
Long-Term Immunity and Antibody Response: Challenges for Developing Efficient COVID-19 Vaccines

 , , ,
, , ,
Abstract
:1. Introduction
2. COVID-19 Vaccine-Mediated Protection: Immunological Mechanisms
3. What Is the Duration of COVID-19 Immunity?
4. Persistence of SARS-CoV-2 Antibodies in the Body
5. Factors Involved in Inducing Long-Term Immunity for COVID-19 Vaccines
6. Concerns about Boosting COVID-19 Vaccines
7. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bok, K.; Sitar, S.; Graham, B.S.; Mascola, J.R. Accelerated COVID-19 vaccine development: Milestones, lessons, and prospects. Immunity 2021, 54, 1636–1651. [Google Scholar]
- Teijaro, J.R.; Farber, D.L. COVID-19 vaccines: Modes of immune activation and future challenges. Nat. Rev. Immunol. 2021, 21, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-D.; Chi, W.-Y.; Su, J.-H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. Coronavirus vaccine development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 1–23. [Google Scholar] [CrossRef]
- Han, X.; Xu, P.; Ye, Q. Analysis of COVID-19 vaccines: Types, thoughts, and application. J. Clin. Lab. Anal. 2021, 35, e23937. [Google Scholar] [CrossRef] [PubMed]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccines Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakney, A.K.; Bekker, L.-G. DNA vaccines join the fight against COVID-19. Lancet 2022, 399, 1281–1282. [Google Scholar] [CrossRef]
- Rahman, M.A.; Islam, M.S. Early approval of COVID-19 vaccines: Pros and cons. Hum. Vaccines Immunother. 2021, 17, 3288–3296. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: Principles, delivery and clinical translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef]
- Kremer, E.J. Pros and cons of adenovirus-based SARS-CoV-2 vaccines. Mol. Ther. 2020, 28, 2303–2304. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. Npj Vaccines 2021, 6, 1–17. [Google Scholar] [CrossRef]
- Kimman, T.G.; Cornelissen, L.A.; Moormann, R.J.; Rebel, J.M.; Stockhofe-Zurwieden, N. Challenges for porcine reproductive and respiratory syndrome virus (PRRSV) vaccinology. Vaccine 2009, 27, 3704–3718. [Google Scholar] [CrossRef]
- Altman, M.O.; Angeletti, D.; Yewdell, J.W. Antibody immunodominance: The key to understanding influenza virus antigenic drift. Viral Immunol. 2018, 31, 142–149. [Google Scholar] [CrossRef]
- Ning, T.; Nie, J.; Huang, W.; Li, C.; Li, X.; Liu, Q.; Zhao, H.; Wang, Y. Antigenic drift of influenza A (H7N9) virus hemagglutinin. J. Infect. Dis. 2019, 219, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Nadimuthu, L.P.R.; Victor, K. Environmental friendly micro cold storage for last-mile Covid-19 vaccine logistics. Environ. Sci. Pollut. Res. 2022, 29, 23767–23778. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.F.; Gaspar, P.D.; Souza, H.J.D. Refrigeration of COVID-19 vaccines: Ideal storage characteristics, energy efficiency and environmental impacts of various vaccine options. Energies 2021, 14, 1849. [Google Scholar] [CrossRef]
- Mercer, J.; Liang, A.; Yoon, J.; Nguyen, J.; Carroll, J.; Coley, K.C. COVID-19 pandemic vaccination preparedness strategies for independent community pharmacies. J. Am. Pharm. Assoc, 2022; in press. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Jaimes, J.A.; Millet, J.K.; Whittaker, G.R. Proteolytic cleavage of the SARS-CoV-2 spike protein and the role of the novel S1/S2 site. IScience 2020, 23, 101212. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.-J.; Jiang, S. The spike protein of SARS-CoV—a target for vaccine and therapeutic development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Moderbacher, C.R.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 2020, 183, 996–1012. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liang, B.; Chen, C.; Wang, H.; Fang, Y.; Shen, S.; Yang, X.; Wang, B.; Chen, L.; Chen, Q.; et al. SARS-CoV-2 infection induces sustained humoral immune responses in convalescent patients following symptomatic COVID-19. Nat. Commun. 2021, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Q.; Liu, Z.; Wang, Q.; Wu, J.; Hu, Y.; Bai, T.; Xie, T.; Huang, M.; Wu, T.; et al. Spike-specific circulating T follicular helper cell and cross-neutralizing antibody responses in COVID-19-convalescent individuals. Nat. Microbiol. 2021, 6, 51–58. [Google Scholar] [CrossRef]
- Vanshylla, K.; Di Cristanziano, V.; Kleipass, F.; Dewald, F.; Schommers, P.; Gieselmann, L.; Gruell, H.; Schlotz, M.; Ercanoglu, M.S.; Stumpf, R.; et al. Kinetics and correlates of the neutralizing antibody response to SARS-CoV-2 infection in humans. Cell Host Microbe 2021, 29, 917–929. [Google Scholar] [CrossRef]
- Bajaj, V.; Gadi, N.; Spihlman, A.P.; Wu, S.C.; Choi, C.H.; Moulton, V.R. Aging, immunity, and COVID-19: How age influences the host immune response to coronavirus infections? Front. Physiol. 2021, 11, 1793. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Sokal, A.; Chappert, P.; Barba-Spaeth, G.; Roeser, A.; Fourati, S.; Azzaoui, I.; Vandenberghe, A.; Fernandez, I.; Meola, A.; Bouvier-Alias, M.; et al. Maturation and persistence of the anti-SARS-CoV-2 memory B cell response. Cell 2021, 184, 1201–1213. [Google Scholar] [CrossRef]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.-W.; Sahi, V.; Figueroa, A.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef]
- Xu, C.; Wang, Y.; Liu, C.; Zhang, C.; Han, W.; Hong, X.; Wang, Y.; Hong, Q.; Wang, S.; Zhao, Q.; et al. Conformational dynamics of SARS-CoV-2 trimeric spike glycoprotein in complex with receptor ACE2 revealed by cryo-EM. Sci. Adv. 2021, 7, eabe5575. [Google Scholar] [CrossRef] [PubMed]
- McCallum, M.; De Marco, A.; Lempp, F.A.; Tortorici, M.A.; Pinto, D.; Walls, A.C.; Beltramello, M.; Chen, A.; Liu, Z.; Zatta, F.; et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. Cell 2021, 184, 2332–2347. [Google Scholar] [CrossRef] [PubMed]
- Tortorici, M.A.; Beltramello, M.; Lempp, F.A.; Pinto, D.; Dang, H.V.; Rosen, L.E.; McCallum, M.; Bowen, J.; Minola, A.; Jaconi, S.; et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science 2020, 370, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Park, Y.-J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A.; Bianchi, S.; Jaconi, S.; Culap, K.; Zatta, F.; De Marco, A.; et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef]
- Liu, L.; Wei, Q.; Lin, Q.; Fang, J.; Wang, H.; Kwok, H.; Tang, H.; Nishiura, K.; Peng, J.; Tan, Z.; et al. Anti–spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection. JCI Insight 2019, 4, e123158. [Google Scholar] [CrossRef]
- Li, D.; Edwards, R.J.; Manne, K.; Martinez, D.R.; Schäfer, A.; Alam, S.M.; Wiehe, K.; Lu, X.; Parks, R.; Sutherland, L.L.; et al. In vitro and in vivo functions of SARS-CoV-2 infection-enhancing and neutralizing antibodies. Cell 2021, 184, 4203–4219. [Google Scholar] [CrossRef]
- Escriou, N.; Callendret, B.; Lorin, V.; Combredet, C.; Marianneau, P.; Février, M.; Tangy, F. Protection from SARS coronavirus conferred by live measles vaccine expressing the spike glycoprotein. Virology 2014, 452, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Liniger, M.; Zuniga, A.; Tamin, A.; Azzouz-Morin, T.N.; Knuchel, M.; Marty, R.R.; Wiegand, M.; Weibel, S.; Kelvin, D.; Rota, P.A.; et al. Induction of neutralising antibodies and cellular immune responses against SARS coronavirus by recombinant measles viruses. Vaccine 2008, 26, 2164–2174. [Google Scholar] [CrossRef]
- Zhu, F.-C.; Li, Y.-H.; Guan, X.-H.; Hou, L.-H.; Wang, W.-J.; Li, J.-X.; Wu, S.-P.; Wang, B.-S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef]
- Bosch, B.J.; Martina, B.E.; Van Der Zee, R.; Lepault, J.; Haijema, B.J.; Versluis, C.; Heck, A.J.; De Groot, R.; Osterhaus, A.D.; Rottier, P.J. Severe acute respiratory syndrome coronavirus (SARS-CoV) infection inhibition using spike protein heptad repeat-derived peptides. Proc. Natl. Acad. Sci. USA 2004, 101, 8455–8460. [Google Scholar] [CrossRef] [Green Version]
- Keng, C.-T.; Zhang, A.; Shen, S.; Lip, K.-M.; Fielding, B.C.; Tan, T.H.; Chou, C.-F.; Loh, C.B.; Wang, S.; Fu, J.; et al. Amino acids 1055 to 1192 in the S2 region of severe acute respiratory syndrome coronavirus S protein induce neutralizing antibodies: Implications for the development of vaccines and antiviral agents. J. Virol. 2005, 79, 3289–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lip, K.-M.; Shen, S.; Yang, X.; Keng, C.-T.; Zhang, A.; Oh, H.-L.J.; Li, Z.-H.; Hwang, L.-A.; Chou, C.-F.; Fielding, B.C.; et al. Monoclonal antibodies targeting the HR2 domain and the region immediately upstream of the HR2 of the S protein neutralize in vitro infection of severe acute respiratory syndrome coronavirus. J. Virol. 2006, 80, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, G.; Li, J.; Nie, Y.; Shi, X.; Lian, G.; Wang, W.; Yin, X.; Zhao, Y.; Qu, X.; et al. Identification of an antigenic determinant on the S2 domain of the severe acute respiratory syndrome coronavirus spike glycoprotein capable of inducing neutralizing antibodies. J. Virol. 2004, 78, 6938–6945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yuan, Z.; Matsumoto, M.; Hengge, U.R.; Chang, Y.-F. Immune responses with DNA vaccines encoded different gene fragments of severe acute respiratory syndrome coronavirus in BALB/c mice. Biochem. Biophys. Res. Commun. 2005, 327, 130–135. [Google Scholar] [CrossRef]
- Yao, Y.; Jeyanathan, M.; Haddadi, S.; Barra, N.G.; Vaseghi-Shanjani, M.; Damjanovic, D.; Lai, R.; Afkhami, S.; Chen, Y.; Dvorkin-Gheva, A.; et al. Induction of autonomous memory alveolar macrophages requires T cell help and is critical to trained immunity. Cell 2018, 175, 1634–1650. [Google Scholar] [CrossRef] [Green Version]
- Widge, A.T.; Rouphael, N.G.; Jackson, L.A.; Anderson, E.J.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Durability of responses after SARS-CoV-2 mRNA-1273 vaccination. N. Engl. J. Med. 2021, 384, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH 1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Paik, D.H.; Farber, D.L. Anti-viral protective capacity of tissue resident memory T cells. Curr. Opin. Virol. 2021, 46, 20–26. [Google Scholar] [CrossRef]
- Mao, T.; Israelow, B.; Suberi, A.; Zhou, L.; Reschke, M.; Peña-Hernández, M.A.; Dong, H.; Homer, R.J.; Saltzman, W.M.; Iwasaki, A. Unadjuvanted intranasal spike vaccine booster elicits robust protective mucosal immunity against sarbecoviruses. bioRxiv 2022. [Google Scholar] [CrossRef]
- Knezevic, I.; Mattiuzzo, G.; Page, M.; Minor, P.; Griffiths, E.; Nuebling, M.; Moorthy, V. WHO International Standard for evaluation of the antibody response to COVID-19 vaccines: Call for urgent action by the scientific community. Lancet Microbe 2021, 3, e235–e240. [Google Scholar] [CrossRef]
- Omicron-Targeted Vaccines do no Better Than Original Jabs in Early Tests. Available online: https://www.nature.com/articles/d41586-022-00003-y (accessed on 5 May 2022).
- O’Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: Can it offer protection against COVID-19? Nat. Rev. Immunol. 2020, 20, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Ripperger, T.J.; Uhrlaub, J.L.; Watanabe, M.; Wong, R.; Castaneda, Y.; Pizzato, H.A.; Thompson, M.R.; Bradshaw, C.; Weinkauf, C.C.; Bime, C.; et al. Orthogonal SARS-CoV-2 serological assays enable surveillance of low-prevalence communities and reveal durable humoral immunity. Immunity 2020, 53, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Doria-Rose, N.; Suthar, M.S.; Makowski, M.; O’Connell, S.; McDermott, A.B.; Flach, B.; Ledgerwood, J.E.; Mascola, J.R.; Graham, B.S.; Lin, B.C.; et al. Antibody persistence through 6 months after the second dose of mRNA-1273 vaccine for Covid-19. N. Engl. J. Med. 2021, 384, 2259–2261. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.-M.; et al. Covid-19 vaccine effectiveness against the Omicron (B. 1.1. 529) variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: A pooled analysis of four randomised trials. Lancet 2021, 397, 881–891. [Google Scholar] [CrossRef]
- Dufloo, J.; Grzelak, L.; Staropoli, I.; Madec, Y.; Tondeur, L.; Anna, F.; Pelleau, S.; Wiedemann, A.; Planchais, C.; Buchrieser, J.; et al. Asymptomatic and symptomatic SARS-CoV-2 infections elicit polyfunctional antibodies. Cell Rep. Med. 2021, 2, 100275. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Mustapha, J.O.; Abdullahi, I.N.; Ajagbe, O.O.; Emeribe, A.U.; Fasogbon, S.A.; Onoja, S.O.; Ugwu, C.E.; Umeozuru, C.M.; Ajayi, F.O.; Tanko, W.N.; et al. Understanding the implications of SARS-CoV-2 re-infections on immune response milieu, laboratory tests and control measures against COVID-19. Heliyon 2021, 7, e05951. [Google Scholar] [CrossRef]
- Kaneko, N.; Kuo, H.-H.; Boucau, J.; Farmer, J.R.; Allard-Chamard, H.; Mahajan, V.S.; Piechocka-Trocha, A.; Lefteri, K.; Osborn, M.; Bals, J.; et al. Loss of Bcl-6-expressing T follicular helper cells and germinal centers in COVID-19. Cell 2020, 183, 143–157. [Google Scholar] [CrossRef]
- Hasan, A.; Al-Ozairi, E.; Al-Baqsumi, Z.; Ahmad, R.; Al-Mulla, F. Cellular and Humoral Immune Responses in Covid-19 and Immunotherapeutic Approaches. ImmunoTargets Ther. 2021, 10, 63. [Google Scholar] [CrossRef]
- Camou, F.; Tinevez, C.; Beguet-Yachine, M.; Bellecave, P.; Ratiarison, D.; Tumiotto, C.; Lafarge, X.; Guisset, O.; Mourissoux, G.; Lafon, M.E.; et al. Feasibility of convalescent plasma therapy in severe COVID-19 patients with persistent SARS-CoV-2 viremia. J. Med. Virol. 2021, 93, 5594–5598. [Google Scholar] [CrossRef] [PubMed]
- Moubarak, M.; Kasozi, K.I.; Hetta, H.F.; Shaheen, H.M.; Rauf, A.; Al-Kuraishy, H.M.; Qusti, S.; Alshammari, E.M.; Ayikobua, E.T.; Ssempijja, F. The rise of SARS-CoV-2 variants and the role of convalescent plasma therapy for management of infections. Life 2021, 11, 734. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.N.; Abdel-Moneim, A.S. Convalescent Plasma: A Potential Life-Saving Therapy for Coronavirus Disease 2019 (COVID-19). Front. Public Health 2020, 8, 437. [Google Scholar] [CrossRef] [PubMed]
- Janiaud, P.; Axfors, C.; Schmitt, A.M.; Gloy, V.; Ebrahimi, F.; Hepprich, M.; Smith, E.R.; Haber, N.A.; Khanna, N.; Moher, D.; et al. Association of Convalescent Plasma Treatment With Clinical Outcomes in Patients With COVID-19: A Systematic Review and Meta-analysis. JAMA 2021, 325, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Jedlicka, A.; Pekosz, A. The Xs and Y of immune responses to viral vaccines. Lancet Infect. Dis. 2010, 10, 338–349. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Conti, P.; Younes, A. Coronavirus COV-19/SARS-CoV-2 affects women less than men: Clinical response to viral infection. J. Biol. Regul. Homeost. Agents 2020, 34, 339–343. [Google Scholar]
- Schurz, H.; Salie, M.; Tromp, G.; Hoal, E.G.; Kinnear, C.J.; Möller, M. The X chromosome and sex-specific effects in infectious disease susceptibility. Hum. Genom. 2019, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, T.; Tober-Lau, P.; Hillus, D.; Helbig, E.T.; Lippert, L.J.; Thibeault, C.; Koch, W.; Landgraf, I.; Michel, J.; Bergfeld, L.; et al. Delayed antibody and T-cell response to BNT162b2 vaccination in the elderly, Germany. Emerg. Infect. Dis. 2021, 27, 2174. [Google Scholar] [CrossRef]
- Pannus, P.; Neven, K.Y.; De Craeye, S.; Heyndrickx, L.; Kerckhove, S.V.; Georges, D.; Michiels, J.; Francotte, A.; Van Den Bulcke, M.; Zrein, M.; et al. Poor antibody response to BioNTech/Pfizer COVID-19 vaccination in SARS-CoV-2 naïve residents of nursing homes. medRxiv 2021. [Google Scholar] [CrossRef]
- Castelo-Branco, C.; Soveral, I. The immune system and aging: A Review. Gynecol. Endocrinol. 2014, 30, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Fink, A.L.; Klein, S.L. Sex and gender impact immune responses to vaccines among the elderly. Physiology 2015, 30, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-S.; Wang, S.-T.; Chou, P. Efficacy and cost-effectiveness of influenza vaccination of the elderly in a densely populated and unvaccinated community. Vaccine 2002, 20, 2494–2499. [Google Scholar] [CrossRef]
- Callaway, E. COVID vaccine boosters: The most important questions. Nature 2021, 596, 178–180. [Google Scholar] [CrossRef]
- Kamar, N.; Abravanel, F.; Marion, O.; Couat, C.; Izopet, J.; Del Bello, A. Three doses of an mRNA Covid-19 vaccine in solid-organ transplant recipients. N. Engl. J. Med. 2021, 385, 661–662. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Kalkstein, N.; Mizrahi, B.; Alroy-Preis, S.; Ash, N.; Milo, R.; et al. Protection of BNT162b2 Vaccine Booster against Covid-19 in Israel. N. Engl. J. Med. 2021, 385, 1393–1400. [Google Scholar] [CrossRef]
- Bruxvoort, K.; Sy, L.S.; Qian, L.; Ackerson, B.K.; Luo, Y.; Lee, G.S.; Tian, Y.; Florea, A.; Takhar, H.S.; Tubert, J.E.; et al. Real-world effectiveness of the mRNA-1273 vaccine against COVID-19: Interim results from a prospective observational cohort study. Lancet Reg. Health Am. 2022, 6, 100134. [Google Scholar] [CrossRef]
- Thompson, M.G.; Stenehjem, E.; Grannis, S.; Ball, S.W.; Naleway, A.L.; Ong, T.C.; DeSilva, M.B.; Natarajan, K.; Bozio, C.H.; Lewis, N. Effectiveness of COVID-19 vaccines in ambulatory and inpatient care settings. New Engl. J. Med. 2021, 385, 1355–1371. [Google Scholar] [CrossRef]
- Chung, J.Y.; Thone, M.N.; Kwon, Y.J. COVID-19 vaccines: The status and perspectives in delivery points of view. Adv. Drug Deliv. Rev. 2020, 170, 1–25. [Google Scholar] [CrossRef]
- Pati, R.; Shevtsov, M.; Sonawane, A. Nanoparticle vaccines against infectious diseases. Front. Immunol. 2018, 9, 2224. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, G.; Madou, M.J.; Kalra, S.; Chopra, V.; Ghosh, D.; Martinez-Chapa, S.O. Nanotechnology for COVID-19: Therapeutics and vaccine research. ACS Nano 2020, 14, 7760–7782. [Google Scholar] [CrossRef] [PubMed]
- Sepand, M.R.; Aghsami, M.; Keshvadi, M.H.; Bigdelou, B.; Behzad, R.; Zanganeh, S.; Shadboorestan, A. The role of macrophage polarization and function in environmental toxicant-induced cancers. Environ. Res. 2021, 196, 110933. [Google Scholar] [CrossRef] [PubMed]
- Sepand, M.R.; Ghavami, M.; Zanganeh, S.; Stacks, S.; Ghasemi, F.; Montazeri, H.; Corbo, C.; Derakhshankhah, H.; Ostad, S.N.; Ghahremani, M.H.; et al. Impact of plasma concentration of transferrin on targeting capacity of nanoparticles. Nanoscale 2020, 12, 4935–4944. [Google Scholar] [CrossRef] [PubMed]


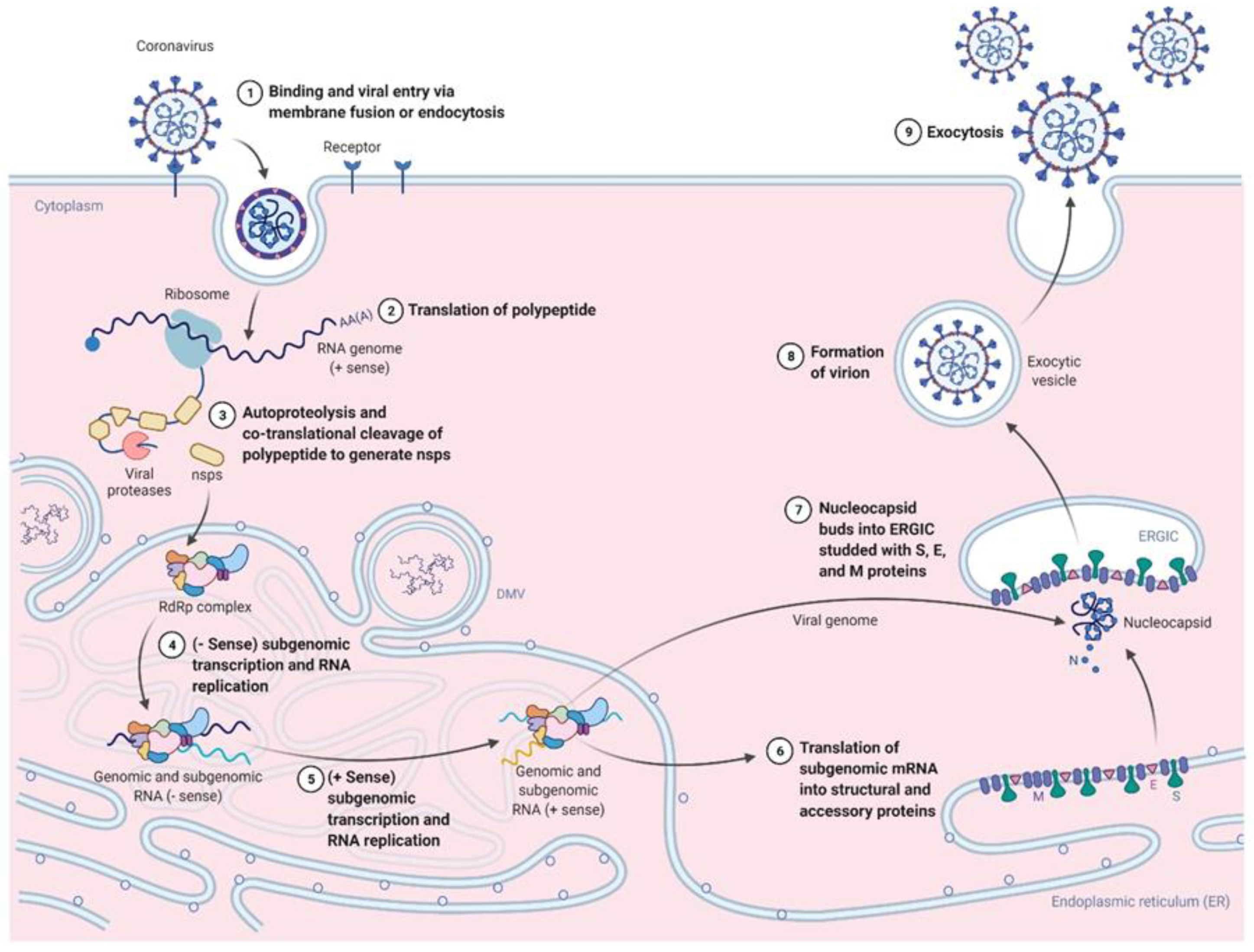{kind=link}
| Vaccine Types | Mechanisms | Benefits | Drawbacks | References |
|---|---|---|---|---|
| DNA vaccines (e.g., Inovio INO-4800) | -The DNA can penetrate the cell and produce target antigens through the host cell’s transcription and translation systems. | -Live viruses are not required for DNA vaccinations. -Plasmid DNA is made relatively easily, and compared with the viruses, double-stranded DNA molecules are more stable and, for storing long-term, can be freeze-dried. | -Their application may be limited due to the delivery method. It is challenging for the administered vaccine to reach the cell nucleus. Physical delivery methods are usually needed (e.g., electroporation). -They have the potential to integrate into the genome. | [3,4,5,6] |
| mRNA vaccines (e.g., Moderna mRNA-1273 and Pfizer-BioNTech BNT162b2) | -The mRNA can enter the cytoplasm and undergo translation in the host cells. | -Theoretically, they are safer since they do not directly interact with the host DNA. | -The vaccines’ immunological effect may be insufficient, and the vaccines are often delivered as part of a complex to increase efficacy. -They may need lower storage temperature. -Vaccine efficacy may be reduced by activation of an antiviral immune response involving interferons. | [3,4,7,8] |
| Non-replicating viral vector vaccines (e.g., Johnson & Johnson JNJ-78436735, Oxford-AstraZeneca ChAdOx1, Sputnik V Gam-COVID-Vac) | -Viral vectors can encode the antigens of interest in a different virus. The antigens mimic an infection in the body. | -The efficacy of these vaccines is fairly high. -They have been studied for decades. | -The manufacturing process is relatively complex. -Recombinant viruses have the ability to integrate into the host genome. -The body may have pre-existing immunity to the vector, which can decrease the immune response. | [3,4,9,10] |
| Inactivated vaccines (e.g., Sinopharm BBIBP-CorV and Sinovac CoronaVac) | -These vaccines can be created using chemical and radiation inactivation techniques, which result in the loss of viral pathogenicity. | -They can elicit strong immune responses. -They have been widely studied and are known to be relatively safe. | -The immunogenic epitopes may be altered during inactivation. -Inactivated vaccines for SARS coronavirus have been associated with lung pathology involving eosinophils. | [3,10] |
| Live attenuated vaccines | -These vaccines have diminished pathogenicity of the virus through mutations or deletions in the viral genome. | -They mimic natural infections and have high immunogenicity. | -They could still be pathogenic in the host, especially in the immunocompromised. | [4,10] |
| Subunit vaccines (e.g., Novavax NVX-CoV2373) | -They use fragments of viral antigens created from recombinant protein methods to elicit an immune response. | -They have a high level of safety. -They are relatively simple to produce. | -They have relatively lower immunogenicity. Thus, they are often used with adjuvants. | [4,10] |
| Vaccine Name | Vaccine Type [10] | Platform | Original Dose | Storage Temperature [14,15,16] |
|---|---|---|---|---|
| Moderna (mRNA-1273) | mRNA vaccine | mRNA in lipid nanoparticle | 2 | −50 °C to −15 °C (up to 9 months) 2° to 8 °C (up to 30 days) |
| Pfizer-BioNTech (BNT162b2) | mRNA vaccine | mRNA in lipid nanoparticle | 2 | Formulation for 12 years or older: −90 °C to −60 °C (up to 9 months) −25 °C to −15 °C (up to 2 weeks) 2 °C to 8 °C (up to 31 days after thawing) |
| Johnson & Johnson (JNJ-78436735) | Non-replicating viral vector vaccine | Non-replicating human adenovirus | 1 | 2 °C to 8 °C |
| Oxford–AstraZeneca (ChAdOx1) | Non-replicating viral vector vaccine | Non-replicating chimpanzee adenovirus | 2 | 2 °C to 8 °C |
| Sputnik V (Gam-COVID-Vac) | Non-replicating viral vector vaccine | Non-replicating human adenovirus | 2 | −18 °C |
| Sinopharm (BBIBP-CorV) | Inactivated vaccine | Inactivated virus | 2 | 2 °C to 8 °C |
| Sinovac (CoronaVac) | Inactivated vaccine | Inactivated virus | 2 | 2 °C to 8 °C |
| Novavax (NVX-CoV2373) | Subunit vaccine | Spike protein and Matrix M adjuvant | 2 | 2 °C to 8 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepand, M.R.; Bigdelou, B.; Ho, J.Q.; Sharaf, M.; Lannigan, A.J.; Sullivan, I.M.; da Silva, A.P.; Barrett, L.O.; McGoldrick, S.; Lnu, Y.; et al. Long-Term Immunity and Antibody Response: Challenges for Developing Efficient COVID-19 Vaccines. Antibodies 2022, 11, 35. https://doi.org/10.3390/antib11020035
Sepand MR, Bigdelou B, Ho JQ, Sharaf M, Lannigan AJ, Sullivan IM, da Silva AP, Barrett LO, McGoldrick S, Lnu Y, et al. Long-Term Immunity and Antibody Response: Challenges for Developing Efficient COVID-19 Vaccines. Antibodies. 2022; 11(2):35. https://doi.org/10.3390/antib11020035
Chicago/Turabian StyleSepand, Mohammad Reza, Banafsheh Bigdelou, Jim Q. Ho, Mohammad Sharaf, Alexis J. Lannigan, Ian M. Sullivan, Alecsander P. da Silva, Leland O. Barrett, Scott McGoldrick, Yuvraj Lnu, and et al. 2022. "Long-Term Immunity and Antibody Response: Challenges for Developing Efficient COVID-19 Vaccines" Antibodies 11, no. 2: 35. https://doi.org/10.3390/antib11020035
APA StyleSepand, M. R., Bigdelou, B., Ho, J. Q., Sharaf, M., Lannigan, A. J., Sullivan, I. M., da Silva, A. P., Barrett, L. O., McGoldrick, S., Lnu, Y., Lynch, S. E., Boisclair, J. M., Barnard-Pratt, D. D., & Zanganeh, S. (2022). Long-Term Immunity and Antibody Response: Challenges for Developing Efficient COVID-19 Vaccines. Antibodies, 11(2), 35. https://doi.org/10.3390/antib11020035