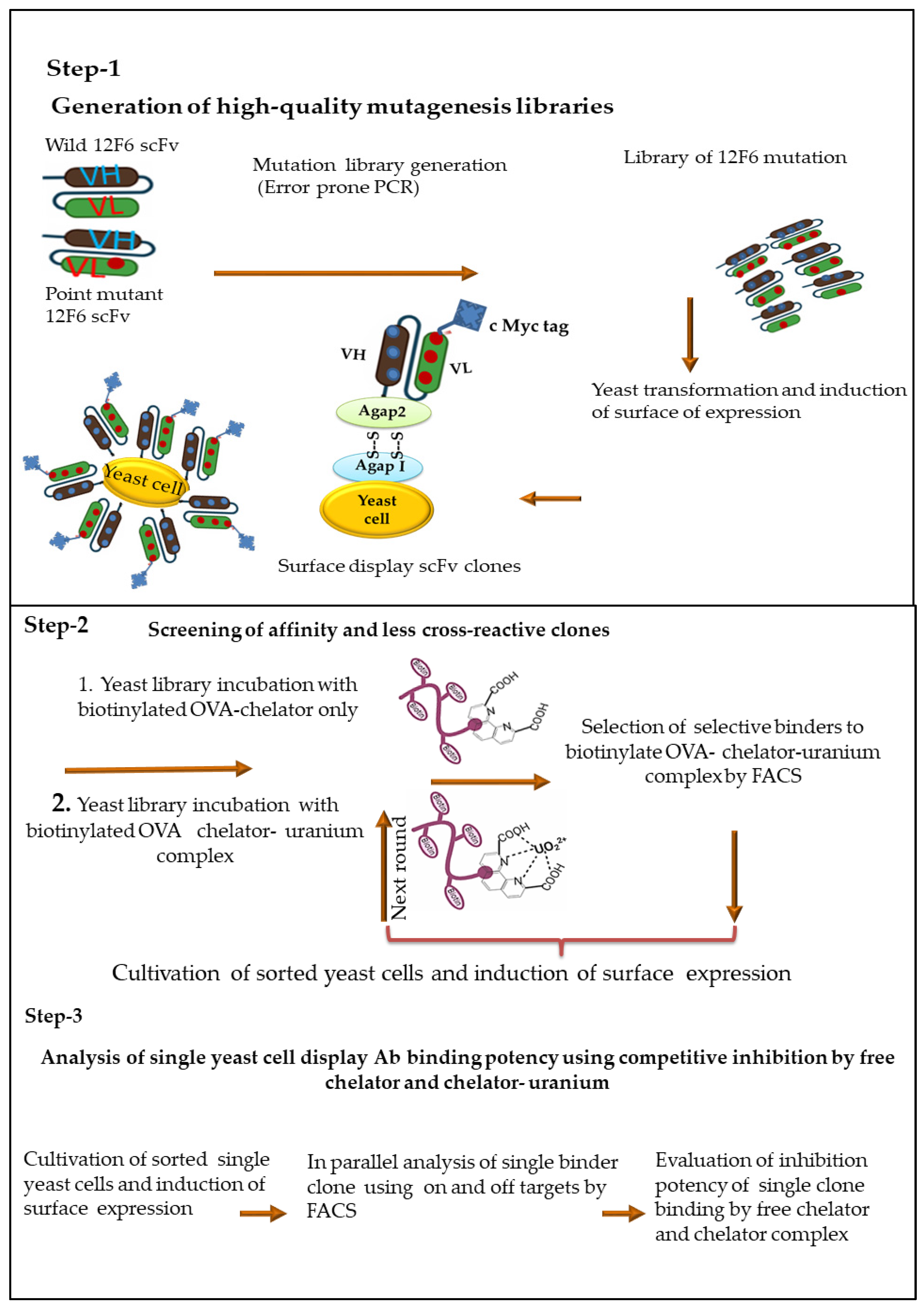
2.4. Generation Random Mutagenesis-Based Antibody Fragment Yeast Library
2.5. Flow Cytometry Binding Assay for Selection of Specific Yeast Clones against Metal Ion
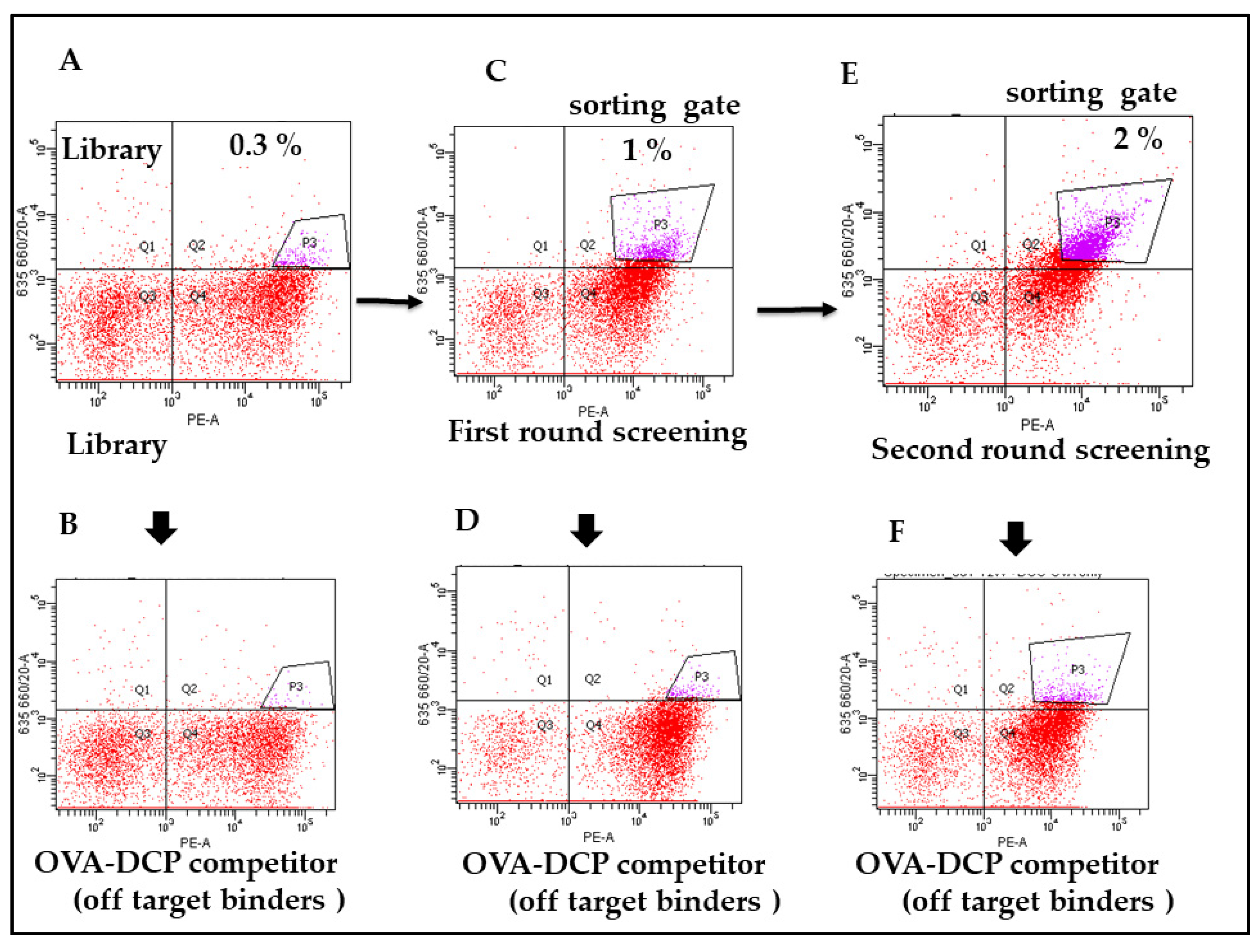
2.6. Metal Specific scFvs Yeast Library Screening by FACS Sorting
2.7. Evaluation of FACS Sorted Yeast Library Clones against Metal Ion
2.8. Soluble scFv Expression and Purification Binding Characterization of Soluble scFvs
2.9. Evaluation Selective Clones Binding Affinity and Specificity
3. Results
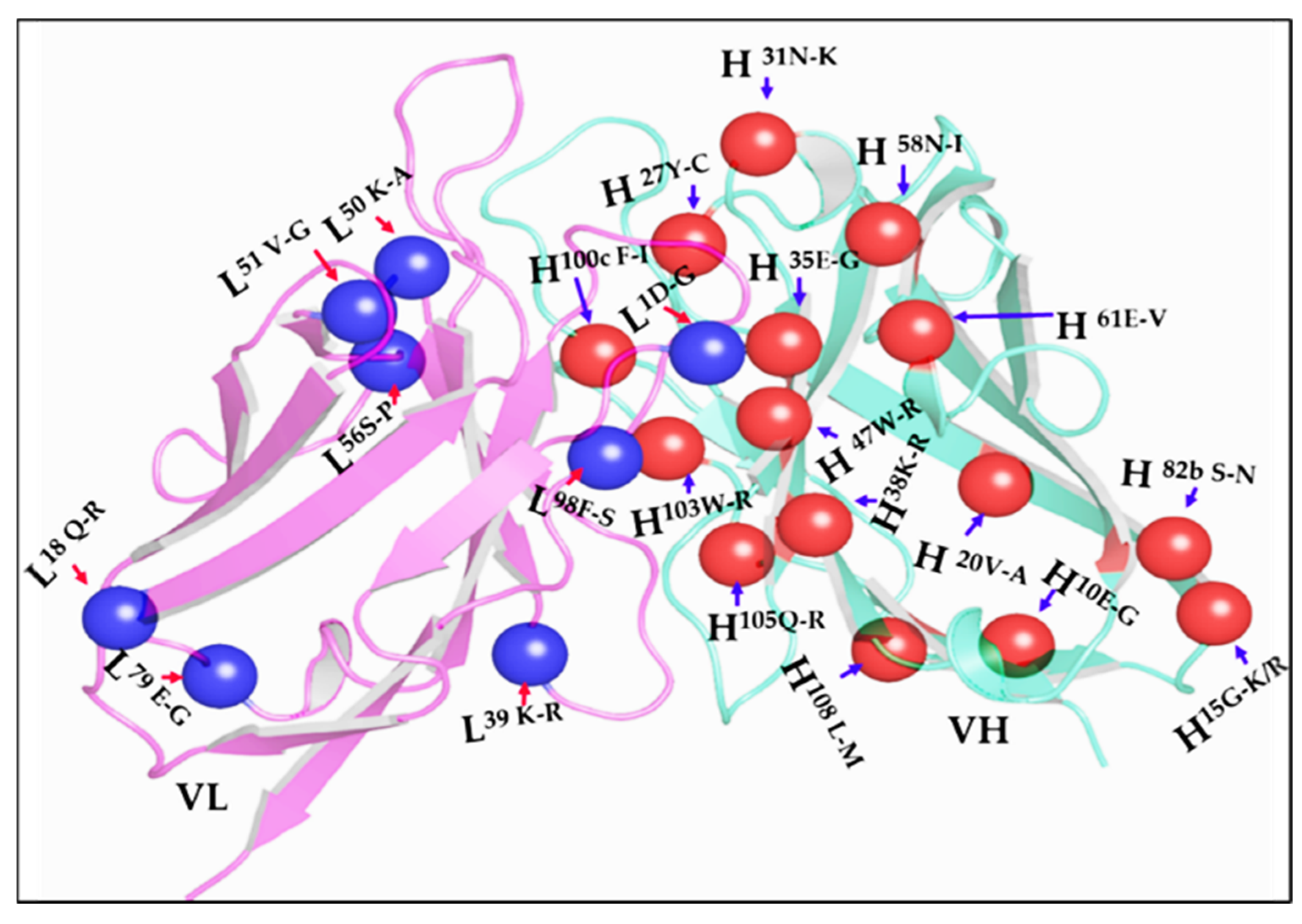
3.1. Multistep Random Mutagenesis Library of an Anti-Uranium 12F6 scFv
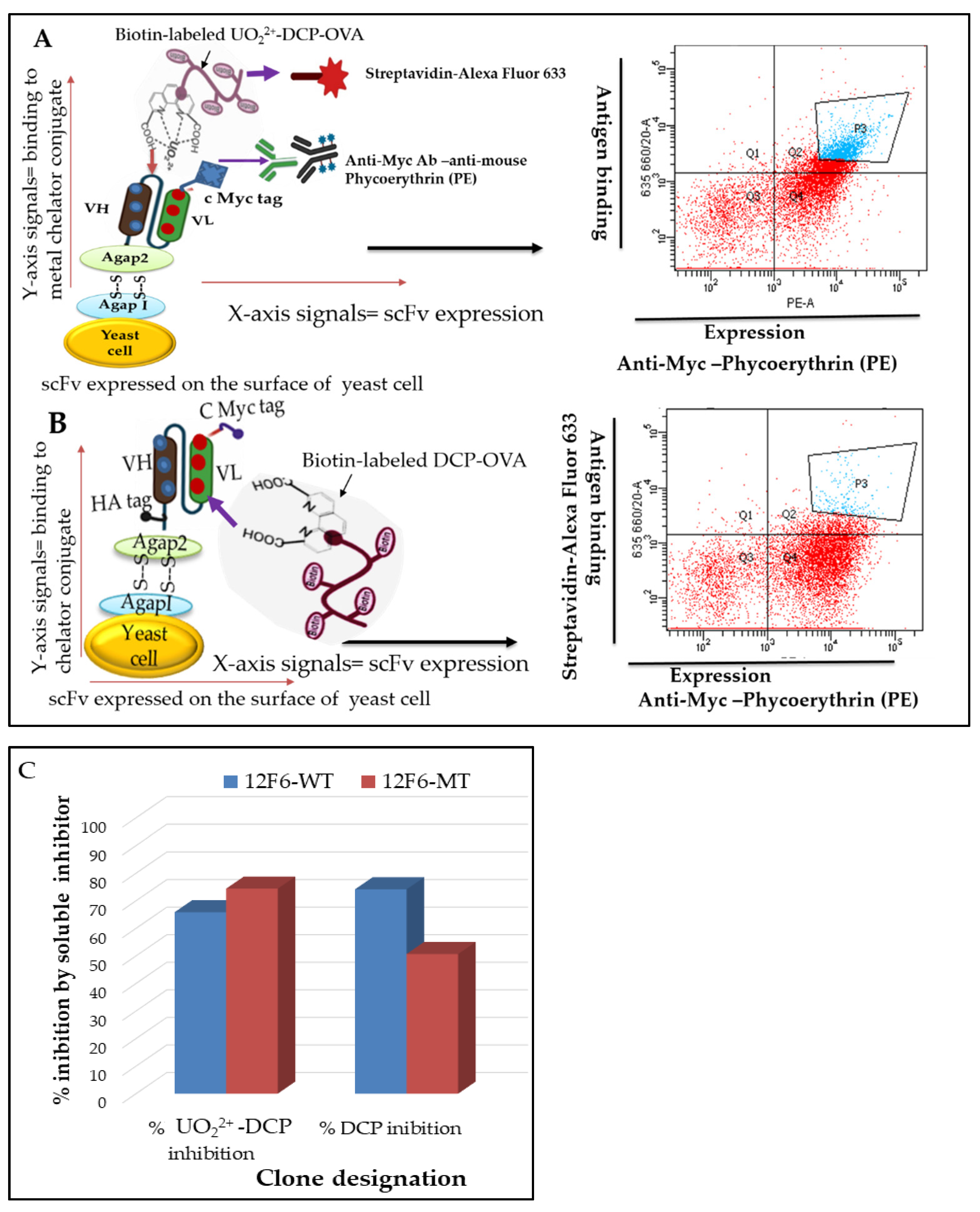
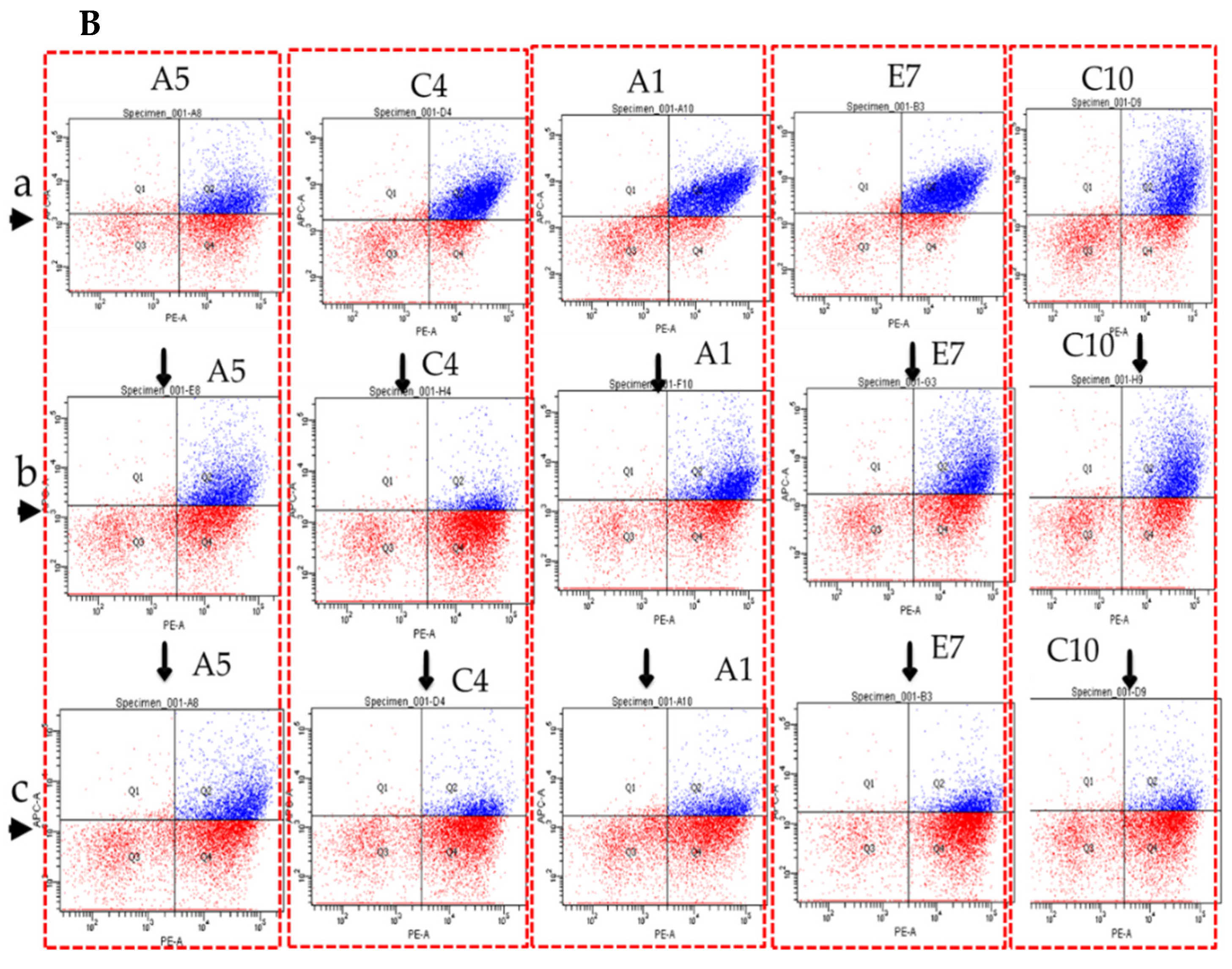
3.2. Generation and Validation of scFvs Display Yeast Cells by FACS Analysis
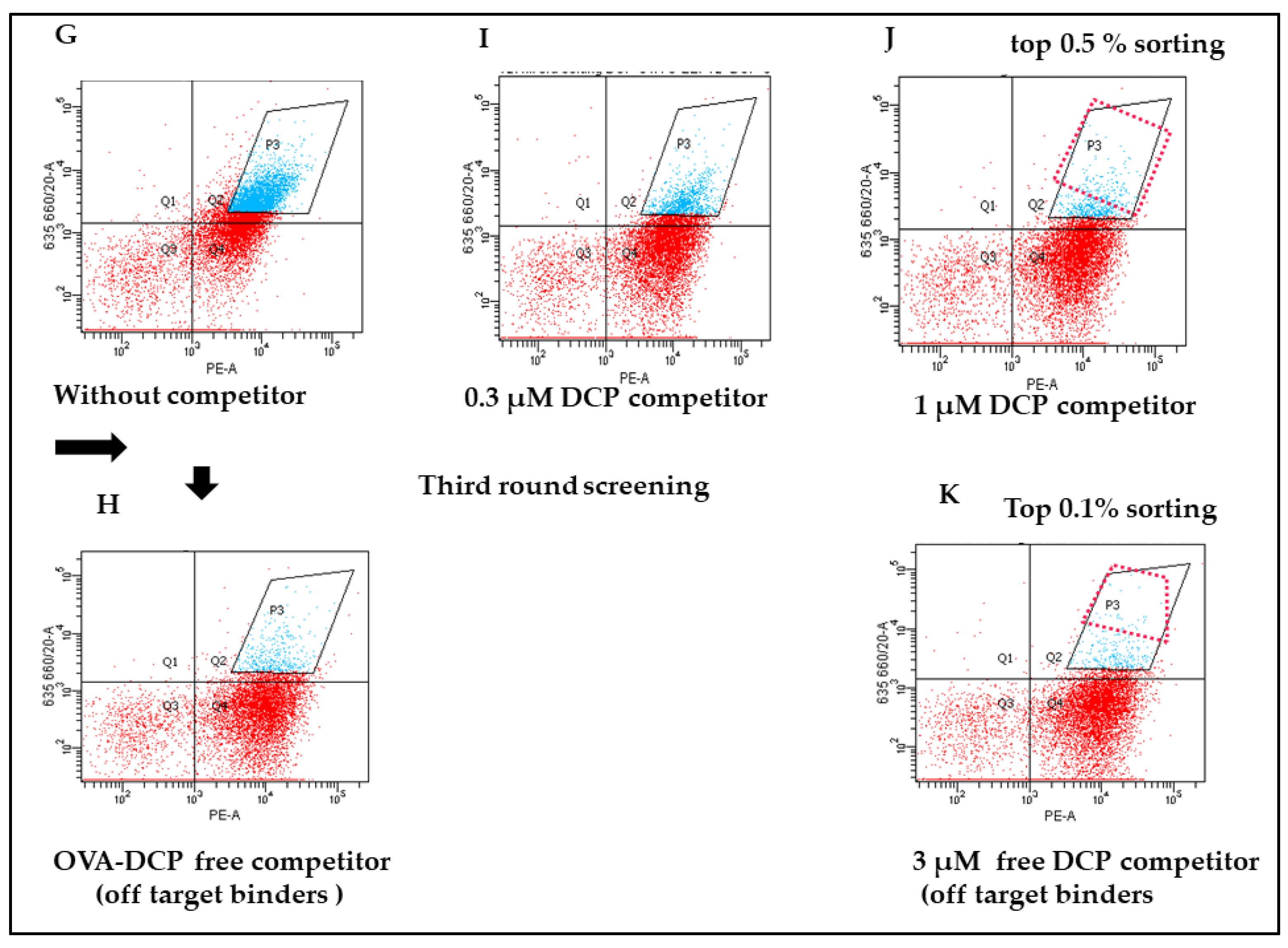
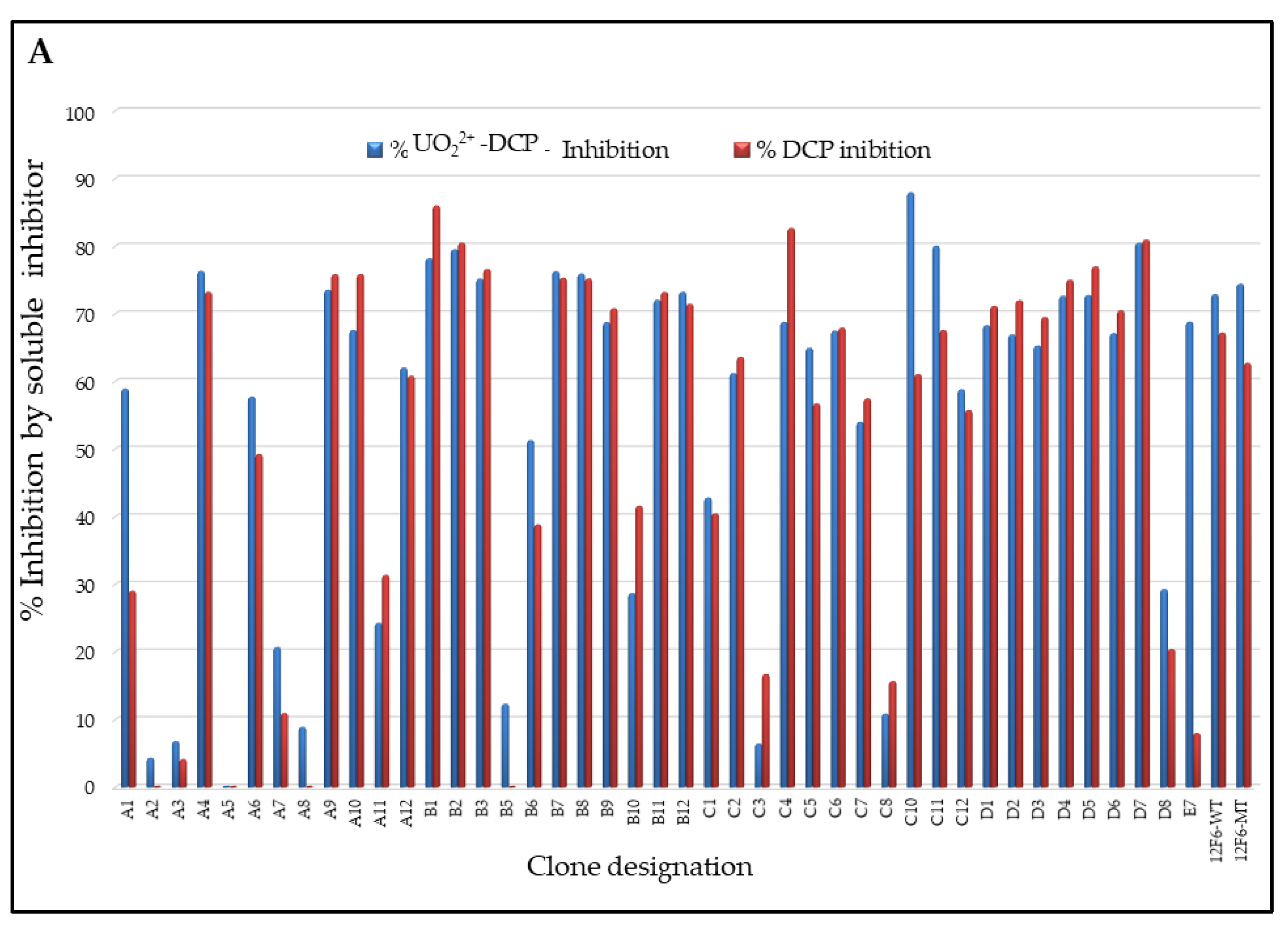
3.3. Specificity and Affinity Enrichment of Yeast-Displayed Mutated scFv Libraries
3.4. Competitive FACS of Monoclonal Yeast Cells, Sequence Analysis, and Competitive ELISA of Periplasmic Extracts
3.5. Studies of the Affinity of the 12F6 scFvs Mutants Using KinExA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bradbury, A.R.M.; Sidhu, S.; Dübel, S.; McCafferty, J. Beyond Natural Antibodies: The Power of in Vitro Display Technologies. Nat. Biotechnol. 2011, 29, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.A.; Arnold, F.H. Exploring Protein Fitness Landscapes by Directed Evolution. Nat. Rev. Mol. Cell Biol. 2009, 10, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Kearns-Jonker, M.; Barteneva, N.; Mencel, R.; Hussain, N.; Shulkin, I.; Xu, A.; Yew, M.; Cramer, D.V. Use of Molecular Modeling and Site-Directed Mutagenesis to Define the Structural Basis for the Immune Response to Carbohydrate Xenoantigens. BMC Immunol. 2007, 8, 3. [Google Scholar] [CrossRef]
- McCullum, E.O.; Williams, B.A.R.; Zhang, J.; Chaput, J.C. Random Mutagenesis by Error-Prone PCR. Methods Mol. Biol. Clifton NJ 2010, 634, 103–109. [Google Scholar]
- Forloni, M.; Liu, A.Y.; Wajapeyee, N. Random Mutagenesis Using Error-Prone DNA Polymerases. Cold Spring Harb. Protoc. 2018, 2018, 220–230. [Google Scholar] [CrossRef]
- Stemmer, W.P.C. Rapid Evolution of a Protein in Vitro by DNA Shuffling. Nature 1994, 370, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Neylon, C. Chemical and Biochemical Strategies for the Randomization of Protein Encoding DNA Sequences: Library Construction Methods for Directed Evolution. Nucleic Acids Res. 2004, 32, 1448–1459. [Google Scholar] [CrossRef]
- Cannon, D.A.; Shan, L.; Du, Q.; Shirinian, L.; Rickert, K.W.; Rosenthal, K.L.; Korade, M., III; van Vlerken-Ysla, L.E.; Buchanan, A.; Vaughan, T.J.; et al. Experimentally guided computational antibody affinity maturation with de novo docking, modelling and rational design. PLoS Comput. Biol. 2019, 15, e1006980. [Google Scholar] [CrossRef]
- Sheehan, J.; Marasco, W.A. Phage and Yeast Display. Microbiol. Spectr. 2015, 3, AID-0028. [Google Scholar] [CrossRef]
- Cherf, G.M.; Cochran, J.R. Applications of Yeast Surface Display for Protein Engineering. Methods Mol. Biol. Clifton NJ 2015, 1319, 155–175. [Google Scholar]
- Sun, Y.; Ban, B.; Bradbury, A.; Ansari, G.A.S.; Blake, D.A. Combining Yeast Display and Competitive FACS to Select Rare Hapten-Specific Clones from Recombinant Antibody Libraries. Anal. Chem. 2016, 88, 9181–9189. [Google Scholar] [CrossRef]
- Orcutt, K.D.; Slusarczyk, A.L.; Cieslewicz, M.; Ruiz-Yi, B.; Bhushan, K.R.; Frangioni, J.V.; Wittrup, K.D. Engineering an Antibody with Picomolar Affinity to DOTA Chelates of Multiple Radionuclides for Pretargeted Radioimmunotherapy and Imaging. Nucl. Med. Biol. 2011, 38, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, A.; Silva, D.-A.; Rocklin, G.J.; Hicks, D.R.; Vergara, R.; Murapa, P.; Bernard, S.M.; Zhang, L.; Lam, K.-H.; Yao, G.; et al. Massively Parallel de Novo Protein Design for Targeted Therapeutics. Nature 2017, 550, 74–79. [Google Scholar] [CrossRef] [PubMed]
- McMahon, C.; Baier, A.S.; Pascolutti, R.; Wegrecki, M.; Zheng, S.; Ong, J.X.; Erlandson, S.C.; Hilger, D.; Rasmussen, S.G.F.; Ring, A.M.; et al. Yeast Surface Display Platform for Rapid Discovery of Conformationally Selective Nanobodies. Nat. Struct. Mol. Biol. 2018, 25, 289–296. [Google Scholar] [CrossRef]
- Conroy, P.J.; Hearty, S.; Leonard, P.; O’Kennedy, R.J. Antibody Production, Design and Use for Biosensor-Based Applications. Semin. Cell Dev. Biol. 2009, 20, 10–26. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, T.; Wang, W.; Wen, Y.; Li, K.; Qian, L.; Zhang, X.; Liu, G. Lateral Flow Biosensors Based on the Use of Micro- and Nanomaterials: A Review on Recent Developments. Mikrochim. Acta 2019, 187, 70. [Google Scholar] [CrossRef]
- Ahn, K.C.; Kim, H.-J.; Mccoy, M.R.; Gee, S.J.; Hammock, B.D. Immunoassays and Biosensors for Monitoring Environmental and Human Exposure to Pyrethroid Insecticides. J. Agric. Food Chem. 2011, 59, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.A.; Ban, B.; Li, X.; Blake, R.C.; Jairo, G.A.; Sun, Y. Re-Engineering Antibodies for Optimum Performance in Uranium Sensors. In Uranium—Past and Future Challenges; Springer: Cham, Switzerland, 2015; pp. 529–536. [Google Scholar]
- Ahmed, S.; Ning, J.; Peng, D.; Chen, T.; Ahmad, I.; Ali, A.; Lei, Z.; Abu bakr Shabbir, M.; Cheng, G.; Yuan, Z. Current Advances in Immunoassays for the Detection of Antibiotics Residues: A Review. Food Agric. Immunol. 2020, 31, 268–290. [Google Scholar] [CrossRef]
- López Marzo, A.M.; Pons, J.; Blake, D.A.; Merkoçi, A. All-Integrated and Highly Sensitive Paper Based Device with Sample Treatment Platform for Cd2+ Immunodetection in Drinking/Tap Waters. Anal. Chem. 2013, 85, 3532–3538. [Google Scholar] [CrossRef]
- Quesada-González, D.; Jairo, G.A.; Blake, R.C.; Blake, D.A.; Merkoçi, A. Uranium (VI) Detection in Groundwater Using a Gold Nanoparticle/Paper-Based Lateral Flow Device. Sci. Rep. 2018, 8, 16157. [Google Scholar] [CrossRef]
- Blake, R.C.; Pavlov, A.R.; Khosraviani, M.; Ensley, H.E.; Kiefer, G.E.; Yu, H.; Li, X.; Blake, D.A. Novel Monoclonal Antibodies with Specificity for Chelated Uranium (VI): Isolation and Binding Properties. Bioconjug. Chem. 2004, 15, 1125–1136. [Google Scholar] [CrossRef] [PubMed]
- Melton, S.J.; Yu, H.; Williams, K.H.; Morris, S.A.; Long, P.E.; Blake, D.A. Field-Based Detection and Monitoring of Uranium in Contaminated Groundwater Using Two Immunosensors. Environ. Sci. Technol. 2009, 43, 6703–6709. [Google Scholar] [CrossRef]
- Feldhaus, M.; Siegel, R. Flow Cytometric Screening of Yeast Surface Display Libraries. Methods Mol. Biol. Clifton NJ 2004, 263, 311–332. [Google Scholar]
- Chao, G.; Lau, W.L.; Hackel, B.J.; Sazinsky, S.L.; Lippow, S.M.; Wittrup, K.D. Isolating and Engineering Human Antibodies Using Yeast Surface Display. Nat. Protoc. 2006, 1, 755–768. [Google Scholar] [CrossRef]
- Boder, E.T.; Wittrup, K.D. Yeast Surface Display for Screening Combinatorial Polypeptide Libraries. Nat. Biotechnol. 1997, 15, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Gai, S.A.; Wittrup, K.D. Yeast surface display for protein engineering and characterization. Curr. Opin. Struct. Biol. 2007, 17, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Grzeschik, J.; Yanakieva, D.; Roth, L.; Krah, S.; Hinz, S.C.; Elter, A.; Zollmann, T.; Schwall, G.; Zielonka, S.; Kolmar, H. Yeast Surface Display in Combination with Fluorescence-Activated Cell Sorting Enables the Rapid Isolation of Antibody Fragments Derived from Immunized Chickens. Biotechnol. J. 2019, 14, 1800466. [Google Scholar] [CrossRef]
- Zhu, X.; Kriegel, A.M.; Boustany, C.A.; Blake, D.A. Single-Chain Variable Fragment (scFv) Antibodies Optimized for Environmental Analysis of Uranium. Anal. Chem. 2011, 83, 3717–3724. [Google Scholar] [CrossRef]
- Khosraviani, M.; Blake, R.C.; Pavlov, A.R.; Lorbach, S.C.; Yu, H.; Delehanty, J.B.; Brechbiel, M.W.; Blake, D.A. Binding Properties of a Monoclonal Antibody Directed toward Lead−ChelateComplexes. Bioconjug. Chem. 2000, 11, 267–277. [Google Scholar] [CrossRef]
- Colby, D.W.; Kellogg, B.A.; Graff, C.P.; Yeung, Y.A.; Swers, J.S.; Wittrup, K.D. Engineering Antibody Affinity by Yeast Surface Display. Methods Enzymol. 2004, 388, 348–358. [Google Scholar]
- Kobayashi, N.; Oyama, H.; Kato, Y.; Goto, J.; Söderlind, E.; Borrebaeck, C.A.K. Two-Step in Vitro Antibody Affinity Maturation Enables Estradiol-17beta Assays with More than 10-Fold Higher Sensitivity. Anal. Chem. 2010, 82, 1027–1038. [Google Scholar] [CrossRef]
- Livesay, D.R.; Subramaniam, S. Conserved Sequence and Structure Association Motifs in Antibody-Protein and Antibody-Hapten Complexes. Protein Eng. Des. Sel. PEDS 2004, 17, 463–472. [Google Scholar] [CrossRef]
- Wang, N.; Smith, W.F.; Miller, B.R.; Aivazian, D.; Lugovskoy, A.A.; Reff, M.E.; Glaser, S.M.; Croner, L.J.; Demarest, S.J. Conserved Amino Acid Networks Involved in Antibody Variable Domain Interactions. Proteins 2009, 76, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Tsumoto, K.; Yokota, A.; Kondo, H.; Kumagai, I. Critical Contribution of VH-VL Interaction to Reshaping of an Antibody: The Case of Humanization of Anti-Lysozyme Antibody, HyHEL-10. Protein Sci. Publ. Protein Soc. 2008, 17, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Igawa, T.; Tsunoda, H.; Kikuchi, Y.; Yoshida, M.; Tanaka, M.; Koga, A.; Sekimori, Y.; Orita, T.; Aso, Y.; Hattori, K.; et al. VH/VL Interface Engineering to Promote Selective Expression and Inhibit Conformational Isomerization of Thrombopoietin Receptor Agonist Single-Chain Diabody. Protein Eng. Des. Sel. PEDS 2010, 23, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.W.; Baugher, W.; Rahn, T.; Drengler, S.; Tyner, J. Affinity Maturation of Tacrolimus Antibody for Improved Immunoassay Performance. Clin. Chem. 2008, 54, 1008–1017. [Google Scholar] [CrossRef][Green Version]
- Kabat, E.A.; Wu, T.T. Identical V Region Amino Acid Sequences and Segments of Sequences in Antibodies of Different Specificities. Relative Contributions of VH and VL Genes, Minigenes, and Complementarity-Determining Regions to Binding of Antibody-Combining Sites. J. Immunol. Baltim. Md 1950 1991, 147, 1709–1719. [Google Scholar]
- Chothia, C.; Novotný, J.; Bruccoleri, R.; Karplus, M. Domain Association in Immunoglobulin Molecules. The Packing of Variable Domains. J. Mol. Biol. 1985, 186, 651–663. [Google Scholar] [CrossRef]
- Al Qaraghuli, M.M.; Kubiak-Ossowska, K.; Ferro, V.A.; Mulheran, P.A. Structural Analysis of Anti-Hapten Antibodies to Identify Long-Range Structural Movements Induced by Hapten Binding. Front. Mol. Biosci. 2021, 8, 633526. [Google Scholar] [CrossRef]
- Midelfort, K.S.; Wittrup, K.D. Context-Dependent Mutations Predominate in an Engineered High-Affinity Single Chain Antibody Fragment. Protein Sci. Publ. Protein Soc. 2006, 15, 324–334. [Google Scholar] [CrossRef]
- Wu, D.; Sun, J.; Xu, T.; Wang, S.; Li, G.; Li, Y.; Cao, Z. Stacking and Energetic Contribution of Aromatic Islands at the Binding Interface of Antibody Proteins. Immunome Res. 2010, 6, S1. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Variants | Soluble UO22+-DCP Complex and DCP Fraction | Equilibrium Dissociation Constant (Kd, M) | Binding Fold Difference to DCP vs. UO22+-DCP Complex |
|---|---|---|---|
| 12F6-A1 | UO22+-DCP | 6.1 ± 0.075 × 10–8 M | 142 |
| 12F6-A1 | DCP | 8.7 ± 0.52 × 10–6 M | |
| 12F6-E7 | UO22+-DCP | 6.5 ± 0.08 × 10–8 M | 185 |
| 12F6-E7 | DCP | 1.2 ± 0.05 × 10–5 M | |
| 12F6-C5 | UO22+-DCP | 6.4 ± 0.08 × 10–8 M | 46 |
| 12F6-C5 | DCP | 3.0 ± 0.05 × 10–6 M | |
| 12F6-A6 | UO22+-DCP | 1.1 ± 0.7 × 10–7 M | 22 |
| 12F6-A6 | DCP | 2.5 ± 0.11 × 10–6 M | |
| 12F6-C10 | UO22+-DCP | 3.9 ± 0.08 × 10–8 M | 134 |
| 12F6-C10 | DCP | 5.2 ± 0.12 × 10–6 M | |
| 12F6 -WT | UO22+-DCP | 7.7 ± 0.9 × 10–9 M | 104 |
| 12F6-WT | DCP | 8.0 ± 0.6 × 10–7 M | |
| 12F6-MT | UO22+-DCP | 4.7 ± 0.5 × 10–9 M | 225 |
| 12F6-MT | DCP | 1.2 ± 0.09 ×10–6 M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ban, B.; Blake, R.C., II; Blake, D.A. Yeast Surface Display Platform for Rapid Selection of an Antibody Library via Sequential Counter Antigen Flow Cytometry. Antibodies 2022, 11, 61. https://doi.org/10.3390/antib11040061
Ban B, Blake RC II, Blake DA. Yeast Surface Display Platform for Rapid Selection of an Antibody Library via Sequential Counter Antigen Flow Cytometry. Antibodies. 2022; 11(4):61. https://doi.org/10.3390/antib11040061
Chicago/Turabian StyleBan, Bhupal, Robert C. Blake, II, and Diane A. Blake. 2022. "Yeast Surface Display Platform for Rapid Selection of an Antibody Library via Sequential Counter Antigen Flow Cytometry" Antibodies 11, no. 4: 61. https://doi.org/10.3390/antib11040061
APA StyleBan, B., Blake, R. C., II, & Blake, D. A. (2022). Yeast Surface Display Platform for Rapid Selection of an Antibody Library via Sequential Counter Antigen Flow Cytometry. Antibodies, 11(4), 61. https://doi.org/10.3390/antib11040061