From the Ground Up: Prairies on Reclaimed Mine Land—Impacts on Soil and Vegetation


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
- (a)
- Grassland types: Prairie, Baseline
- (b)
- Prairie age (in 2018): A (14–19 years), B (8–10 years), C (4 years), D (2 years)
- (c)
- Geographic location: C (Central), E (East), N (North), S (South)
- (d)
- Permit type: B (1972–1977), C (1977–1982), U (Unmined)
- (e)
- Native plant cover percentage: A (0–25%), B (25–50%), C (>50%)
2.2. Plant Sampling
2.3. Soil Sampling
2.3.1. Soil Chemistry
2.3.2. Soil Microbial Analyses
- Actinobacteria = Sum of 16:0 10-methyl, 17:0 10-methyl, and 18:0 10-methyl
- General Bacteria = Sum of 14:0 iso, 15:0, and 17:0
- Gram- Bacteria = Sum of 16:1 ω7c, 17:0 cyclo, 19:0 cyclo ω8c, and 18:1 ω7c
- Gram+ Bacteria = Sum of 15:0 iso, 15:0 anteiso, 16:0 iso, 17:0 iso and 17:0 anteiso
- Fungi = Sum of 18:2 ω6c and 18:1 ω9c
- Arbuscular mycorrhiza fungi (AMF) = 16:1 ω5c
- Protozoa = Sum of 20:2 ω6c, 20:3 ω6c and 20:4 ω6c
- Total FAME concentration (sum of all above) served as a marker for soil microbial biomass
2.4. Data Transformations and Statistical Analyses
2.4.1. Microbial Data Transformations
2.4.2. Data Analyses
2.4.3. Significance Testing
3. Results
3.1. Plants
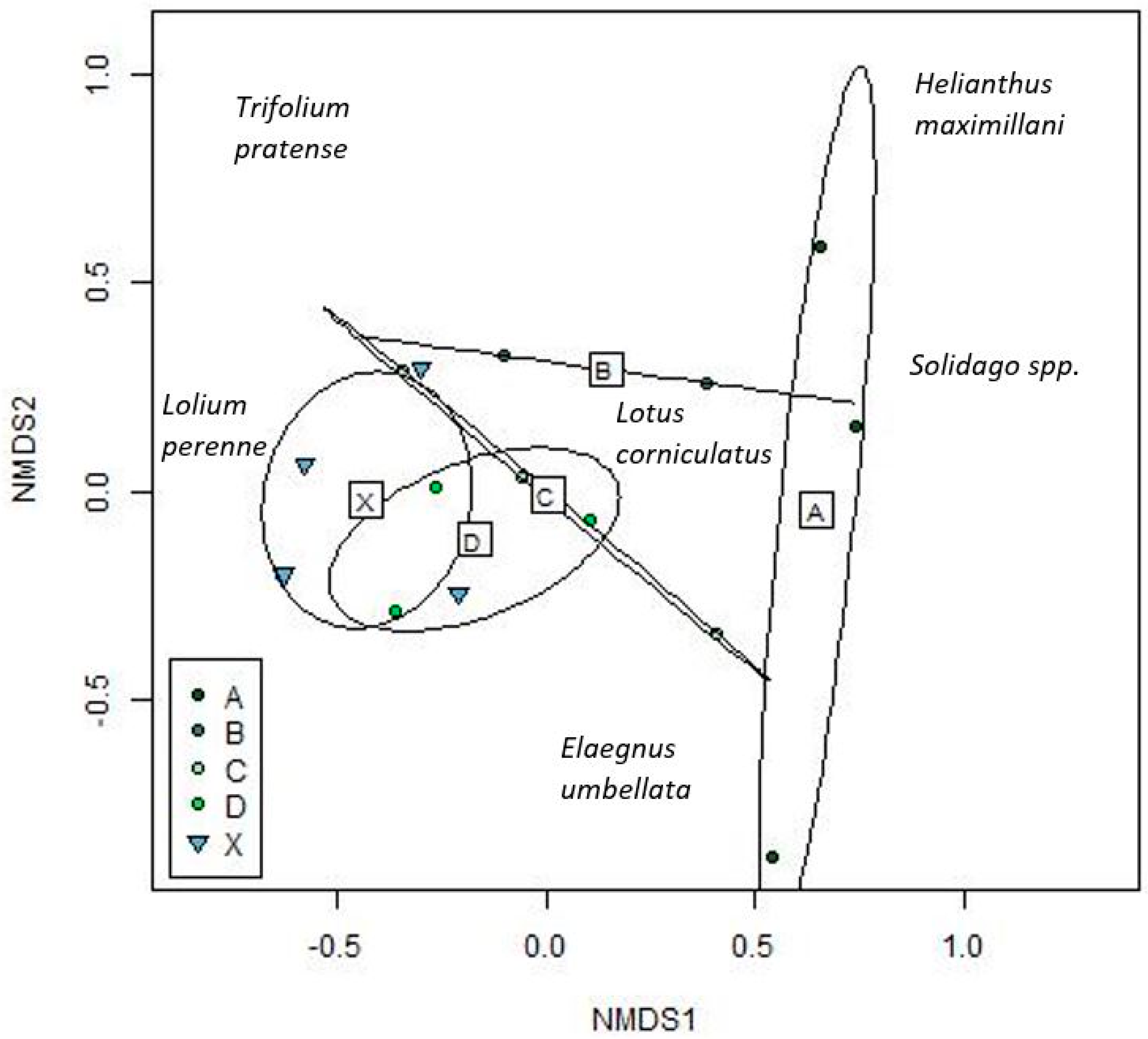
3.1.1. Plant Community Composition
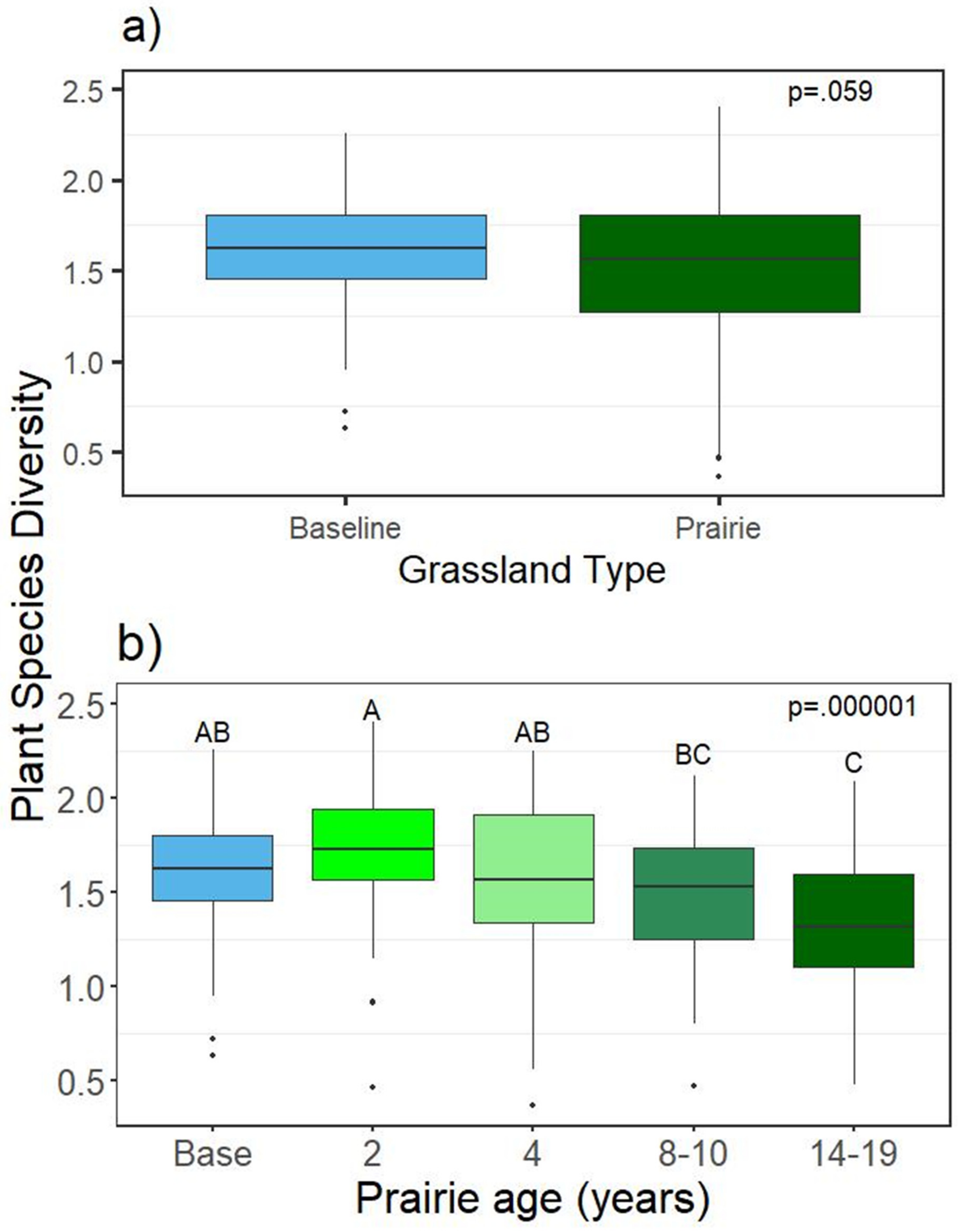
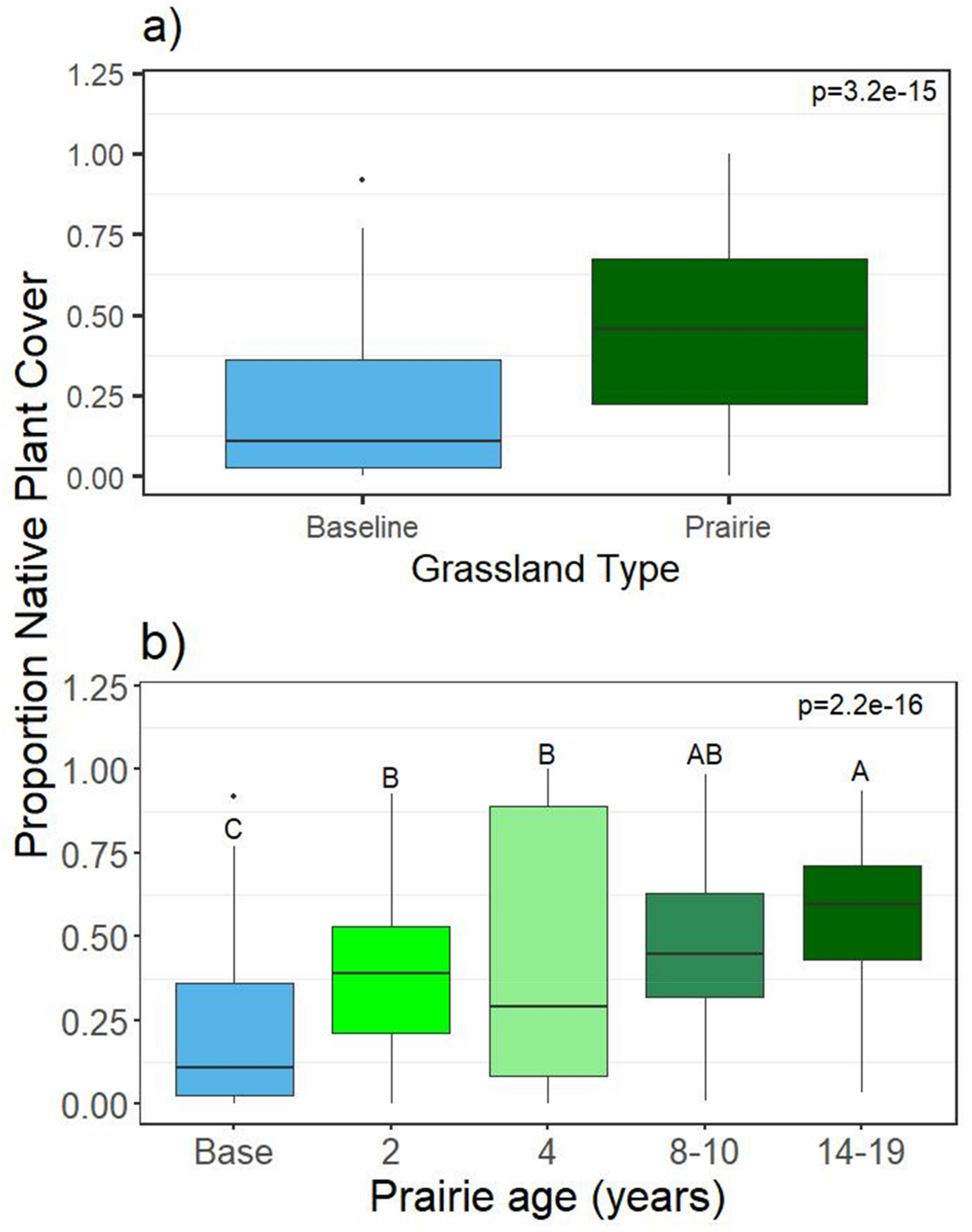
3.1.2. Plant Richness and Diversity
3.2. Soil
3.2.1. Soil Chemistry
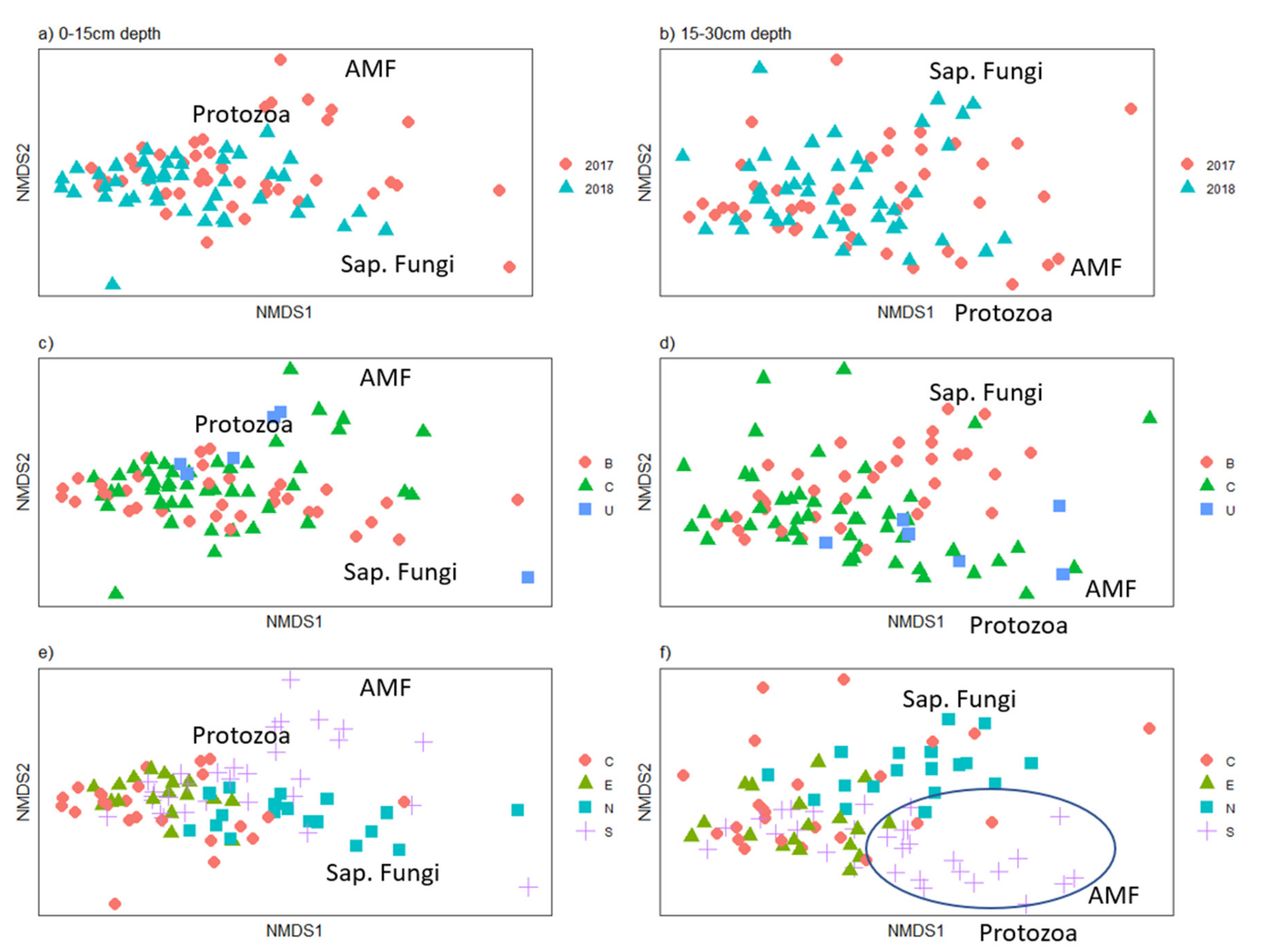
3.2.2. Soil Microbes
4. Discussion
4.1. Plants: Richness, Diversity, and Cover
4.2. Soil Biochemical and Microbial Properties after Prairie Establishment
4.3. Soil Chemical Properties after Prairie Establishment
4.4. Prairie Soil Development over Time
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McAlpine, C.A.; Catterall, C.P.; Mac Nally, R.; Lindenmayer, D.; Reid, J.L.; Holl, K.D.; Bennett, A.F.; Runting, R.K.; Wilson, K.; Hobbs, R.J.; et al. Integrating plant- and animal-based perspectives for more effective restoration of biodiversity. Front. Ecol. Environ. 2016, 14, 37–45. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Harris, J.A. Restoration Ecology: Repairing the Earth’s Ecosystems in the New Millennium. Restor. Ecol. 2001, 9, 239–246. [Google Scholar] [CrossRef]
- Martin, L.M.; Moloney, K.A.; Wilsey, B.J. An assessment of grassland restoration success using species diversity components. J. Appl. Ecol. 2005, 42, 327–336. [Google Scholar] [CrossRef]
- Taft, J.B.; Hauser, C.; Robertson, K.R. Estimating floristic integrity in tallgrass prairie. Biol. Conserv. 2006, 131, 42–51. [Google Scholar] [CrossRef]
- Henderson, R. Response to Henry Howe. Ecol. Restor. N. Am. 1999, 17, 189–192. [Google Scholar] [CrossRef]
- Sluis, W.J. Patterns of Species Richness and Composition in Re-Created Grassland. Restor. Ecol. 2002, 10, 677–684. [Google Scholar] [CrossRef]
- Brye, K.R.; Norman, J.M.; Gower, S.T. Assessing the Progress of a Tallgrass Prairie Restoration in Southern Wisconsin. Am. Midl. Nat. 2002, 148, 218. [Google Scholar] [CrossRef]
- Zipper, C.E.; Burger, J.A.; Skousen, J.G.; Angel, P.N.; Barton, C.D.; Davis, V.; Franklin, J.A. Restoring Forests and Associated Ecosystem Services on Appalachian Coal Surface Mines. Environ. Manag. 2011, 47, 751–765. [Google Scholar] [CrossRef]
- Holl, K.D.; Cairns, J. Vegetational Community Development on Reclaimed Coal Surface Mines in Virginia. Bull. Torrey Bot. Club 1994, 121, 327. [Google Scholar] [CrossRef]
- Cavender, N.; Byrd, S.; Bechtoldt, C.L.; Bauman, J.M. Vegetation Communities of a Coal Reclamation Site in Southeastern Ohio. Northeast. Nat. 2014, 21, 31–46. [Google Scholar] [CrossRef]
- Skousen, J.; Gorman, J.; Pena-Yewtukhiw, E.; King, J.; Stewart, J.; Emerson, P.; Delong, C. Hardwood Tree Survival in Heavy Ground Cover on Reclaimed Land in West Virginia: Mowing and Ripping Effects. J. Environ. Qual. 2009, 38, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Cusser, S.; Goodell, K. Diversity and Distribution of Floral Resources Influence the Restoration of Plant-Pollinator Networks on a Reclaimed Strip Mine. Restor. Ecol. 2013, 21, 713–721. [Google Scholar] [CrossRef]
- Drake, L.D. Erosion control with prairie grasses in Iowa strip-mine reclamation. In Proceedings of the 7th North American Prairie Conference, Springfield, MO, USA, 4–6 August 1980; pp. 4–6. [Google Scholar]
- Rodgers, C.; Anderson, R. Establishment of Grasses on Sewage Sludge-Amended Strip Mine Spoils. In Proceedings of the North American Prairie Conferences, Lincoln, NE, USA, 7–11 August 1988. [Google Scholar]
- Schramm, P.; Kalvin, R.L. The use of prairie in strip mine reclamation. In Proceedings of the Fifth Midwest Prairie Conference; Glenn-Lewin, D.C., Landers, R.Q., Jr., Eds.; Iowa State University: Ames, IA, USA, 1978; pp. 151–153. [Google Scholar]
- Heckman, J.R. Restoration of Degraded Land: A Comparison of Structural and Functional Measurements of Recovery. Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 1997. [Google Scholar]
- Missouri Department of Natural Resources. Warm-Season, Native Grasses on Reclaimed Minelands-Landowner Management Guide. 2014. Available online: http://www.dnr.mo.gov/pubs/pub2082.htm (accessed on 5 November 2016).
- Swab, R.; Lorenz, N.; Byrd, S.; Dick, R. Native vegetation in reclamation: Improving habitat and ecosystem function through using prairie species in mine land reclamation. Ecol. Eng. 2017, 108, 525–536. [Google Scholar] [CrossRef]
- Lannoo, M.; Kinney, V.C.; Heemeyer, J.L.; Engbrecht, N.J.; Gallant, A.L.; Klaver, R.W. Mine Spoil Prairies Expand Critical Habitat for Endangered and Threatened Amphibian and Reptile Species. Diversity 2009, 1, 118–132. [Google Scholar] [CrossRef]
- Camill, P.; McKone, M.J.; Sturges, S.T.; Severud, W.J.; Ellis, E.; Limmer, J.; Martin, C.B.; Navratil, R.T.; Purdie, A.J.; Sandel, B.S.; et al. Community—And Ecosystem-Level Changes in a Species-Rich Tallgrass Prairie Restoration. Ecol. Appl. 2004, 14, 1680–1694. [Google Scholar] [CrossRef]
- McLachlan, S.; Knispel, A. Assessment of long-term tallgrass prairie restoration in Manitoba, Canada. Biol. Conserv. 2005, 124, 75–88. [Google Scholar] [CrossRef]
- Cottam, G.; Wilson, H.C. Community Dynamics on an Artificial Prairie. Ecology 1966, 47, 88–96. [Google Scholar] [CrossRef]
- Baer, S.G.; Kitchen, D.J.; Blair, J.M.; Rice, C.W. Changes in ecosystem structure and function along a chronosequence of restored grasslands. Ecol. Appl. 2002, 12, 1688–1701. [Google Scholar] [CrossRef]
- Hansen, M.J.; Gibson, D.J. Use of multiple criteria in an ecological assessment of a prairie restoration chronosequence. Appl. Veg. Sci. 2014, 17, 63–73. [Google Scholar] [CrossRef]
- Allison, V.J.; Miller, R.M.; Jastrow, J.; Matamala, R.; Zak, D.R. Changes in Soil Microbial Community Structure in a Tallgrass Prairie Chronosequence. Soil Sci. Soc. Am. J. 2005, 69, 1412–1421. [Google Scholar] [CrossRef]
- Matamala, R.; Jastrow, J.D.; Miller, R.M.; Garten, C.T. Temporal Changes in C and N Stocks of Restored Prairie: Implications for C Sequestration Strategies. Ecol. Appl. 2008, 18, 1470–1488. [Google Scholar] [CrossRef] [PubMed]
- Barber, N.A.; Jones, H.P.; Duvall, M.R.; Wysocki, W.P.; Hansen, M.J.; Gibson, D.J. Phylogenetic diversity is maintained despite richness losses over time in restored tallgrass prairie plant communities. J. Appl. Ecol. 2017, 54, 137–144. [Google Scholar] [CrossRef]
- Calderón, K.; Spor, A.; Breuil, M.-C.; Bru, D.; Bizouard, F.; Violle, C.; Barnard, R.L.; Philippot, L. Effectiveness of ecological rescue for altered soil microbial communities and functions. ISME J. 2017, 11, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, L.; Miller, S.P.; Baer, S.; Callaham, M.A.; Montgomery, J.; Pavao-Zuckerman, M.; Rhoades, C.C.; Richardson, S. Integrating Soil Ecological Knowledge into Restoration Management. Restor. Ecol. 2008, 16, 608–617. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Lal, R. Changes in physical and chemical properties of soil after surface mining and reclamation. Geoderma 2011, 161, 168–176. [Google Scholar] [CrossRef]
- Ussiri, D.A.; Lal, R. Method for Determining Coal Carbon in the Reclaimed Minesoils Contaminated with Coal. Soil Sci. Soc. Am. J. 2008, 72, 231–237. [Google Scholar] [CrossRef]
- Corbett, E.; Anderson, R.C.; Rodgers, C.S. Prairie Revegetation of a Strip Mine in Illinois: Fifteen Years after Establishment. Restor. Ecol. 1996, 4, 346–354. [Google Scholar] [CrossRef]
- Norland, M.R. Soil Factors Affecting Mycorrhizal Use in Surface Mine Reclamation; Information Circular/1993; US Department of the Interior, Bureau of Mines: Minneapolis, MN, USA, 1993. [Google Scholar]
- Ngugi, M.R.; Neldner, V.J.; Doley, D.; Kusy, B.; Moore, D.; Richter, C. Soil moisture dynamics and restoration of self-sustaining native vegetation ecosystem on an open-cut coal mine. Restor. Ecol. 2015, 23, 615–624. [Google Scholar] [CrossRef]
- Fornara, D.; Tilman, D. Ecological mechanisms associated with the positive diversity–productivity relationship in an N-limited grassland. Ecology 2009, 90, 408–418. [Google Scholar] [CrossRef]
- Tilman, D.; Hill, J.; Lehman, C. Carbon-Negative Biofuels from Low-Input High-Diversity Grassland Biomass. Science 2006, 314, 1598–1600. [Google Scholar] [CrossRef]
- Samson, F.; Knopf, F. Prairie Conservation in North America. Bioscience 1994, 44, 418–421. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Knapp, A.K. Consequences of Nonequilibrium Resource Availability Across Multiple Time Scales: The Transient Maxima Hypothesis. Am. Nat. 1993, 141, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Braun, E.L. The Deciduous Forests of Eastern North America; MacMillan: New York, NY, USA, 1950. [Google Scholar]
- Soil Survey Staff. Natural Resources Conservation Service. United States Department of Agriculture. Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/ (accessed on 12 January 2019).
- Daubenmire, R. Canopy coverage method of vegetation analysis. Northwest Sci. 1959, 33, 39–64. [Google Scholar]
- United States Department of Agriculture Natural Resources Conservation Service (USDA NRCS). The PLANTS Database. National Plant Data Team, Greensboro, NC, USA. 2011. Available online: http://plants.usda.gov (accessed on 28 July 2017).
- Schwartz, M.W. Defining indigenous species: An introduction. In Assessment and Management of Plant Invasions; Springer: New York, NY, USA, 1997; pp. 7–17. [Google Scholar]
- Transeau, E.N. The Prairie Peninsula. Ecology 1935, 16, 423–437. [Google Scholar] [CrossRef]
- Cooperrider, T.S.; Cusick, A.; Kartesz, J.T. Seventh Catalog of the Vascular Plants of Ohio; Ohio State University Press: Columbus, OH, USA, 2001. [Google Scholar]
- Skousen, J. Methods to determine vegetation production on surface mines. Int. J. Surf. Mining, Reclam. Environ. 1990, 4, 173–179. [Google Scholar] [CrossRef]
- Culman, S.W.; Snapp, S.S.; Freeman, M.A.; Schipanski, M.E.; Beniston, J.; Lal, R.; Drinkwater, L.E.; Franzluebbers, A.J.; Glover, J.D.; Grandy, A.S.; et al. Permanganate Oxidizable Carbon Reflects a Processed Soil Fraction that is Sensitive to Management. Soil Sci. Soc. Am. J. 2012, 76, 494–504. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil Enzymes. In Methods of Soil Analysis Part 2 Microbiological and Biochemical Properties; Weaver, R.W., Angle, J.S., Bottomley, P.S., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Schutter, M.E.; Dick, R.P. Comparison of Fatty Acid Methyl Ester (FAME) Methods for Characterizing Microbial Communities. Soil Sci. Soc. Am. J. 2000, 64, 1659–1668. [Google Scholar] [CrossRef]
- Vallejo, V.E.; Arbeli, Z.; Terán, W.; Lorenz, N.; Dick, R.P.; Roldan, F. Effect of land management and Prosopis juliflora (Sw.) DC trees on soil microbial community and enzymatic activities in intensive silvopastoral systems of Colombia. Agric. Ecosyst. Environ. 2012, 150, 139–148. [Google Scholar] [CrossRef]
- Groffman, P.M.; Eagan, P.; Sullivan, W.M.; Lemunyon, J.L. Grass species and soil type effects on microbial biomass and activity. Plant Soil 1996, 183, 61–67. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impacts of Carbon and Flooding on Soil Microbial Communities: Phospholipid Fatty Acid Profiles and Substrate Utilization Patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Kennedy, A.C. Phospholipid fatty acid profiles and carbon utilization patterns for analysis of microbial community structure under field and greenhouse conditions. Microbiol. Ecol. 1998, 26, 151–163. [Google Scholar] [CrossRef]
- Schutter, M.; Sandeno, J.; Dick, R.P. Seasonal, soil type, and alternative management influences on microbial communities of vegetable cropping systems. Biol. Fertil. Soils 2001, 34, 397–410. [Google Scholar] [CrossRef]
- Zobeck, T.M.; Gill, T.E.; Kennedy, A.C. Enzyme activities and microbial community structure in semiarid agricultural soils. Biol. Fertil. Soils 2003, 38, 216–227. [Google Scholar] [CrossRef]
- Lee, Y.B.; Lorenz, N.; Dick, L.K.; Dick, R.P. Cold Storage and Pretreatment Incubation Effects on Soil Microbial Properties. Soil Sci. Soc. Am. J. 2007, 71, 1299–1305. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Version 4.0.0 Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 11 July 2020).
- Wickham, B. GGplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wright, K. Corrgram: Plot a Correlogram. R Package Version 1.13. 2018. Available online: https://CRAN.R-project.org/package=corrgram (accessed on 6 November 2020).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package; R package Version 2.5-5; 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 17 November 2020).
- Pohlert, T. PMCMRplus: Calculate Pairwise Multiple Comparisons of Mean Rank Sums Extended; R package version 1.7.0. Available online: https://cran.r-project.org/web/packages/PMCMRplus/ (accessed on 17 November 2020).
- Howe, H.F. Dominance, Diversity and Grazing in Tallgrass Restoration: Ecology has much to contribute to debates over the role of grazing in restoration—And much to learn from the results of experiments in restorative grazing. Ecol. Restor. N. Am. 1999, 17, 59–66. [Google Scholar] [CrossRef]
- Williams, D.W.; Jackson, L.L.; Smith, D.D. Effects of Frequent Mowing on Survival and Persistence of Forbs Seeded into a Species-Poor Grassland. Restor. Ecol. 2007, 15, 24–33. [Google Scholar] [CrossRef]
- Piper, J.K.; Schmidt, E.S.; Janzen, A.J. Effects of Species Richness on Resident and Target Species Components in a Prairie Restoration. Restor. Ecol. 2007, 15, 189–198. [Google Scholar] [CrossRef]
- Foster, B.L. Establishment, competition and the distribution of native grasses among Michigan old-fields. J. Ecol. 1999, 87, 476–489. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Bardgett, R.D.; Vitousek, P.M.; Maestre, F.T.; Williams, M.A.; Eldridge, D.J.; Lambers, H.; Eneuhauser, S.; Gallardo, A.; García-Velázquez, L.; et al. Changes in belowground biodiversity during ecosystem development. Proc. Natl. Acad. Sci. USA 2019, 116, 6891–6896. [Google Scholar] [CrossRef]
- Lorenz, K.; Lal, R. Stabilization of organic carbon in chemically separated pools in reclaimed coal mine soils in Ohio. Geoderma 2007, 141, 294–301. [Google Scholar] [CrossRef]
- Rumpel, C.; Balesdent, J.; Grootes, P.; Weber, E.; Kögel-Knabner, I. Quantification of lignite- and vegetation-derived soil carbon using 14C activity measurements in a forested chronosequence. Geoderma 2003, 112, 155–166. [Google Scholar] [CrossRef]
- Knops, J.M.; Tilman, D. Dynamics of soil nitrogen and carbon accumulation for 61 years after agricultural abandonment. Ecology 2000, 81, 88–98. [Google Scholar] [CrossRef]
- Cook, B.D.; Jastrow, J.D.; Miller, R.M. Root and mycorrhizal endophyte development in a chronosequence of restored tallgrass prairie. New Phytol. 1988, 110, 355–362. [Google Scholar] [CrossRef]
- Knight, T.R.; Dick, R.P. Differentiating microbial and stabilized β-glucosidase activity relative to soil quality. Soil Biol. Biochem. 2004, 36, 2089–2096. [Google Scholar] [CrossRef]
- Akala, V.A.; Lal, R. Soil Organic Carbon Pools and Sequestration Rates in Reclaimed Minesoils in Ohio. J. Environ. Qual. 2001, 30, 2098–2104. [Google Scholar] [CrossRef]
- Shukla, M.K.; Lal, R.; Underwood, J.; Ebinger, M. Physical and Hydrological Characteristics of Reclaimed Minesoils in Southeastern Ohio. Soil Sci. Soc. Am. J. 2004, 68, 1352–1359. [Google Scholar] [CrossRef]
- Shukla, M.K.; Lal, R.; VanLeeuwen, D. Spatial Variability of Aggregate-Associated Carbon and Nitrogen Contents in the Reclaimed Minesoils of Eastern Ohio. Soil Sci. Soc. Am. J. 2007, 71, 1748–1757. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
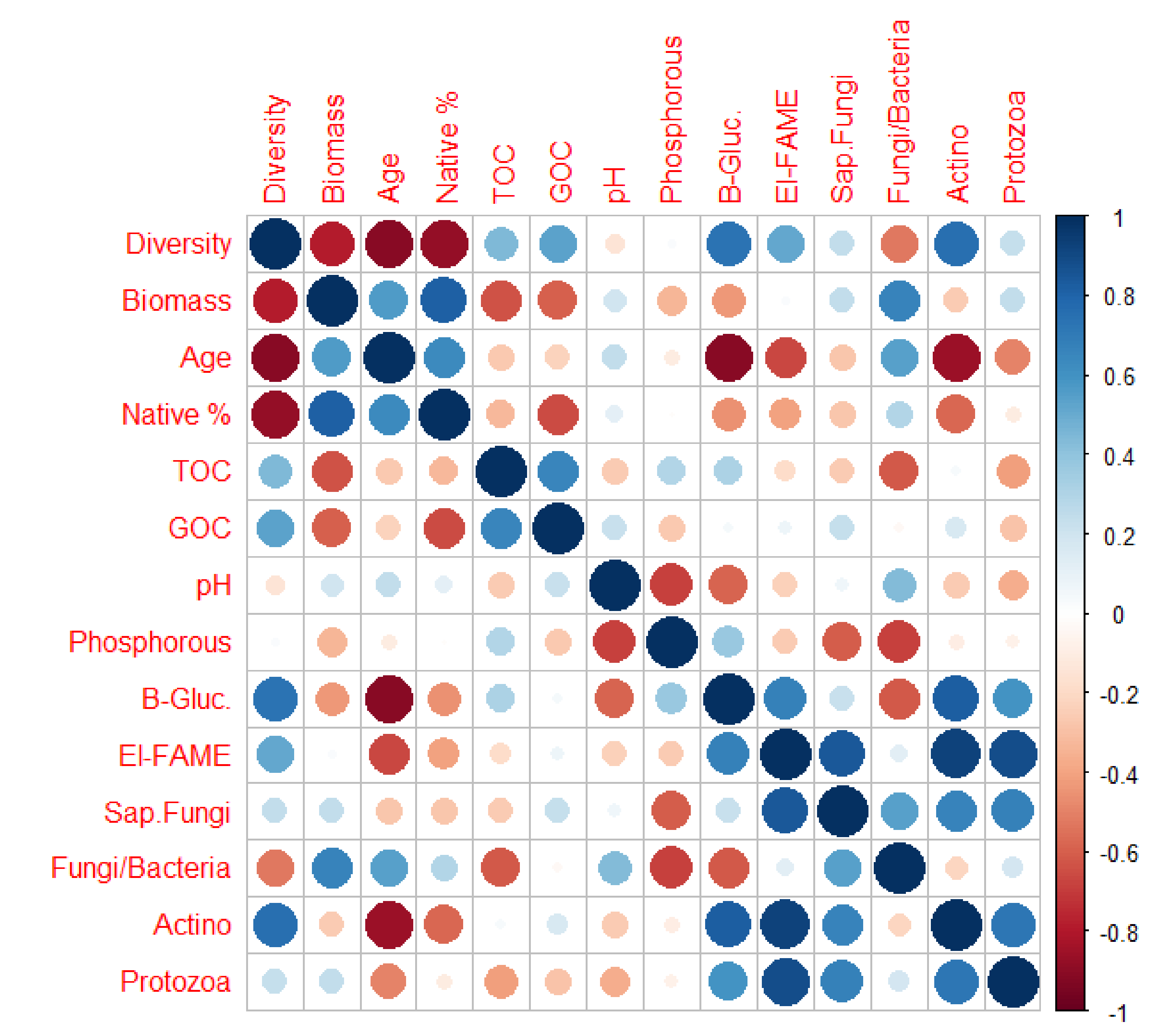{kind=link}
| Prairie Name | Grassland Type | Date Planted Groups a | Geographic Group | Reclamation Permit | Native Cover Category b | pH (0–15 cm) | pH (15–30 cm) | Total Organic Carbon (TOC) (15 cm) | TOC (15–30 cm) |
|---|---|---|---|---|---|---|---|---|---|
| Bison Base | Baseline | X | Central | B (1972–77) | A | 7.8 | 7.8 | 1.976 | 0.654 |
| Butterfly Habitat Base | Baseline | X | South | C (1977–84) | A | 7.7 | 8.0 | 2.315 | 1.248 |
| Butterfly Habitat Expansion | Prairie | B | South | C (1977–84) | C | 7.4 | 7.4 | 2.871 | 0.734 |
| Butterfly Habitat | Prairie | A | South | C (1977–84) | B | 6.8 | 7.8 | 1.93 | 0.888 |
| Bison Knoll | Prairie | C | Central | B (1972–77) | C | 7.8 | 7.9 | 2.39 | 0.634 |
| Beetle | Prairie | B | East | C (1977–84) | B | 7.7 | 7.7 | 2.155 | 0.725 |
| Dip Base | Baseline | X | East | C (1977–84) | A | 7.7 | 8.0 | 2.314 | 1.007 |
| Dip | Prairie | D | East | C (1977–84) | B | 7.6 | 7.6 | 2.277 | 0.998 |
| Hill 1 | Prairie | C | Central | B (1972–77) | A | 7.5 | 8.0 | 2.987 | 0.835 |
| Lake Trail | Prairie | A | Central | C (1977–84) | C | 7.1 | 7.1 | 2.568 | 2.197 |
| North Prairie | Prairie | A | North | B (1972–77) | C | 7.8 | 8.2 | 2.215 | 0.805 |
| Nomad Ridge | Prairie | C | South | Unmined | C | 7.6 | 8.0 | 1.883 | 0.393 |
| Overlook | Prairie | D | South | C (1977–84) | B | 7.1 | 7.8 | 3.178 | 0.663 |
| Zion Base | Baseline | X | North | B (1972–77) | A | 7.9 | 8.1 | 1.391 | 1.202 |
| Zion Prairie | Prairie | D | North | B (1972–77) | B | 7.8 | 7.8 | 2.243 | 0.602 |
- Date planted (as prairie) groups: A= 14–19 years B= 8–10 years C= 4 years D= 2 years X= Baseline (shaded: cool season).
- Native cover category: A = 6–20% B = 36–48% C = 54–93%
| 0-15 cm Soil Depth | 15-30 cm Soil Depth | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total Fungi (n mol g−1 clay) | Total El-FAME (n mol g−1 clay) | SOC (%) | Total Fungi (n mol g−1 clay) | Total El-FAME (n mol g−1 clay) | SOC (%) | |||||
| Grassland type | ||||||||||
| p value | 0.016 | 0.015 | 0.133 | 0.040 | 0.029 | 0.59 | ||||
| Baseline (cool season) | 180.24 | A1 | 483.46 | A | 1.22 | 38.80 | A | 87.13 | A | 0.33 |
| Prairie | 232.66 | B | 607.82 | B | 1.62 | 54.44 | B | 118.30 | B | 0.22 |
| Prairie age (in 2018) | ||||||||||
| p value | 0.006 | 0.065 | 0.454 | 0.0008 | 0.0004 | 0.515 | ||||
| A (14–19 years) | 282.70 | A | 667.53 | 1.37 | 77.94 | A | 155.75 | A | 0.50 | |
| B (8–10 years) | 225.81 | AB | 566.45 | 1.77 | 55.22 | AB | 116.51 | AB | 0.19 | |
| C (4 years) | 214.87 | AB | 626.01 | 1.58 | 47.57 | AB | 112.29 | AB | 0.02 | |
| D (2 years) | 204.97 | AB | 557.49 | 1.81 | 37.29 | B | 88.04 | B | 0.15 | |
| X (Baseline- not planted) | 180.24 | B | 483.46 | 1.22 | 38.80 | B | 87.13 | B | 0.33 | |
| Geographic location (on site) | ||||||||||
| p value | 0.109 | 0.009 | 0.118 | 0.20 | 0.025 | 0.746 | ||||
| Central | 235.55 | 679.31 | A | 1.57 | 56.70 | 130.69 | A | 0.37 | ||
| East | 228.19 | 583.67 | AB | 1.82 | 41.66 | 94.82 | AB | 0.30 | ||
| North | 171.05 | 468.49 | B | 1.40 | 40.98 | 80.97 | B | 0.15 | ||
| South | 227.95 | 526.25 | AB | 1.05 | 55.86 | 119.94 | AB | 0.10 | ||
| Mining Permit area | ||||||||||
| p value | 0.83 | 0.63 | 0.28 | 0.34 | 0.18 | 0.19 | ||||
| B (1972–1977) | 221.65 | 588.93 | 1.30 | 42.50 | 93.24 | 0.08 | ||||
| C (1977–1982) | 217.02 | 565.21 | 1.69 | 56.23 | 123.14 | 0.40 | ||||
| Unmined | 214.16 | 564.51 | 1.39 | 49.26 | 105.28 | 0.04 | ||||
| Proportion Native Vegetation in Grassland | ||||||||||
| p value | 0.003 | 0.03 | 0.6 | 0.002 | 0.003 | 0.788 | ||||
| A 0–25% | 184.32 | A | 513.64 | 1.35 | 40.13 | A | 92.35 | A | 0.26 | |
| B 25–50% | 209.73 | A | 555.53 | 1.64 | 44.49 | A | 97.68 | A | 0.16 | |
| C >50% | 261.98 | B | 654.80 | 1.55 | 66.18 | B | 139.93 | B | 0.32 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swab, R.M.; Lorenz, N.; Lee, N.R.; Culman, S.W.; Dick, R.P. From the Ground Up: Prairies on Reclaimed Mine Land—Impacts on Soil and Vegetation. Land 2020, 9, 455. https://doi.org/10.3390/land9110455
Swab RM, Lorenz N, Lee NR, Culman SW, Dick RP. From the Ground Up: Prairies on Reclaimed Mine Land—Impacts on Soil and Vegetation. Land. 2020; 9(11):455. https://doi.org/10.3390/land9110455
Chicago/Turabian StyleSwab, Rebecca M., Nicola Lorenz, Nathan R. Lee, Steven W. Culman, and Richard P. Dick. 2020. "From the Ground Up: Prairies on Reclaimed Mine Land—Impacts on Soil and Vegetation" Land 9, no. 11: 455. https://doi.org/10.3390/land9110455
APA StyleSwab, R. M., Lorenz, N., Lee, N. R., Culman, S. W., & Dick, R. P. (2020). From the Ground Up: Prairies on Reclaimed Mine Land—Impacts on Soil and Vegetation. Land, 9(11), 455. https://doi.org/10.3390/land9110455