
New Opportunities in Glycan Engineering for Therapeutic Proteins


{kind=link}
{kind=link}
Abstract
:1. Introduction
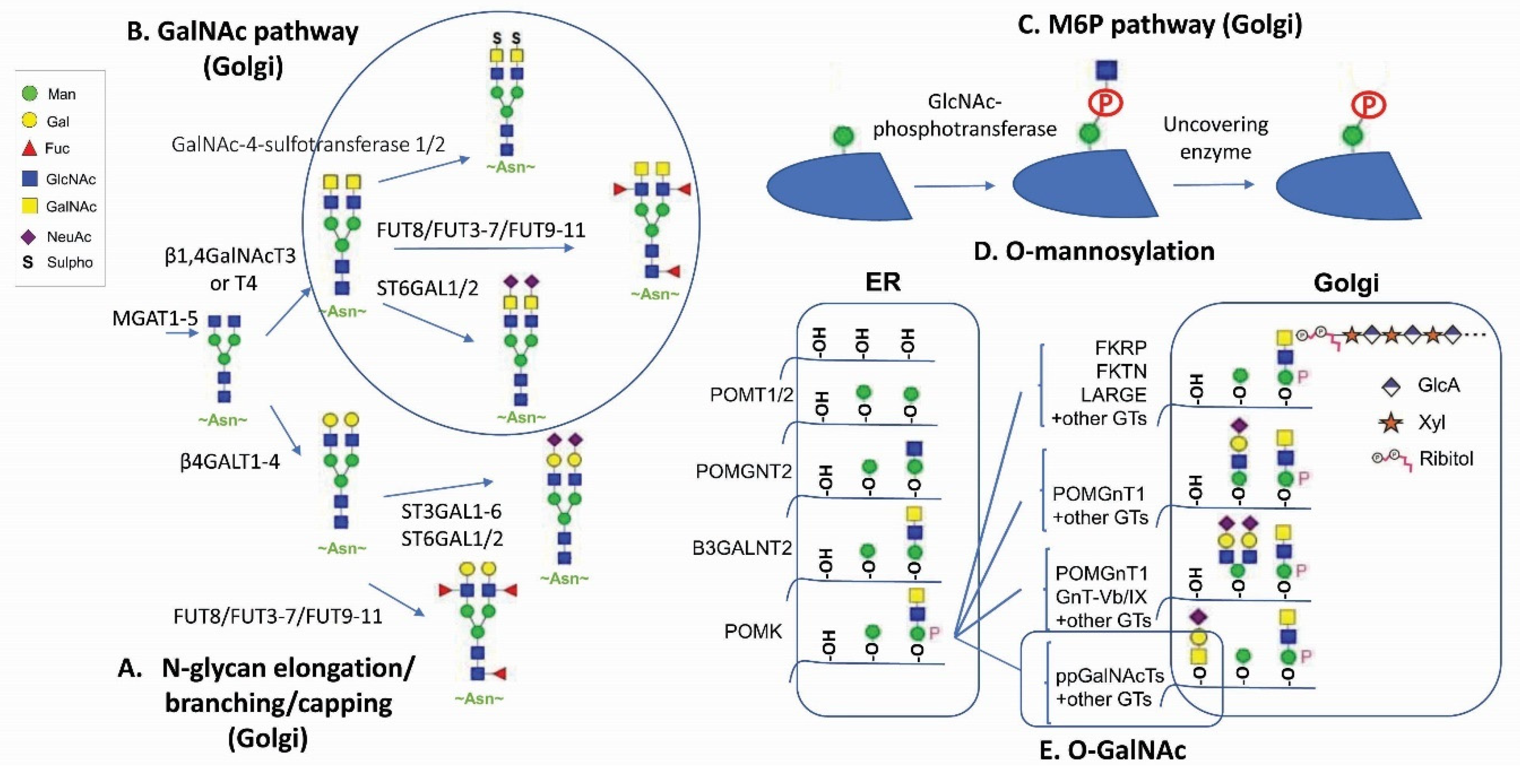
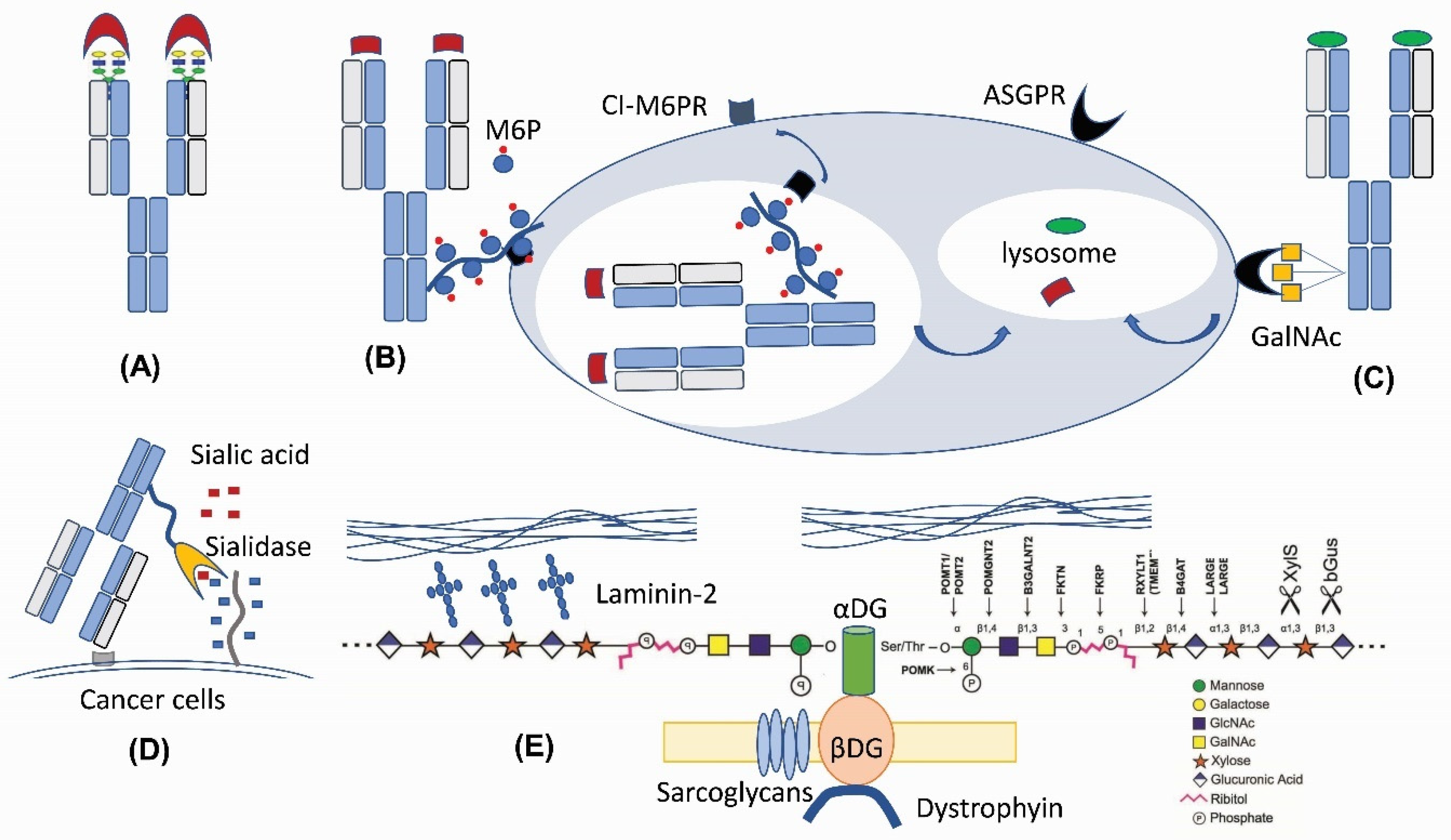
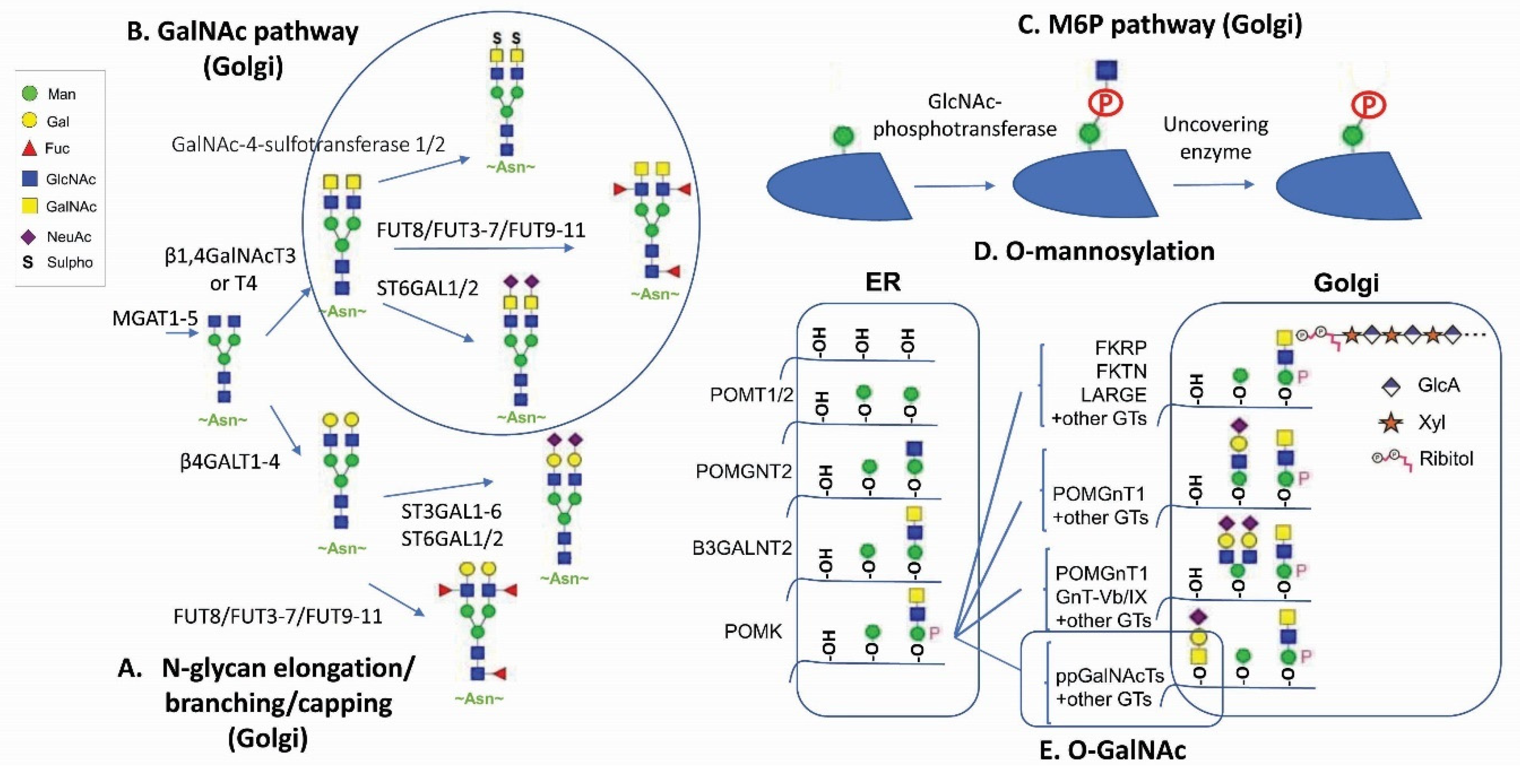
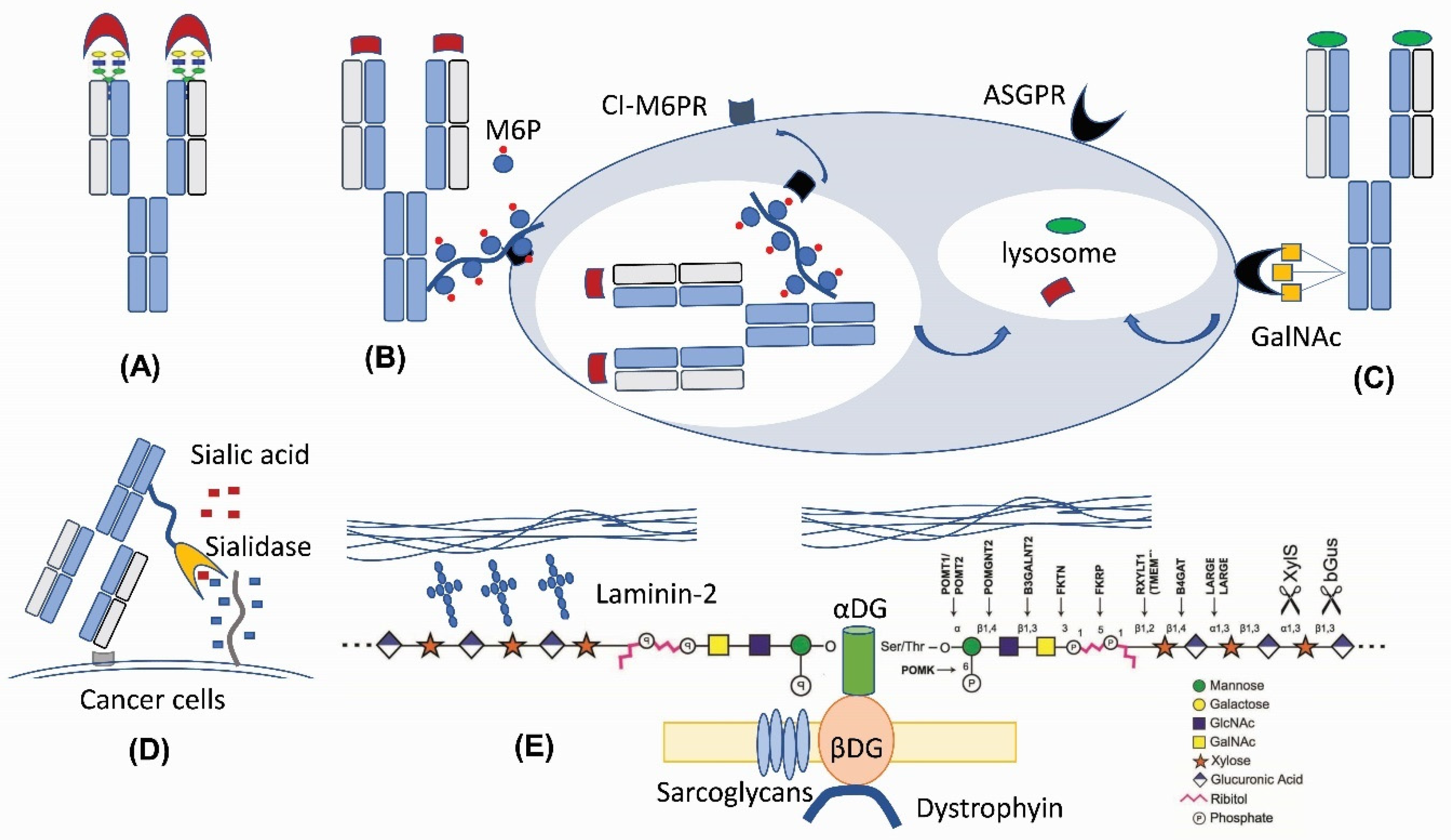
2. Glycans as an Unconventional Strategy for Antibody Diversification
3. GalNAc Binder—A New Application Based on Previous Findings
4. M6P—A Lysosomal Route for Non-Lysosomal Enzymes
5. O-Linked Glycan: New Tricks for an Old Player for Biological Systems
6. Glycoengineering as a Continued Theme for Biotherapeutics Applications
7. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; Aebi, M.; Darvill, A.G.; Kinoshita, T.; Packer, N.H.; Prestegard, J.H.; et al. Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017. [Google Scholar]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [Green Version]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.A.; Pedram, K.; Malaker, S.A.; Batista, P.J.; Smith, B.A.; Johnson, A.G.; George, B.M.; Majzoub, K.; Villalta, P.W.; Carette, J.E.; et al. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell 2021, 184, 3109–3124.e22. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, Y.; Büll, C.; Chen, Y.-H.; Wandall, H.H.; Yang, Z.; Clausen, H. Genetic glycoengineering in mammalian cells. J. Biol. Chem. 2021, 296, 100448. [Google Scholar] [CrossRef] [PubMed]
- Fournet, M.; Bonté, F.; Desmoulière, A. Glycation Damage: A Possible Hub for Major Pathophysiological Disorders and Aging. Aging Dis. 2018, 9, 880–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slawson, C.; Hart, G.W. O-GlcNAc signalling: Implications for cancer cell biology. Nat. Rev. Cancer 2011, 11, 678–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornfeld, R.; Kornfeld, S. Assembly of asparagine-linked oligosaccharides. Annu. Rev. Biochem. 1985, 54, 631–664. [Google Scholar] [CrossRef]
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nat. Rev. Drug Discov. 2009, 8, 226–234. [Google Scholar] [CrossRef]
- Hirata, T.; Kizuka, Y. N-Glycosylation. Adv. Exp. Med. Biol. 2021, 1325, 3–24. [Google Scholar]
- Donini, R.; Haslam, S.M.; Kontoravdi, C. Glycoengineering Chinese hamster ovary cells: A short history. Biochem. Soc. Trans. 2021, 49, 915–931. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Werner, A. Sweet Rules: Linking Glycosylation to Antibody Function. Exp. Suppl. 2021, 112, 365–393. [Google Scholar]
- Manzella, S.M.; Hooper, L.V.; Baenziger, J.U. Oligosaccharides Containing β1,4-Linked N-Acetylgalactosamine, a Paradigm for Protein-specific Glycosylation. J. Biol. Chem. 1996, 271, 12117–12120. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, M.F.; Prata, M.J.; Alves, S. Mannose-6-phosphate pathway: A review on its role in lysosomal function and dysfunction. Mol. Genet. Metab. 2012, 105, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Yoshida-Moriguchi, T.; Campbell, K.P. Matriglycan: A novel polysaccharide that links dystroglycan to the basement membrane. Glycobiology 2015, 25, 702–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanyavuz, A.; Marey-Jarossay, A.; Lacroix-Desmazes, S.; Dimitrov, J.D. Breaking the law: Unconventional strategies for antibody diversification. Nat. Rev. Immunol. 2019, 19, 355–368. [Google Scholar] [CrossRef]
- Banik, S.M.; Pedram, K.; Wisnovsky, S.; Ahn, G.; Riley, N.M.; Bertozzi, C.R. Lysosome-targeting chimaeras for degradation of extracellular proteins. Nature 2020, 584, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Ahn, G.; Banik, S.M.; Miller, C.L.; Riley, N.M.; Cochran, J.R.; Bertozzi, C.R. LYTACs that engage the asialoglycoprotein receptor for targeted protein degradation. Nat. Chem. Biol. 2021, 17, 937–946. [Google Scholar] [CrossRef]
- Caianiello, D.F.; Zhang, M.; Ray, J.D.; Howell, R.A.; Swartzel, J.C.; Branham, E.M.J.; Chirkin, E.; Sabbasani, V.R.; Gong, A.Z.; McDonald, D.M.; et al. Bifunctional small molecules that mediate the degradation of extracellular proteins. Nat. Chem. Biol. 2021, 17, 947–953. [Google Scholar] [CrossRef]
- Zhou, Y.; Teng, P.; Montgomery, N.T.; Li, X.; Tang, W. Development of Triantennary N-Acetylgalactosamine Conjugates as Degraders for Extracellular Proteins. ACS Central Sci. 2021, 7, 499–506. [Google Scholar] [CrossRef]
- Gray, M.A.; Stanczak, M.A.; Mantuano, N.R.; Xiao, H.; Pijnenborg, J.F.A.; Malaker, S.A.; Miller, C.L.; Weidenbacher, P.A.; Tanzo, J.T.; Ahn, G.; et al. Targeted glycan degradation potentiates the anticancer immune response in vivo. Nat. Chem. Biol. 2020, 16, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Woods, E.C.; Vukojicic, P.; Bertozzi, C.R. Precision glycocalyx editing as a strategy for cancer immunotherapy. Proc. Natl. Acad. Sci. USA 2016, 113, 10304–10309. [Google Scholar] [CrossRef] [Green Version]
- Reed, U.C. Congenital muscular dystrophy. Part II: A review of pathogenesis and therapeutic perspectives. Arq. Neuro-Psiquiatr. 2009, 67, 343–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, M.; Venzke, D.; Anderson, M.E.; Yoshida-Moriguchi, T.; Glushka, J.N.; Nairn, A.V.; Galizzi, M.; Moremen, K.W.; Campbell, K.P.; Wells, L. HNK-1 sulfotransferase modulates α-dystroglycan glycosylation by 3-O-sulfation of glucuronic acid on matriglycan. Glycobiology 2020, 30, 817–829. [Google Scholar] [CrossRef] [PubMed]
- van de Bovenkamp, F.S.; Hafkenscheid, L.; Rispens, T.; Rombouts, Y. The Emerging Importance of IgG Fab Glycosylation in Immunity. J. Immunol. 2016, 196, 1435–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondt, A.; Rombouts, Y.; Selman, M.H.J.; Hensbergen, P.; Reiding, K.R.; Hazes, J.M.W.; Dolhain, R.J.E.M.; Wuhrer, M. Immunoglobulin G (IgG) Fab Glycosylation Analysis Using a New Mass Spectrometric High-throughput Profiling Method Reveals Pregnancy-associated Changes. Mol. Cell. Proteom. 2014, 13, 3029–3039. [Google Scholar] [CrossRef] [Green Version]
- van de Bovenkamp, F.S.; Derksen, N.I.; Ooijevaar-de Heer, P.; van Schie, K.A.; Kruithof, S.; Berkowska, M.A.; van der Schoot, C.E.; IJspeert, H.; van der Burg, M.; Gils, A. Adaptive antibody diversification through N-linked glycosylation of the immunoglobulin variable region. Proc. Natl. Acad. Sci. USA 2018, 115, 1901–1906. [Google Scholar] [CrossRef] [Green Version]
- van de Bovenkamp, F.S.; Derksen, N.I.; van Breemen, M.J.; de Taeye, S.W.; Ooijevaar-de Heer, P.; Sanders, R.W.; Rispens, T. Variable Domain N-Linked Glycans Acquired During Antigen-Specific Immune Responses Can Contribute to Immunoglobulin G Antibody Stability. Front. Immunol. 2018, 9, 740. [Google Scholar] [CrossRef]
- Lefranc, M.P. IMGT, the International ImMunoGeneTics Information System. Cold Spring Harb. Protoc. 2011, 2011, 595–603. [Google Scholar] [CrossRef]
- Song, R.; Oren, D.A.; Franco, D.; Seaman, M.S.; Ho, D.D. Strategic addition of an N-linked glycan to a monoclonal antibody improves its HIV-1–neutralizing activity. Nat. Biotechnol. 2013, 31, 1047–1052. [Google Scholar] [CrossRef] [Green Version]
- Jacquemin, M.; Radcliffe, C.M.; Lavend’Homme, R.; Wormald, M.; VanderElst, L.; Wallays, G.; Dewaele, J.; Collen, D.; Vermylen, J.; Dwek, R.A.; et al. Variable region heavy chain glycosylation determines the anticoagulant activity of a factor VIII antibody. J. Thromb. Haemost. 2006, 4, 1047–1055. [Google Scholar] [CrossRef]
- Koers, J.; Derksen, N.I.L.; Heer, P.O.-D.; Nota, B.; Van De Bovenkamp, F.S.; Vidarsson, G.; Rispens, T. Biased N-Glycosylation Site Distribution and Acquisition across the Antibody V Region during B Cell Maturation. J. Immunol. 2019, 202, 2220–2228. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, Y.; Willemze, A.; van Beers, J.J.; Shi, J.; Kerkman, P.F.; van Toorn, L.; Janssen, G.M.; Zaldumbide, A.; Hoeben, R.C.; Pruijn, G.J.; et al. Extensive glycosylation of ACPA-IgG variable domains modulates binding to citrullinated antigens in rheumatoid arthritis. Ann. Rheum. Dis. 2016, 75, 578–585. [Google Scholar] [CrossRef]
- Hafkenscheid, L.; Bondt, A.; Scherer, H.U.; Huizinga, T.W.J.; Wuhrer, M.; Toes, R.; Rombouts, Y. Structural Analysis of Variable Domain Glycosylation of Anti-Citrullinated Protein Antibodies in Rheumatoid Arthritis Reveals the Presence of Highly Sialylated Glycans. Mol. Cell. Proteom. 2017, 16, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Vletter, E.M.; Koning, M.T.; Scherer, H.U.; Veelken, H.; Toes, R.E.M. A Comparison of Immunoglobulin Variable Region N-Linked Glycosylation in Healthy Donors, Autoimmune Disease and Lymphoma. Front. Immunol. 2020, 11, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallick, S.C.; Kabat, E.A.; Morrison, S.L. Glycosylation of a VH residue of a monoclonal antibody against alpha (1----6) dextran increases its affinity for antigen. J. Exp. Med. 1988, 168, 1099–1109. [Google Scholar] [CrossRef]
- Khurana, S.; Raghunathan, V.; Salunke, D.M. The Variable Domain Glycosylation in a Monoclonal Antibody Specific to GnRH Modulates Antigen Binding. Biochem. Biophys. Res. Commun. 1997, 234, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Coloma, M.J.; Trinh, R.K.; Martinez, A.R.; Morrison, S.L. Position effects of variable region carbohydrate on the affinity and in vivo behavior of an anti-(1→6) dextran antibody. J. Immunol. 1999, 162, 2162–2170. [Google Scholar] [PubMed]
- Leibiger, H.; Wüstner, D.; Stigler, R.-D.; Marx, U. Variable domain-linked oligosaccharides of a human monoclonal IgG: Structure and influence on antigen binding. Biochem. J. 1999, 338, 529–538. [Google Scholar] [CrossRef]
- Culver, E.L.; Van De Bovenkamp, F.S.; Derksen, N.I.L.; Koers, J.; Cargill, T.; Barnes, E.; de Neef, L.; Koeleman, C.A.M.; Aalberse, R.C.; Wuhrer, M.; et al. Unique patterns of glycosylation in immunoglobulin subclass G4-related disease and primary sclerosing cholangitis. J. Gastroenterol. Hepatol. 2018, 34, 1878–1886. [Google Scholar] [CrossRef] [PubMed]
- Sabouri, Z.; Schofield, P.; Horikawa, K.; Spierings, E.; Kipling, D.; Randall, K.L.; Langley, D.; Roome, B.; Vazquez-Lombardi, R.; Rouet, R.; et al. Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity. Proc. Natl. Acad. Sci. USA 2014, 111, E2567–E2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, G.-Y.; Zhang, B.; McKee, K.; O’Dell, S.; Kwon, Y.D.; Zhou, T.; Blinn, J.; Lloyd, K.; Parks, R.; Von Holle, T.; et al. Eliminating antibody polyreactivity through addition ofN-linked glycosylation. Protein Sci. 2015, 24, 1019–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.-J.; Luo, J.; O’Neil, K.T.; Kang, J.; Lacy, E.R.; Canziani, G.; Baker, A.; Huang, M.; Tang, Q.M.; Raju, T.; et al. Structure-based engineering of a monoclonal antibody for improved solubility. Protein Eng. Des. Sel. 2010, 23, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Pepinsky, R.B.; Silvian, L.; Berkowitz, S.A.; Farrington, G.; Lugovskoy, A.; Walus, L.; Eldredge, J.; Capili, A.; Mi, S.; Graff, C.; et al. Improving the solubility of anti-LINGO-1 monoclonal antibody Li33 by isotype switching and targeted mutagenesis. Protein Sci. 2010, 19, 954–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, Y.; Lin, A.; Fiete, D.; Steirer, L.; Baenziger, J.U. Modulation of Mannose and Asialoglycoprotein Receptor Expression Determines Glycoprotein Hormone Half-life at Critical Points in the Reproductive Cycle. J. Biol. Chem. 2014, 289, 12157–12167. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.; Fiete, D.; Blake, N.M.J.; Beranek, M.; Oates, E.L.; Mi, Y.; Roseman, D.S.; Baenziger, J.U. A Necessary and Sufficient Determinant for Protein-selective Glycosylation in Vivo. J. Biol. Chem. 2008, 283, 1985–1991. [Google Scholar] [CrossRef] [Green Version]
- Kawar, Z.S.; Haslam, S.M.; Morris, H.R.; Dell, A.; Cummings, R.D. Novel poly-GalNAcbeta1-4GlcNAc (LacdiNAc) and fucosylated poly-LacdiNAc N-glycans from mammalian cells expressing beta1,4-N-acetylgalactosaminyltransferase and alpha1,3-fucosyltransferase. J. Biol. Chem. 2005, 280, 12810–12819. [Google Scholar] [CrossRef] [Green Version]
- Nimtz, M.; Conradt, H.S.; Mann, K. LacdiNAc (GalNAcbeta1-4GlcNAc) is a major motif in N-glycan structures of the chicken eggshell protein ovocleidin-116. Biochim. Biophys. Acta 2004, 1675, 71–80. [Google Scholar] [CrossRef]
- Bonar, D.; Hanisch, F.-G. Trefoil Factor Family Domains Represent Highly Efficient Conformational Determinants for N-Linked N,N′-di-N-acetyllactosediamine (LacdiNAc) Synthesis. J. Biol. Chem. 2014, 289, 29677–29690. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Gotoh, M.; Kiyohara, K.; Kameyama, A.; Kubota, T.; Kikuchi, N.; Ishizuka, Y.; Iwasaki, H.; Togayachi, A.; Kudo, T.; et al. Molecular cloning and characterization of a novel human beta 1,4-N-acetylgalactosaminyltransferase, beta 4GalNAc-T3, responsible for the synthesis of N,N’-diacetyllactosediamine, galNAc beta 1-4GlcNAc. J. Biol. Chem. 2003, 278, 47534–47544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, M.; Sato, T.; Kiyohara, K.; Kameyama, A.; Kikuchi, N.; Kwon, Y.D.; Ishizuka, Y.; Iwai, T.; Nakanishi, H.; Narimatsu, H. Molecular cloning and characterization of beta1,4-N-acetylgalactosaminyltransferases IV synthesizing N,N′-diacetyllactosediamine. FEBS Lett. 2004, 562, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Mengeling, B.J.; Manzella, S.M.; Baenziger, J.U. A cluster of basic amino acids within an alpha-helix is essential for alpha-subunit recognition by the glycoprotein hormone N-acetylgalactosaminyltransferase. Proc. Natl. Acad. Sci. USA 1995, 92, 502–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Jagarlapudi, S.; Weng, Y.; Ly, M.; Rouse, J.C.; McClure, K.; Ishino, T.; Zhang, Y.; Sousa, E.; Cohen, J.; et al. Structure-function relationships of the soluble form of the antiaging protein Klotho have therapeutic implications for managing kidney disease. J. Biol. Chem. 2020, 295, 3115–3133. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.; Ng, S.K. Impact of host cell line choice on glycan profile. Crit. Rev. Biotechnol. 2017, 38, 851–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.L.; Goh, J.; Srinivasan, H.; Liu, K.I.; Gowher, A.; Shanmugam, R.; Lim, H.L.; Choo, M.; Tang, W.Q.; Tan, A.H.-M.; et al. A human expression system based on HEK293 for the stable production of recombinant erythropoietin. Sci. Rep. 2019, 9, 116768. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.; Dahms, N.M.; Kornfeld, S. Mannose 6-phosphate receptors: New twists in the tale. Nat. Rev. Mol. Cell Biol. 2003, 4, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, R. Treatments for lysosomal storage disorders. Biochem. Soc. Trans. 2010, 38, 1465–1468. [Google Scholar] [CrossRef]
- Gary-Bobo, M.; Nirde, P.; Jeanjean, A.; Morere, A.; Garcia, M. Mannose 6-Phosphate Receptor Targeting and its Applications in Human Diseases. Curr. Med. Chem. 2007, 14, 2945–2953. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, X.; Kyazike, J.; Zhou, Q.; Thurberg, B.L.; Raben, N.; Mattaliano, R.J.; Cheng, S.H. Conjugation of Mannose 6-Phosphate-containing Oligosaccharides to Acid α-Glucosidase Improves the Clearance of Glycogen in Pompe Mice. J. Biol. Chem. 2004, 279, 50336–50341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, C.R.; Burke, R.L.; Kornfeld, S.; Gregory, W.; Masiarz, F.R.; Dingwell, K.S.; Johnson, D.C. Herpes simplex virus glycoprotein D acquires mannose 6-phosphate residues and binds to mannose 6-phosphate receptors. J. Biol. Chem. 1994, 269, 17067–17074. [Google Scholar] [CrossRef]
- Brunetti, C.R.; Burke, R.L.; Hoflack, B.; Ludwig, T.; Dingwell, K.S.; Johnson, D.C. Role of mannose-6-phosphate receptors in herpes simplex virus entry into cells and cell-to-cell transmission. J. Virol. 1995, 69, 3517–3528. [Google Scholar] [CrossRef] [Green Version]
- Tiels, P.; Baranova, E.; Piens, K.; De Visscher, C.; Pynaert, G.; Nerinckx, W.; Stout, J.; Fudalej, F.; Hulpiau, P.; Tännler, S.; et al. A bacterial glycosidase enables mannose-6-phosphate modification and improved cellular uptake of yeast-produced recombinant human lysosomal enzymes. Nat. Biotechnol. 2012, 30, 1225–1231. [Google Scholar] [CrossRef]
- Kato, K.; Jeanneau, C.; Tarp, M.A.; Benet-Pagès, A.; Lorenz-Depiereux, B.; Bennett, E.P.; Mandel, U.; Strom, T.M.; Clausen, H. Polypeptide GalNAc-transferase T3 and familial tumoral calcinosis. Secretion of fibroblast growth factor 23 requires O-glycosylation. J. Biol. Chem. 2006, 281, 18370–18377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Mao, Y.; Narimatsu, Y.; Ye, Z.; Tian, W.; Goth, C.; Lira-Navarrete, E.; Pedersen, N.B.; Benito-Vicente, A.; Martin, C.; et al. Site-specific O-glycosylation of members of the low-density lipoprotein receptor superfamily enhances ligand interactions. J. Biol. Chem. 2018, 293, 7408–7422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Gomathinayagam, S.; Hamuro, L.; Prueksaritanont, T.; Wang, W.; Stadheim, T.A.; Hamilton, S.R. The Impact of Glycosylation on the Pharmacokinetics of a TNFR2:Fc Fusion Protein Expressed in Glycoengineered Pichia Pastoris. Pharm. Res. 2013, 30, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Goth, C.; Halim, A.; Khetarpal, S.A.; Rader, D.J.; Clausen, H.; Schjoldager, K.T.-B.G. A systematic study of modulation of ADAM-mediated ectodomain shedding by site-specific O-glycosylation. Proc. Natl. Acad. Sci. USA 2015, 112, 14623–14628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goth, C.K.; Tuhkanen, H.E.; Khan, H.; Lackman, J.J.; Wang, S.; Narimatsu, Y.; Hansen, L.H.; Overall, C.M.; Clausen, H.; Schjoldager, K.T.; et al. Site-specific O-Glycosylation by Polypeptide N-Acetylgalactosaminyltransferase 2 (GalNAc-transferase T2) Co-regulates beta1-Adrenergic Receptor N-terminal Cleavage. J. Biol. Chem. 2017, 292, 4714–4726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, L.H.; Madsen, T.D.; Goth, C.K.; Clausen, H.; Chen, Y.; Dzhoyashvili, N.; Iyer, S.R.; Sangaralingham, S.J.; Burnett, J.C.; Rehfeld, J.F.; et al. Discovery of O-glycans on atrial natriuretic peptide (ANP) that affect both its proteolytic degradation and potency at its cognate receptor. J. Biol. Chem. 2019, 294, 12567–12578. [Google Scholar] [CrossRef] [Green Version]
- Holdener, B.C.; Haltiwanger, R.S. Protein O-fucosylation: Structure and function. Curr. Opin. Struct. Biol. 2019, 56, 78–86. [Google Scholar] [CrossRef]
- Takeuchi, H.; Yu, H.; Hao, H.; Takeuchi, M.; Ito, A.; Li, H.; Haltiwanger, R.S. O-Glycosylation modulates the stability of epidermal growth factor-like repeats and thereby regulates Notch trafficking. J. Biol. Chem. 2017, 292, 15964–15973. [Google Scholar] [CrossRef] [Green Version]
- Madsen, T.D.; Hansen, L.H.; Hintze, J.; Ye, Z.; Jebari, S.; Andersen, D.; Joshi, H.J.; Ju, T.; Goetze, J.P.; Martin, C.; et al. An atlas of O-linked glycosylation on peptide hormones reveals diverse biological roles. Nat. Commun. 2020, 11, 14033. [Google Scholar] [CrossRef]
- Zhong, X.; Kieras, E.; Sousa, E.; D’Antona, A.; Baber, J.C.; He, T.; Desharnais, J.; Wood, L.; Luxenberg, D.; Stahl, M.; et al. Pyroglutamate and O-Linked Glycan Determine Functional Production of Anti-IL17A and Anti-IL22 Peptide-Antibody Bispecific Genetic Fusions. J. Biol. Chem. 2013, 288, 1409–1419. [Google Scholar] [CrossRef] [Green Version]
- Larsen, I.S.B.; Narimatsu, Y.; Joshi, H.J.; Siukstaite, L.; Harrison, O.J.; Brasch, J.; Goodman, K.M.; Hansen, L.; Shapiro, L.; Honig, B.; et al. Discovery of an O-mannosylation pathway selectively serving cadherins and protocadherins. Proc. Natl. Acad. Sci. USA 2017, 114, 11163–11168. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, M.O.; Capicciotti, C.J.; Liu, L.; Praissman, J.; Mead, D.G.; Brindley, M.A.; Willer, T.; Campbell, K.P.; Moremen, K.W.; Wells, L.; et al. Cell Surface Glycan Engineering Reveals that Matriglycan Alone can Recapitulate Dystroglycan Binding and Function. bioRxiv 2021. [Google Scholar] [CrossRef]
- Gumlaw, N.; Sevigny, L.M.; Zhao, H.; Luo, Z.; Bangari, D.S.; Masterjohn, E.; Chen, Y.; McDonald, B.; Magnay, M.; Travaline, T.; et al. biAb Mediated Restoration of the Linkage between Dystroglycan and Laminin-211 as a Therapeutic Approach for α-Dystroglycanopathies. Mol. Ther. 2019, 28, 664–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inamori, K.-I.; Yoshida-Moriguchi, T.; Hara, Y.; Anderson, M.E.; Yu, L.; Campbell, K.P. Dystroglycan Function Requires Xylosyl- and Glucuronyltransferase Activities of LARGE. Science 2012, 335, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, R.; Kanagawa, M.; Yoshida-Moriguchi, T.; Rader, E.P.; Ng, R.A.; Michele, D.E.; Muirhead, D.E.; Kunz, S.; Moore, S.; Iannaccone, S.T.; et al. Basal lamina strengthens cell membrane integrity via the laminin G domain-binding motif of -dystroglycan. Proc. Natl. Acad. Sci. USA 2009, 106, 12573–12579. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, G.A.; Barton, N.W.; Pastores, G.; Dambrosia, J.M.; Banerjee, T.K.; McKee, M.A.; Parker, C.; Schiffmann, R.; Hill, S.C.; Brady, R.O. Enzyme Therapy in Type 1 Gaucher Disease: Comparative Efficacy of Mannose-Terminated Glucocerebrosidase from Natural and Recombinant Sources. Ann. Intern. Med. 1995, 122, 33–39. [Google Scholar] [CrossRef]
- Steirer, L.M.; Park, E.I.; Townsend, R.R.; Baenziger, J.U. The Asialoglycoprotein Receptor Regulates Levels of Plasma Glycoproteins Terminating with Sialic Acid α2,6-Galactose. J. Biol. Chem. 2009, 284, 3777–3783. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, H.M.; Christensen, E.I.; Appa, R.S.; Seested, T. The unsialylated subpopulation of recombinant activated factor VII binds to the asialo-glycoprotein receptor (ASGPR) on primary rat hepatocytes. Thromb. Haemost. 2010, 104, 1166–1173. [Google Scholar] [CrossRef]
- Tian, W.; Ye, Z.; Wang, S.; Schulz, M.A.; Van Coillie, J.; Sun, L.; Chen, Y.-H.; Narimatsu, Y.; Hansen, L.; Kristensen, C.; et al. The glycosylation design space for recombinant lysosomal replacement enzymes produced in CHO cells. Nat. Commun. 2019, 10, 1785. [Google Scholar] [CrossRef] [Green Version]
- Elliott, S.; Lorenzini, T.; Asher, S.; Aoki, K.; Brankow, D.; Buck, L.; Busse, L.; Chang, D.; Fuller, J.; Grant, J.; et al. Enhancement of therapeutic protein in vivo activities through glycoengineering. Nat. Biotechnol. 2003, 21, 414–421. [Google Scholar] [CrossRef]
- Stabach, P.R.; Zimmerman, K.; Adame, A.; Kavanagh, D.; Saeui, C.T.; Agatemor, C.; Gray, S.; Cao, W.; De La Cruz, E.M.; Yarema, K.J.; et al. Improving the Pharmacodynamics and In Vivo Activity of ENPP1-Fc Through Protein and Glycosylation Engineering. Clin. Transl. Sci. 2020, 14, 362–372. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Ravetch, J.V. Anti-Inflammatory Actions of Intravenous Immunoglobulin. Annu. Rev. Immunol. 2008, 26, 513–533. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.M.; Nimmerjahn, F.; Ashline, D.J.; Reinhold, V.N.; Paulson, J.C.; Ravetch, J.V. Recapitulation of IVIG Anti-Inflammatory Activity with a Recombinant IgG Fc. Science 2008, 320, 373–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.T.; Ravetch, J.V. Functional diversification of IgGs through Fc glycosylation. J. Clin. Investig. 2019, 129, 3492–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagan, J.D.; Kitaoka, M.; Anthony, R.M. Engineered Sialylation of Pathogenic Antibodies In Vivo Attenuates Autoimmune Disease. Cell 2017, 172, 564–577.e13. [Google Scholar] [CrossRef] [Green Version]
- Yin, B.; Gao, Y.; Chung, C.-Y.; Yang, S.; Blake, E.; Stuczynski, M.C.; Tang, J.; Kildegaard, H.F.; Andersen, M.R.; Zhang, H.; et al. Glycoengineering of Chinese hamster ovary cells for enhanced erythropoietin N-glycan branching and sialylation. Biotechnol. Bioeng. 2015, 112, 2343–2351. [Google Scholar] [CrossRef] [PubMed]
- Shade, K.-T.C.; Conroy, M.E.; Washburn, N.; Kitaoka, M.; Huynh, D.J.; Laprise, E.; Patil, S.U.; Shreffler, W.G.; Anthony, R.M. Sialylation of immunoglobulin E is a determinant of allergic pathogenicity. Nature 2020, 582, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Mirakhur, B.; Chan, E.; Le, Q.-T.; Berlin, J.; Morse, M.; Murphy, B.A.; Satinover, S.M.; Hosen, J.; Mauro, D.; et al. Cetuximab-Induced Anaphylaxis and IgE Specific for Galactose-α-1,3-Galactose. N. Engl. J. Med. 2008, 358, 1109–1117. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, E.; Macauley, M.S. Targeted self-destruction. Nat. Chem. Biol. 2020, 16, 1281–1283. [Google Scholar] [CrossRef]
- Zhong, X.; Somers, W. Recent Advances in Glycosylation Modifications in the Context of Therapeutic Glycoproteins. In Integrative Proteomics; Leung, E., Ed.; InTech: London, UK, 2012; pp. 183–196. [Google Scholar]
- Mondal, N.; Buffone, A.; Stolfa, G.; Antonopoulos, A.; Lau, J.T.Y.; Haslam, S.M.; Dell, A.; Neelamegham, S. ST3Gal-4 is the primary sialyltransferase regulating the synthesis of E-, P-, and L-selectin ligands on human myeloid leukocytes. Blood 2015, 125, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Baggen, J.; Thibaut, H.J.; Staring, J.; Jae, L.T.; Liu, Y.; Guo, H.; Slager, J.J.; de Bruin, J.W.; van Vliet, A.L.W.; Blomen, V.A.; et al. Enterovirus D68 receptor requirements unveiled by haploid genetics. Proc. Natl. Acad. Sci. USA 2016, 113, 1399–1404. [Google Scholar] [CrossRef] [Green Version]
- Qi, F.; Isaji, T.; Duan, C.; Yang, J.; Wang, Y.; Fukuda, T.; Gu, J. ST3GAL3, ST3GAL4, and ST3GAL6 differ in their regulation of biological functions via the specificities for the alpha2,3-sialylation of target proteins. FASEB J. 2020, 34, 881–897. [Google Scholar] [CrossRef] [Green Version]
- Haga, K.; Ettayebi, K.; Tenge, V.R.; Karandikar, U.C.; Lewis, M.A.; Lin, S.-C.; Neill, F.H.; Ayyar, B.V.; Zeng, X.-L.; Larson, G.; et al. Genetic Manipulation of Human Intestinal Enteroids Demonstrates the Necessity of a Functional Fucosyltransferase 2 Gene for Secretor-Dependent Human Norovirus Infection. mBio 2020, 11, e00251-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, M.J.; Shah, S.R.; Saeui, C.T.; Ariss, R.; Yarema, K.J. Improving Immunotherapy Through Glycodesign. Front. Immunol. 2018, 9, 2485. [Google Scholar] [CrossRef] [PubMed]
- Van Landuyt, L.; Lonigro, C.; Meuris, L.; Callewaert, N. Customized protein glycosylation to improve biopharmaceutical function and targeting. Curr. Opin. Biotechnol. 2018, 60, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Hudak, J.E.; Bertozzi, C.R. Glycotherapy: New Advances Inspire a Reemergence of Glycans in Medicine. Chem. Biol. 2013, 21, 16–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhu, Z.; Chen, W.; Feng, Y.; Dimitrov, D.S. Crystallizable Fragment Glycoengineering for Therapeutic Antibodies Development. Front. Immunol. 2017, 8, 1554. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Chung, C.-Y.; Chough, S.; Betenbaugh, M.J. Antibody glycoengineering strategies in mammalian cells. Biotechnol. Bioeng. 2018, 115, 1378–1393. [Google Scholar] [CrossRef]
- Yamane-Ohnuki, N.; Kinoshita, S.; Inoue-Urakubo, M.; Kusunoki, M.; Iida, S.; Nakano, R.; Wakitani, M.; Niwa, R.; Sakurada, M.; Uchida, K.; et al. Establishment ofFUT8 knockout Chinese hamster ovary cells: An ideal host cell line for producing completely defucosylated antibodies with enhanced antibody-dependent cellular cytotoxicity. Biotechnol. Bioeng. 2004, 87, 614–622. [Google Scholar] [CrossRef]
- Zhong, X.; Cooley, C.; Seth, N.; Juo, Z.S.; Presman, E.; Resendes, N.; Kumar, R.; Allen, M.; Mosyak, L.; Stahl, M.; et al. Engineering novel Lec1 glycosylation mutants in CHO-DUKX cells: Molecular insights and effector modulation of N-acetylglucosaminyltransferase I. Biotechnol. Bioeng. 2012, 109, 1723–1734. [Google Scholar] [CrossRef]
- Ferrara, C.; Brünker, P.; Suter, T.; Moser, S.; Püntener, U.; Umaña, P. Modulation of therapeutic antibody effector functions by glycosylation engineering: Influence of Golgi enzyme localization domain and co-expression of heterologous beta1, 4-N-acetylglucosaminyltransferase III and Golgi alpha-mannosidase II. Biotechnol. Bioeng. 2006, 93, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.M.; Kowle, R.L.; Lian, Z.; Strifler, B.A.; Witcher, D.R.; Parekh, B.S.; Wang, T.; Frye, C.C. Modulation of IgG1 immunoeffector function by glycoengineering of the GDP-fucose biosynthesis pathway. Biotechnol. Bioeng. 2017, 115, 705–718. [Google Scholar] [CrossRef] [PubMed]
- von Horsten, H.H.; Ogorek, C.; Blanchard, V.; Demmler, C.; Giese, C.; Winkler, K.; Kaup, M.; Berger, M.; Jordan, I.; Sandig, V. Production of non-fucosylated antibodies by co-expression of heterologous GDP-6-deoxy-D-lyxo-4-hexulose reductase. Glycobiology 2010, 20, 1607–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.F.; Shahreel, W.; Wan, C.; Teo, G.; Hayati, N.; Tay, S.J.; Tong, W.H.; Yang, Y.; Rudd, P.M.; Zhang, P.; et al. Inactivation of GDP-fucose transporter gene (Slc35c1) in CHO cells by ZFNs, TALENs and CRISPR-Cas9 for production of fucose-free antibodies. Biotechnol. J. 2015, 11, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.G.; Mujacic, M.; Frohn, M.J.; Pickrell, A.J.; Kodama, P.; Bagal, D.; Miguel, T.S.; Sickmier, E.A.; Osgood, S.; Swietlow, A.; et al. Facile Modulation of Antibody Fucosylation with Small Molecule Fucostatin Inhibitors and Cocrystal Structure with GDP-Mannose 4,6-Dehydratase. ACS Chem. Biol. 2016, 11, 2734–2743. [Google Scholar] [CrossRef]
- Burkart, M.D.; Vincent, S.P.; Düffels, A.; Murray, B.; Ley, S.; Wong, C.-H. Chemo-enzymatic synthesis of fluorinated sugar nucleotide: Useful mechanistic Probes for glycosyltransferases. Bioorg. Med. Chem. 2000, 8, 1937–1946. [Google Scholar] [CrossRef]
- Okeley, N.M.; Alley, S.C.; Anderson, M.E.; Boursalian, T.E.; Burke, P.J.; Emmerton, K.M.; Jeffrey, S.C.; Klussman, K.; Law, C.-L.; Sussman, D.; et al. Development of orally active inhibitors of protein and cellular fucosylation. Proc. Natl. Acad. Sci. USA 2013, 110, 5404–5409. [Google Scholar] [CrossRef] [Green Version]
- Rillahan, C.D.; Antonopoulos, A.; Lefort, C.T.; Sonon, R.; Azadi, P.; Ley, K.; Dell, A.; Haslam, S.M.; Paulson, J.C. Global metabolic inhibitors of sialyl- and fucosyltransferases remodel the glycome. Nat. Chem. Biol. 2012, 8, 661–668. [Google Scholar] [CrossRef] [Green Version]
- Zandberg, W.F.; Kumarasamy, J.; Pinto, B.M.; Vocadlo, D.J. Metabolic Inhibition of Sialyl-Lewis X Biosynthesis by 5-Thiofucose Remodels the Cell Surface and Impairs Selectin-Mediated Cell Adhesion. J. Biol. Chem. 2012, 287, 40021–40030. [Google Scholar] [CrossRef] [Green Version]
- Kizuka, Y.; Nakano, M.; Yamaguchi, Y.; Nakajima, K.; Oka, R.; Sato, K.; Ren, C.-T.; Hsu, T.-L.; Wong, C.-H.; Taniguchi, N. An Alkynyl-Fucose Halts Hepatoma Cell Migration and Invasion by Inhibiting GDP-Fucose-Synthesizing Enzyme FX, TSTA3. Cell Chem. Biol. 2017, 24, 1467–1478.e5. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Ehret, J.; Kolmar, H.; Zimmer, A. Impact of Acetylated and Non-Acetylated Fucose Analogues on IgG Glycosylation. Antibodies 2019, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.T.; Sewatanon, J.; Memoli, M.J.; Wrammert, J.; Bournazos, S.; Bhaumik, S.K.; Pinsky, B.A.; Chokephaibulkit, K.; Onlamoon, N. IgG antibodies to dengue enhanced for FcgammaRIIIA binding determine disease severity. Science 2017, 355, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gu, J.; Ihara, H.; Miyoshi, E.; Honke, K.; Taniguchi, N. Core Fucosylation Regulates Epidermal Growth Factor Receptor-mediated Intracellular Signaling. J. Biol. Chem. 2006, 281, 2572–2577. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Mao, S.; Sun, S.; Li, M.; Li, Z.; Yu, R.; Ma, T.; Gu, J.; Zhang, J.; Taniguchi, N.; et al. Core Fucosylation of the T Cell Receptor Is Required for T Cell Activation. Front. Immunol. 2018, 9, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.M.; Gaidukov, L.; Jung, G.; Tseng, W.A.; Scarcelli, J.J.; Cornell, R.; Marshall, J.K.; Lyles, J.L.; Sakorafas, P.; Chu, A.-H.A.; et al. Small-molecule control of antibody N-glycosylation in engineered mammalian cells. Nat. Chem. Biol. 2019, 15, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Gao, X.; Cadang, L.; Izadi, S.; Liu, P.; Zhang, H.-M.; Hecht, E.; Shim, J.; Magill, G.; Pabon, J.R.; et al. Fc galactosylation follows consecutive reaction kinetics and enhances immunoglobulin G hexamerization for complement activation. mAbs 2021, 13, 1893427. [Google Scholar] [CrossRef]
- Hodoniczky, J.; Zheng, Y.Z.; James, D.C. Control of Recombinant Monoclonal Antibody Effector Functions by Fc N-Glycan Remodeling in Vitro. Biotechnol. Prog. 2005, 21, 1644–1652. [Google Scholar] [CrossRef]
- van Osch, T.L.J.; Nouta, J.; Derksen, N.I.; van Mierlo, G.; van der Schoot, C.E.; Wuhrer, M.; Rispens, T.; Vidarsson, G. Fc Galactosylation Promotes Hexamerization of Human IgG1, Leading to Enhanced Classical Complement Activation. J. Immunol. 2021, 207, 1545–1554. [Google Scholar] [CrossRef]
- Aoyama, M.; Hashii, N.; Tsukimura, W.; Osumi, K.; Harazono, A.; Tada, M.; Kiyoshi, M.; Matsuda, A.; Ishii-Watabe, A. Effects of terminal galactose residues in mannose α1-6 arm of Fc-glycan on the effector functions of therapeutic monoclonal antibodies. mAbs 2019, 11, 826–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narimatsu, Y.; Joshi, H.; Yang, Z.; Gomes, C.; Chen, Y.-H.; Lorenzetti, F.C.; Furukawa, S.; Schjoldager, K.T.; Hansen, L.; Clausen, H.; et al. A validated gRNA library for CRISPR/Cas9 targeting of the human glycosyltransferase genome. Glycobiology 2018, 28, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Narimatsu, Y.; Clausen, T.M.; Gomes, C.; Karlsson, R.T.G.; Steentoft, C.; Spliid, C.B.; Gustavsson, T.; Salanti, A.; Persson, A.; et al. The GAGOme: A cell-based library of displayed glycosaminoglycans. Nat. Methods 2018, 15, 881–888. [Google Scholar] [CrossRef]
- Narimatsu, Y.; Joshi, H.; Nason, R.; Van Coillie, J.; Karlsson, R.T.G.; Sun, L.; Ye, Z.; Chen, Y.-H.; Schjoldager, K.T.; Steentoft, C.; et al. An Atlas of Human Glycosylation Pathways Enables Display of the Human Glycome by Gene Engineered Cells. Mol. Cell 2019, 75, 394–407.e5. [Google Scholar] [CrossRef]
- Critcher, M.; O’Leary, T.; Huang, M.L. Glycoengineering: Scratching the surface. Biochem. J. 2021, 478, 703–719. [Google Scholar] [CrossRef]
- Shen, J.; Jia, L.; Dang, L.; Su, Y.; Zhang, J.; Xu, Y.; Zhu, B.; Chen, Z.; Wu, J.; Lan, R.; et al. StrucGP: De novo structural sequencing of site-specific N-glycan on glycoproteins using a modularization strategy. Nat. Methods 2021, 18, 921–929. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, X.; D’Antona, A.M.; Scarcelli, J.J.; Rouse, J.C. New Opportunities in Glycan Engineering for Therapeutic Proteins. Antibodies 2022, 11, 5. https://doi.org/10.3390/antib11010005
Zhong X, D’Antona AM, Scarcelli JJ, Rouse JC. New Opportunities in Glycan Engineering for Therapeutic Proteins. Antibodies. 2022; 11(1):5. https://doi.org/10.3390/antib11010005
Chicago/Turabian StyleZhong, Xiaotian, Aaron M. D’Antona, John J. Scarcelli, and Jason C. Rouse. 2022. "New Opportunities in Glycan Engineering for Therapeutic Proteins" Antibodies 11, no. 1: 5. https://doi.org/10.3390/antib11010005
APA StyleZhong, X., D’Antona, A. M., Scarcelli, J. J., & Rouse, J. C. (2022). New Opportunities in Glycan Engineering for Therapeutic Proteins. Antibodies, 11(1), 5. https://doi.org/10.3390/antib11010005