Determination of the Environmental Factors that Affect the Growth and Survival of Greek Fir Seedlings


Abstract
1. Introduction
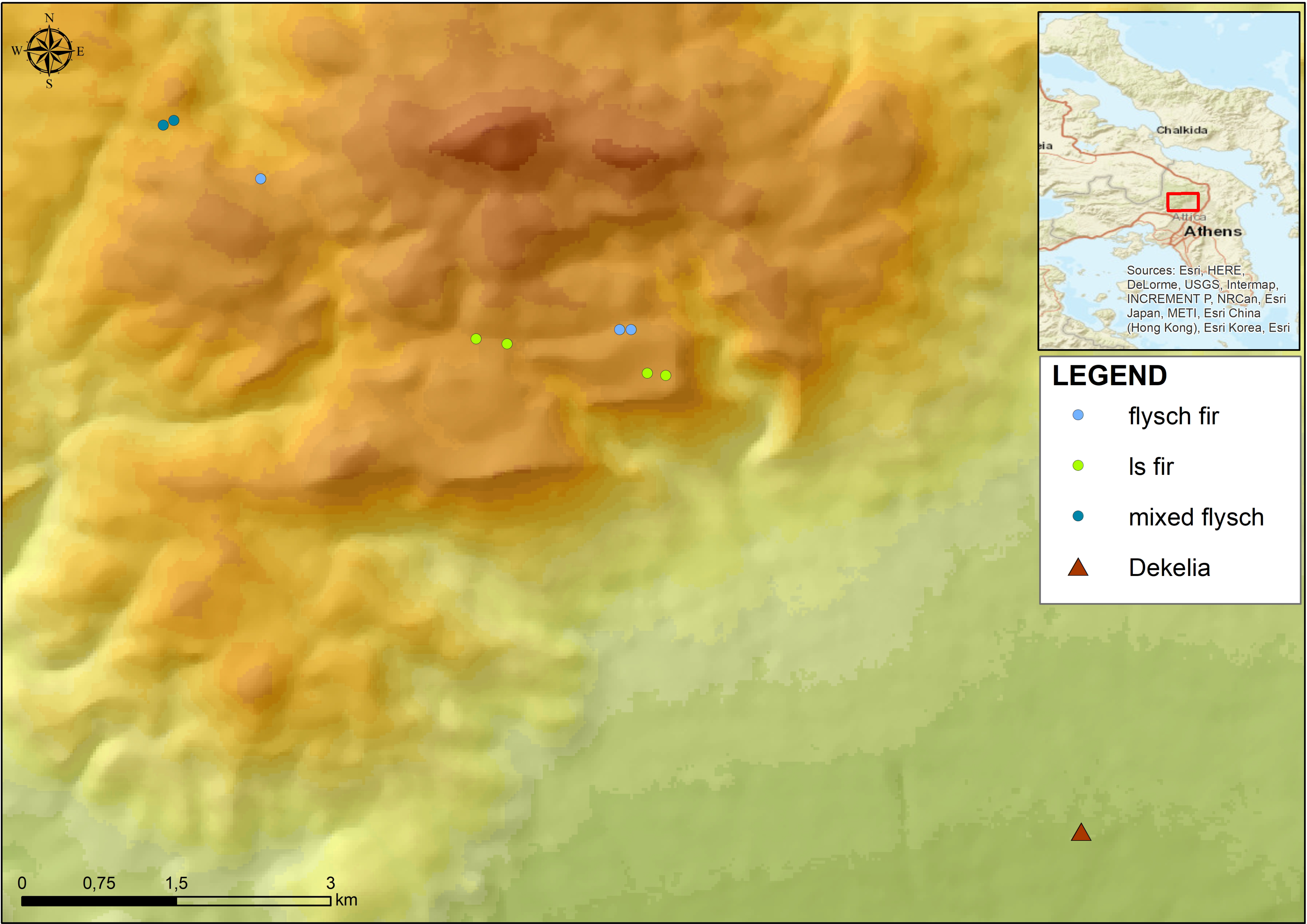
2. Materials and Methods
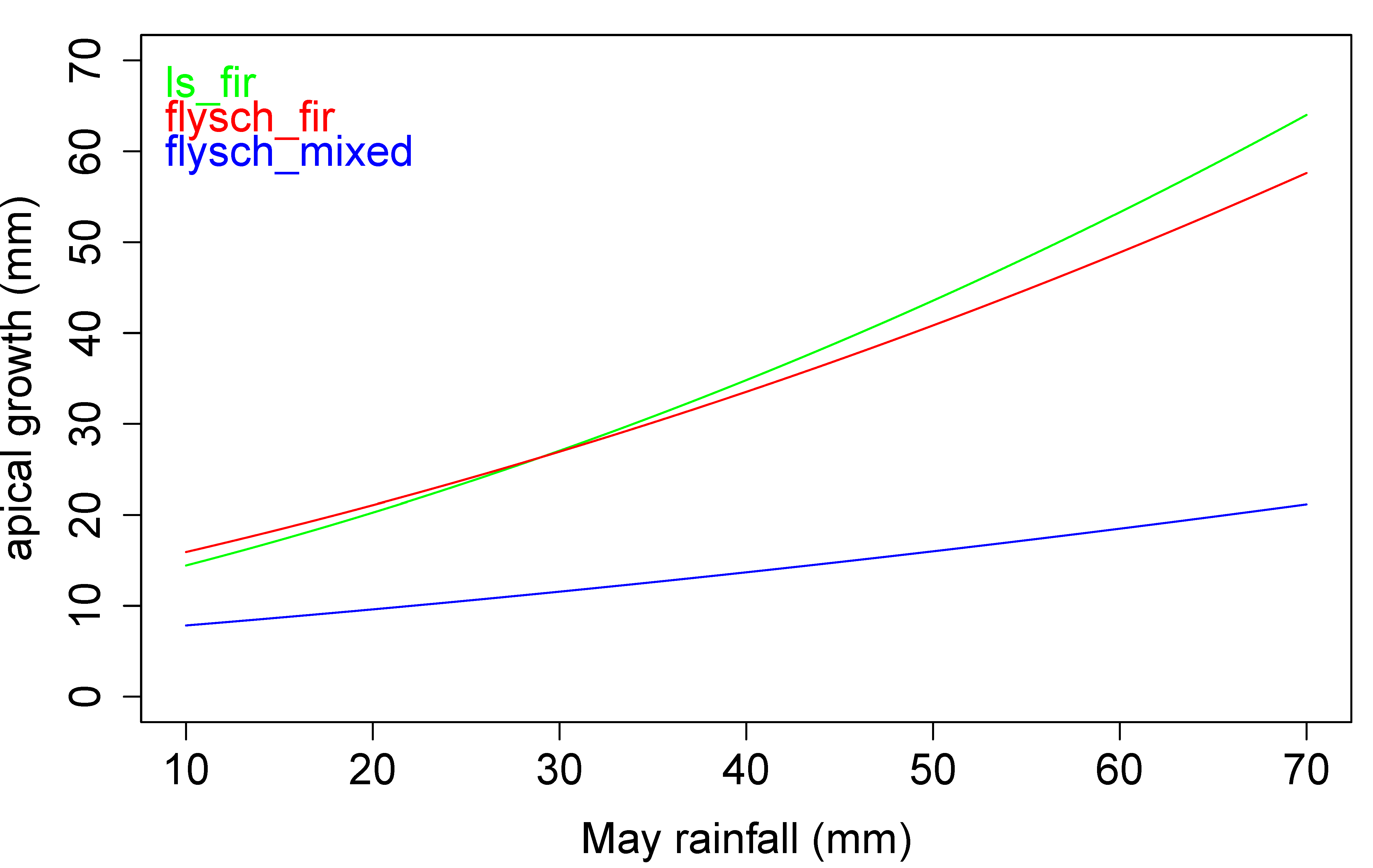
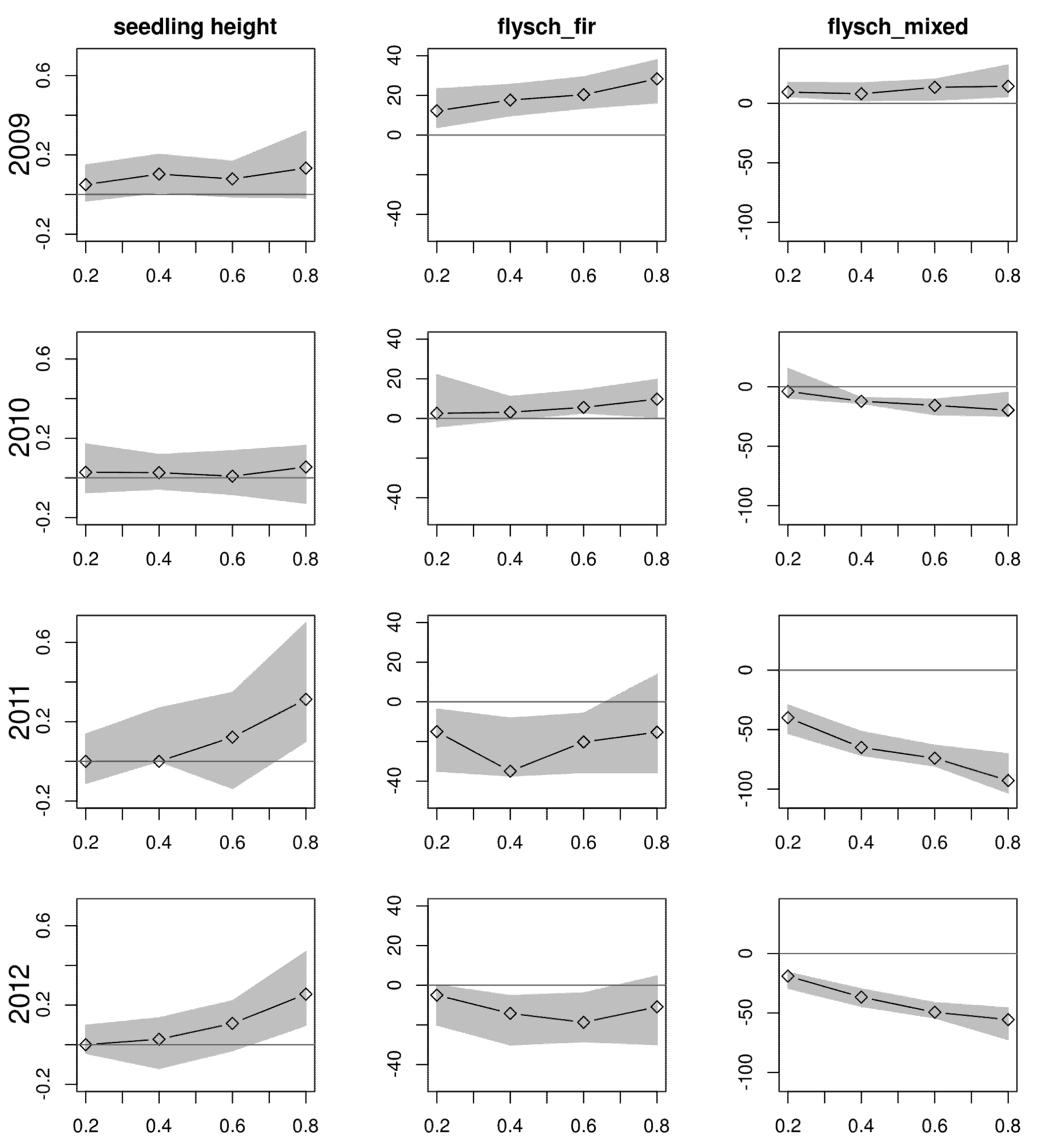
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Dimopoulos, P.; Bergmeier, E.; Eleftheriadou, E.; Theodoropoulos, K.; Gerasimidis, A.; Tsiafouli, M. Identification and Interpretation Guide for the Forest Habitats of Greece; University of Western Greece Publ.: Agrinio, Greece, 2012. [Google Scholar]
- Tsagkari, K.; Karetsos, G.; Proutsos, N. Spatiotemporal investigation on forest fires in Greece. 2011. Available online: http://oikoskopio.gr/pyroskopio/en/ (accessed on 23 March 2020).
- Christopoulou, A.; Kazanis, D.; Fyllas, N.; Arianoutsou, M. Post-fire recovery of Abies cephalonica forest communities: The case of Mt Parnitha National Park, Attica, Greece. iFor.—Biogeosci. For. 2018, 11, 757–764. [Google Scholar] [CrossRef]
- Ganatsas, P.; Daskalakou, E.; Paitaridou, D. First results on early post-fire succession in an Abies cephalonica forest (Parnitha National Park, Greece). iFor.—Biogeosci. For. 2012, 5, 6–12. [Google Scholar] [CrossRef]
- Politi, P.I.; Arianoutsou, M.; Stamou, G.P. Patterns of Abies cephalonica seedling recruitment in Mount Aenos National Park, Cephalonia, Greece. For. Ecol. Manag. 2009, 258, 1129–1136. [Google Scholar] [CrossRef]
- Daskalakou, E.N.; Koutsovoulou, K.; Ioannidis, K.; Koulelis, P.P.; Ganatsas, P.; Thanos, C.A. Masting and regeneration dynamics of Abies cephalonica, the Greek endemic silver fir. Seed Sci. Res. 2019, 29, 227–237. [Google Scholar] [CrossRef]
- Samaras, D.; Gaertner, S.; Reif, A.; Theodoropoulos, K. Drought effects on the floristic differentiation of Greek fir forests in the mountains of central Greece. iFor.—Biogeosci. For. 2015, 8, 786–797. [Google Scholar] [CrossRef]
- Mazza, G.; Gallucci, V.; Manetti, M.C.; Urbinati, C. Tree-Ring Growth Trends of Abies alba Mill: Possible Adaptations to Climate Change in Marginal Populations of Central Italy. Open For. Sci. J. 2013, 46–49. [Google Scholar] [CrossRef][Green Version]
- Pinto, P.E.; Gégout, J.-C.; Hervé, J.-C.; Dhôte, J.-F. Respective importance of ecological conditions and stand composition on Abies alba Mill. dominant height growth. For. Ecol. Manag. 2008, 255, 619–629. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New For. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; del Campo, A.D.; Ceacero, C.J.; Quero, J.L.; Hermoso de Mena, J. On the importance of topography, site quality, stock quality and planting date in a semiarid plantation: Feasibility of using low-density LiDAR. Ecol. Eng. 2014, 67, 25–38. [Google Scholar] [CrossRef]
- Bosela, M.; Popa, I.; Gömöry, D.; Longauer, R.; Tobin, B.; Kyncl, J.; Kyncl, T.; Nechita, C.; Petráš, R.; Sidor, C.G.; et al. Effects of post-glacial phylogeny and genetic diversity on the growth variability and climate sensitivity of European silver fir. J. Ecol. 2016, 104, 716–724. [Google Scholar] [CrossRef]
- Henneb, M.; Thiffault, N.; Valeria, O. Regional Climate, Edaphic Conditions and Establishment Substrates Interact to Influence Initial Growth of Black Spruce and Jack Pine Planted in the Boreal Forest. Forests 2020, 11, 139. [Google Scholar] [CrossRef]
- Moore, J.A.; Hamilton, D.A., Jr.; Xiao, Y.; Byrne, J. Bedrock type significantly affects individual tree mortality for various conifers in the inland Northwest, U.S.A. Can. J. For. Res. 2004, 34, 31–42. [Google Scholar] [CrossRef]
- Odeh, T.; Boulad, N.; Abed, O.; Abu Yahya, A.; Khries, N.; Abu-Jaber, N. The Influence of Geology on Landscape Typology in Jordan: Theoretical Understanding and Planning Implications. Land 2017, 6, 51. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Christopoulou, A.; Galanidis, A.; Michelaki, C.Z.; Dimitrakopoulos, P.G.; Fulé, P.Z.; Arianoutsou, M. Tree growth-climate relationships in a forest-plot network on Mediterranean mountains. Sci. Total Environ. 2017, 598, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Noy-Meir, I. Desert Ecosystems: Environment and Producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Fien, E.K.P.; Fraver, S.; Teets, A.; Weiskittel, A.R.; Hollinger, D.Y. Drivers of individual tree growth and mortality in an uneven-aged, mixed-species conifer forest. For. Ecol. Manag. 2019, 449, 117446. [Google Scholar] [CrossRef]
- Detsis, V.; Efthimiou, G.; Theodoropoulou, O.; Siorokou, S. Reforestation with Abies cephalonica Loudon: A five-year study of seedling survival. Manag. Environ. Qual. An Int. J. 2016, 27, 136–145. [Google Scholar] [CrossRef]
- Reppa, E. Factors Affecting the Efficiency of Erosion Control Measures Following Wildfire. Unpublished Master’s Thesis, Harokopio University, Athens, Greece, 2010. (In Greek). [Google Scholar]
- Amorgianiotis, G. Management Plan of the Parnitha National Park; Forestry of Parnitha: Athens, Greece, 1997. (In Greek) [Google Scholar]
- Raftoyannis, Y.; Radoglou, K.; Bredemeier, M. Effects of mistletoe infestation on the decline and mortality of Abies cephalonica in Greece. Ann. For. Res. 2015, 58, 1. [Google Scholar] [CrossRef]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Siegwolf, R.T.W.; Cotrufo, M.F. Tree rings indicate different drought resistance of a native (Abies alba Mill.) and a nonnative (Picea abies (L.) Karst.) species co-occurring at a dry site in Southern Italy. For. Ecol. Manag. 2009, 257, 820–828. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 23 March 2020).
- Winter, B. Linear models and linear mixed effects models in R with linguistic applications. arXiv 2013, arXiv:1308.5499. Available online: http://arxiv.org/pdf/1308.5499.pdf (accessed on 23 March 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [PubMed]
- Gastwirth, J.L.; Gel, Y.R.; Hui, W.L.W.; Lyubchich, V.; Miao, W.; Noguchi, K. Lawstat: Tools for Biostatistics, Public Policy, and Law. R package version 3.3. 2019. Available online: https://cran.r-project.org/package=lawstat (accessed on 23 March 2020).
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R 2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R package version 1.43.6. 2019. Available online: https://cran.r-project.org/package=MuMIn (accessed on 23 March 2020).
- Cade, B.S.; Noon, B.R. A gentle introduction to quantile regression for ecologists. Front. Ecol. Environ. 2003, 1, 412–420. [Google Scholar] [CrossRef]
- Koenker, R. Quantreg: Quantile Regression. R package version 5.36. 2018. Available online: https://cran.r-project.org/package=quantreg (accessed on 23 March 2020).
- Lele, S.R.; Keim, J.L.; Solymos, P. ResourceSelection: Resource Selection (Probability) Functions for Use-Availability Data. R package version 0.3-5. 2019. Available online: https://cran.r-project.org/package=ResourceSelection (accessed on 23 March 2020).
- Paul, P.; Pennell, M.L.; Lemeshow, S. Standardizing the power of the Hosmer-Lemeshow goodness of fit test in large data sets. Stat. Med. 2013, 32, 67–80. [Google Scholar] [CrossRef]
- Wightman, M.G.; Gonzalez-Benecke, C.A.; Dinger, E.J. Interactive Effects of Stock Type and Forest Vegetation Management Treatments on Douglas-Fir Seedling Growth and Survival—Ten-Year Results. Forests 2019, 10, 1002. [Google Scholar] [CrossRef]
- Charron, L.; Hermanutz, L. Simplicity is key: Restoration protocols for nonregenerating forests degraded by overabundant herbivores. Restor. Ecol. 2017, 25, 432–441. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Puértolas, J.; Cuesta, B.; Peñuelas, J.L.; Uscola, M.; Heredia-Guerrero, N.; Rey Benayas, J.M. Increase in size and nitrogen concentration enhances seedling survival in Mediterranean plantations. Insights from an ecophysiological conceptual model of plant survival. New For. 2012, 43, 755–770. [Google Scholar] [CrossRef]
- Reisman-Berman, O. Age-related change in canopy traits shifts conspecific facilitation to interference in a semi-arid shrubland. Ecography (Cop.) 2007, 30, 459–470. [Google Scholar] [CrossRef]
- Nakos, G. Forest soils of Greece: Physical, chemical and biological properties. For. Ecol. Manag. 1979, 2, 35–51. [Google Scholar] [CrossRef]
- Koutavas, A. CO2 fertilization and enhanced drought resistance in Greek firs from Cephalonia Island, Greece. Glob. Chang. Biol. 2013, 19, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, A. Tree-ring patterns and climate response of Mediterranean fir populations in Central Greece. Dendrochronologia 2016, 40, 17–25. [Google Scholar] [CrossRef]
- Torres, J.; Marques, J.; Alves, P.; Costa, H.; Honrado, J. Local lithological drivers of post-fire vegetation recovery and implications for fire-prone regions. Ecol. Res. 2017, 32, 37–49. [Google Scholar] [CrossRef]
- Marcolin, E.; Marzano, R.; Vitali, A.; Garbarino, M.; Lingua, E. Post-Fire Management Impact on Natural Forest Regeneration through Altered Microsite Conditions. Forests 2019, 10, 1014. [Google Scholar] [CrossRef]
- Vickers, L.; Houser, J.; Rooni, J.; Guldin, J. Some Lessons Learned on Early Survival and Growth of Containerized, Locally-Sourced Ponderosa Pine Seedlings in the Davis Mountains of Western Texas, US. Forests 2019, 10, 267. [Google Scholar] [CrossRef]
- Chang, K.-H.; Price, D.T.; Chen, J.M.; Kurz, W.A.; Boisvenue, C.; Hogg, E.H.; Black, T.A.; Gonsamo, A.; Wu, C.; Hember, R.A. Simulating impacts of water stress on woody biomass in the southern boreal region of western Canada using a dynamic vegetation model. Agric. For. Meteorol. 2014, 198–199, 142–154. [Google Scholar] [CrossRef]
- Fensham, R.J.; Butler, D.W.; Foley, J. How does clay constrain woody biomass in drylands? Glob. Ecol. Biogeogr. 2015, 24, 950–958. [Google Scholar] [CrossRef]
- Fernandez-Illescas, C.P.; Porporato, A.; Laio, F.; Rodriguez-Iturbe, I. The ecohydrological role of soil texture in a water-limited ecosystem. Water Resour. Res. 2001, 37, 2863–2872. [Google Scholar] [CrossRef]
- Renne, R.R.; Bradford, J.B.; Burke, I.C.; Lauenroth, W.K. Soil texture and precipitation seasonality influence plant community structure in North American temperate shrub steppe. Ecology 2019, 100, e02824. [Google Scholar] [CrossRef]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary Production of the Central Grassland Region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Yang, Y.H.; Fang, J.Y.; Pan, Y.D.; Ji, C.J. Aboveground biomass in Tibetan grasslands. J. Arid Environ. 2009, 73, 91–95. [Google Scholar] [CrossRef]
- Weng, E.; Luo, Y. Soil hydrological properties regulate grassland ecosystem responses to multifactor global change: A modeling analysis. J. Geophys. Res. 2008, 113, G03003. [Google Scholar] [CrossRef]
- Fady, B. Biologie et forêt, caractéristiques écologiques et sylvicoles des sapins de Grèce dans leur aire naturelle et en plantation dans le sud de la France. Rev. For. Française 1993, 119–133. [Google Scholar] [CrossRef]
- Daskalakou, E.N.; Koutsovoulou, K.; Mavroeidi, L.; Tsiamitas, C.; Kafali, E.; Radaiou, P.-E.; Ganatsas, P.; Thanos, C.A. Interannual variability of germination and cone/seed morphometric characteristics in the endemic Grecian fir (Abies cephalonica) over an 8-year-long study. Seed Sci. Res. 2018, 28, 24–33. [Google Scholar] [CrossRef]
- Papageorgiou, A.C.; Kostoudi, C.; Sorotos, I.; Varsamis, G.; Korakis, G.; Drouzas, A.D. Diversity in needle morphology and genetic markers in a marginal Abies cephalonica (Pinaceae) population. Ann. For. Res. 2015, 58, 217–234. [Google Scholar] [CrossRef]
- Valdecantos, A.; Fuentes, D.; Smanis, A.; Llovet, J.; Morcillo, L.; Bautista, S. Effectiveness of Low-Cost Planting Techniques for Improving Water Availability to Olea europaea Seedlings in Degraded Drylands. Restor. Ecol. 2014, 22, 327–335. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | 05 | 06 | 07 | 08 | 09 | 10 |
|---|---|---|---|---|---|---|
| 2009 | 44.1 | 0 | 71.4 | 0 | 60.1 | 92.8 |
| 2010 | 16.6 | 21.5 | 30.5 | 0 | 15.8 | 92.4 |
| 2011 | 61.4 | 21.0 | 0 | 0 | 3.5 | 32.1 |
| 2012 | 23.9 | 0 | 0 | 1.9 | 3.6 | 39.7 |
| Year | Site type | mean | median | sd |
|---|---|---|---|---|
| 2009 | ls_fir | 52.43 | 49.5 | 23.44 |
| flysch_fir | 64.14 | 64.5 | 27.84 | |
| flysch_mixed | 50.41 | 48.5 | 25.2 | |
| 2010 | ls_fir | 40.18 | 40 | 16.5 |
| flysch_fir | 41.41 | 40 | 22.9 | |
| flysch_mixed | 26.57 | 21 | 15.34 | |
| 2011 | ls_fir | 111.82 | 110 | 46.18 |
| flysch_fir | 82.12 | 70 | 43.88 | |
| flysch_mixed | 37.55 | 35 | 16.08 | |
| 2012 | ls_fir | 69.89 | 65 | 38.92 |
| flysch_fir | 53.59 | 45 | 33.9 | |
| flysch_mixed | 23.23 | 20 | 10.99 |
| Fixed effects | ||||||
| estimate | sd | 2.50% | 97.50% | |||
| intercept | 3.00 | 0.47 | 1.93 | 3.96 | ||
| slope: initial height | 0.006 | 0.002 | 0.001 | 0.010 | ||
| slope: age | 0.30 | 0.05 | 0.20 | 0.40 | ||
| Random effects of rainfall by site type | ||||||
| ls_fir | flysch_fir | flysch_mixed | ||||
| intercept | 3.1 | 3.39 | 2.5 | |||
| slope | 0.07 | 0.06 | 0.03 | |||
| Random effects: standard deviation | ||||||
| sd | 2.50% | 97.50% | ||||
| Intercept: subject | 0.83 | 0.65 | 1.01 | |||
| slope: rainfall|site type | 0.06 | 0.03 | 0.17 | |||
| Intercept: site type | 0.43 | 0.00 | 1.52 | |||
| sigma | 1.90 | 1.81 | 2.00 | |||
| Year | Site Type | n | Mean | Median | sd | Survival |
|---|---|---|---|---|---|---|
| 2009 | ls_fir | 192 | 45.66 | 45 | 24.48 | 73.55 |
| flysch_fir | 147 | 62.29 | 60 | 34.26 | 81.82 | |
| flysch_mixed | 103 | 56.51 | 52 | 28.94 | 69.77 | |
| 2010 | ls_fir | 189 | 34.39 | 35 | 21.9 | 89.33 |
| flysch_fir | 136 | 41.9 | 40 | 23.21 | 94.77 | |
| flysch_mixed | 71 | 24.61 | 21 | 14.32 | 79.78 | |
| 2011 | ls_fir | 122 | 107.66 | 100 | 46.91 | 74.69 |
| flysch_fir | 122 | 85.94 | 72.5 | 47.27 | 89.73 | |
| flysch_mixed | 62 | 36.94 | 35 | 15.48 | 90.14 |
| Coefficients | 2.50% | 97.50% | |
|---|---|---|---|
| 2009 | |||
| (Intercept) | 32.00 | 18.79 | 41.58 |
| initial height (mm) | 0.10 | 0.04 | 0.17 |
| site flysch_fir | 19.90 | 13.55 | 25.64 |
| site flysch_mixed | 9.00 | 3.25 | 17.40 |
| 2010 | |||
| (Intercept) | 28.00 | 20.64 | 37.92 |
| initial height (mm) | 0.05 | −0.03 | 0.10 |
| site flysch_fir | 7.50 | −1.21 | 11.82 |
| site flysch_mixed | −13.50 | −17.72 | −9.74 |
| 2011 | |||
| (Intercept) | 88.13 | 71.61 | 111.29 |
| initial height (mm) | 0.13 | −0.06 | 0.24 |
| site flysch_fir | −24.13 | −51.99 | −13.80 |
| site flysch_mixed | −68.13 | −73.49 | −60.48 |
| 2012 | |||
| (Intercept) | 64.17 | 52.06 | 73.45 |
| initial height (mm) | 0.01 | −0.07 | 0.11 |
| site flysch_fir | −20.08 | −27.66 | −12.83 |
| site flysch_mixed | −44.79 | −50.79 | −41.12 |
| Estimate | sd | |
|---|---|---|
| 2009, H-L test p = 0.43 | ||
| intercept | −0.962 * | 0.423 |
| initial height | 0.017 *** | 0.003 |
| flysch_fir | 1.074 *** | 0.278 |
| flysch_mixed | 0.007 | 0.252 |
| 2010, H-L test p = 0.78 | ||
| intercept | 0.809 | 0.660 |
| initial height | 0.011 * | 0.005 |
| flysch_fir | 1.171 * | 0.479 |
| flysch_mixed | −0.628 | 0.364 |
| 2011, H-L test p = 0.22 | ||
| intercept | −1.954 ** | 0.642 |
| initial height | 0.026 *** | 0.005 |
| flysch_fir | 2.035 *** | 0.408 |
| flysch_mixed | 1.358 ** | 0.460 |
| 2012, H-L test p = 0.87 | ||
| intercept | 1.713 * | 0.739 |
| initial height | 0.002 | 0.005 |
| flysch_fir | 0.440 | 0.458 |
| flysch_mixed | −0.572 | 0.420 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Detsis, V.; Efthimiou, G.; Theodoropoulou, O.; Siorokou, S. Determination of the Environmental Factors that Affect the Growth and Survival of Greek Fir Seedlings. Land 2020, 9, 100. https://doi.org/10.3390/land9040100
Detsis V, Efthimiou G, Theodoropoulou O, Siorokou S. Determination of the Environmental Factors that Affect the Growth and Survival of Greek Fir Seedlings. Land. 2020; 9(4):100. https://doi.org/10.3390/land9040100
Chicago/Turabian StyleDetsis, Vassilis, Georgios Efthimiou, Olga Theodoropoulou, and Stavroula Siorokou. 2020. "Determination of the Environmental Factors that Affect the Growth and Survival of Greek Fir Seedlings" Land 9, no. 4: 100. https://doi.org/10.3390/land9040100
APA StyleDetsis, V., Efthimiou, G., Theodoropoulou, O., & Siorokou, S. (2020). Determination of the Environmental Factors that Affect the Growth and Survival of Greek Fir Seedlings. Land, 9(4), 100. https://doi.org/10.3390/land9040100