The Ecosystem Effects of Sand-Binding Shrub Hippophae rhamnoides in Alpine Semi-Arid Desert in the Northeastern Qinghai–Tibet Plateau


Abstract
1. Introduction
2. Materials and Methods
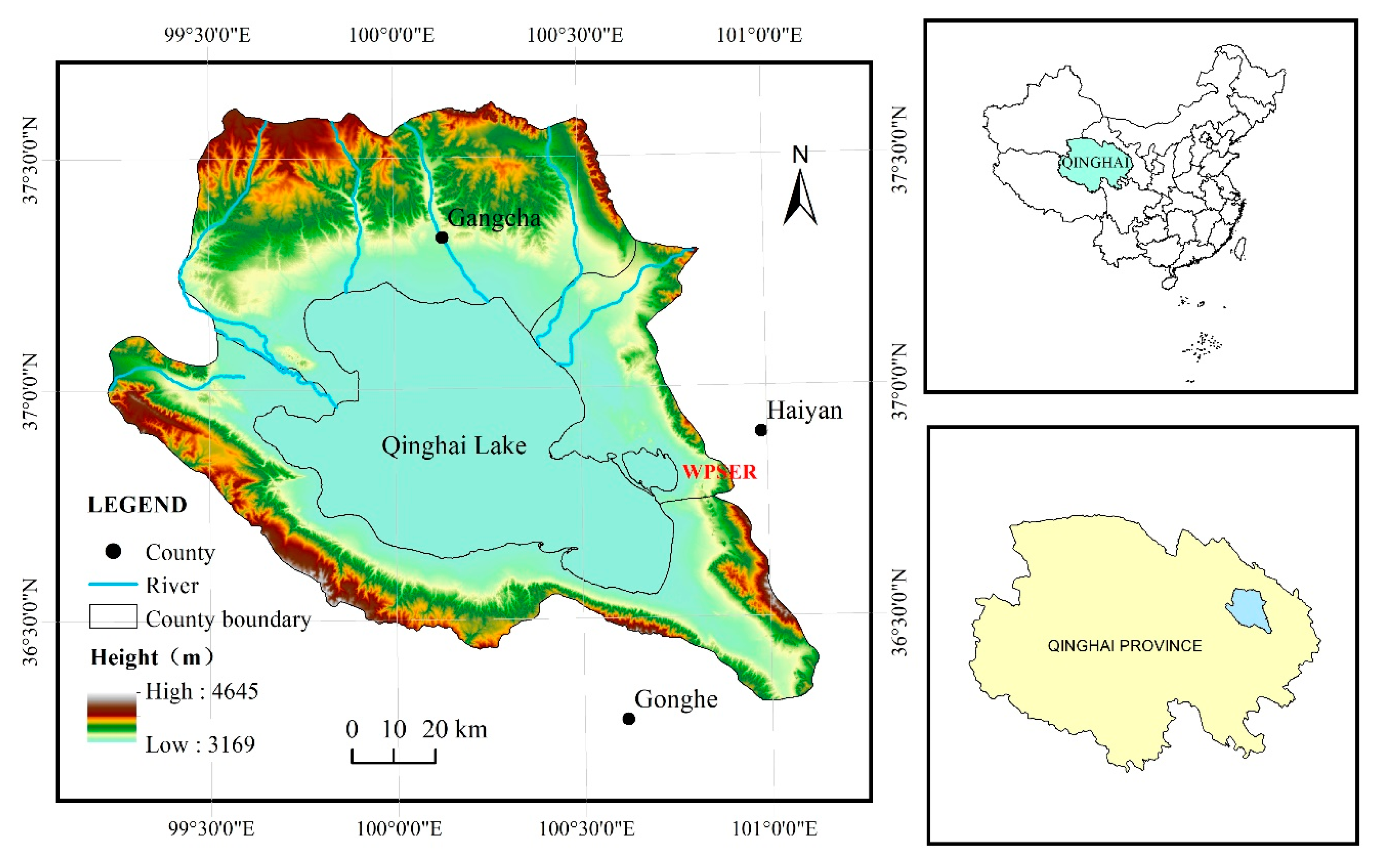
2.1. Study Area
2.2. Plant Feature Investigation and Analysis
2.3. Aeolian Activity Investigation and Analysis
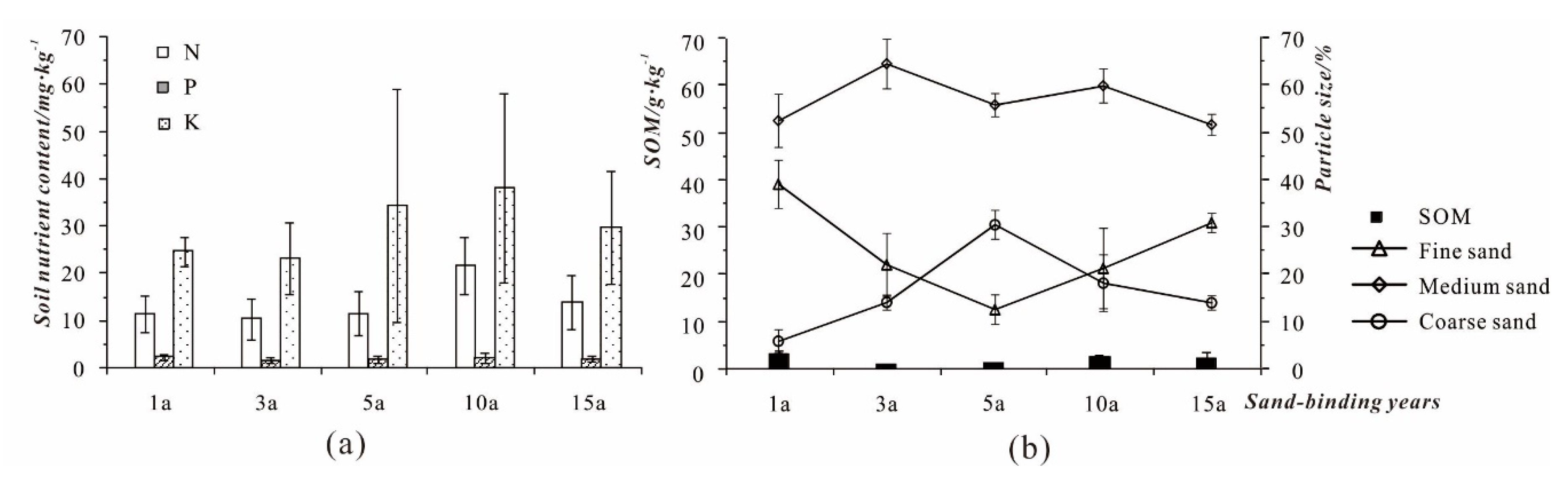
2.4. Soil Sample Collection and Analysis
2.5. Data Analysis
3. Results
3.1. Ecological Features of Aged H. rhamnoides Communities
3.1.1. Community Composition
3.1.2. Community Diversity
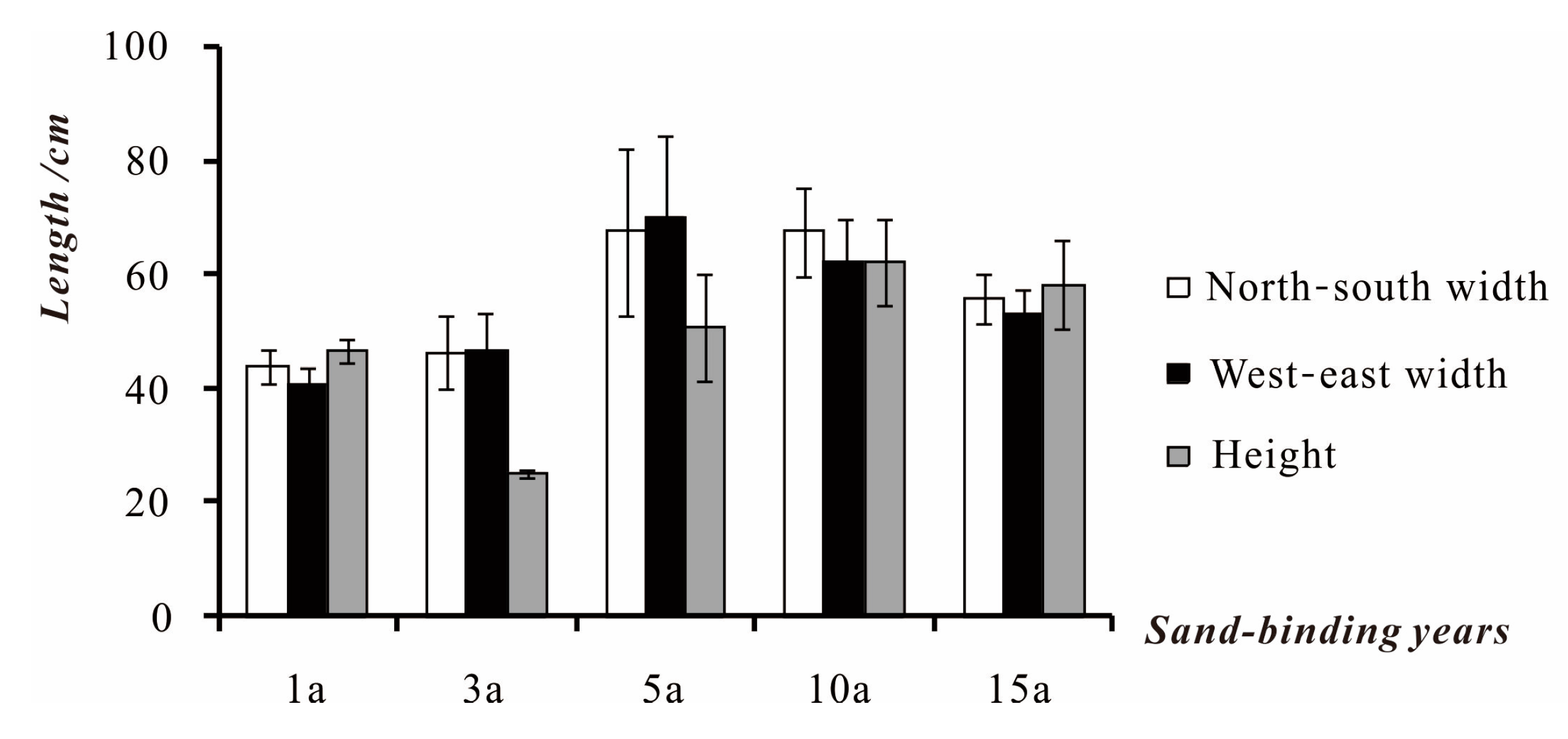
3.1.3. Community Structure
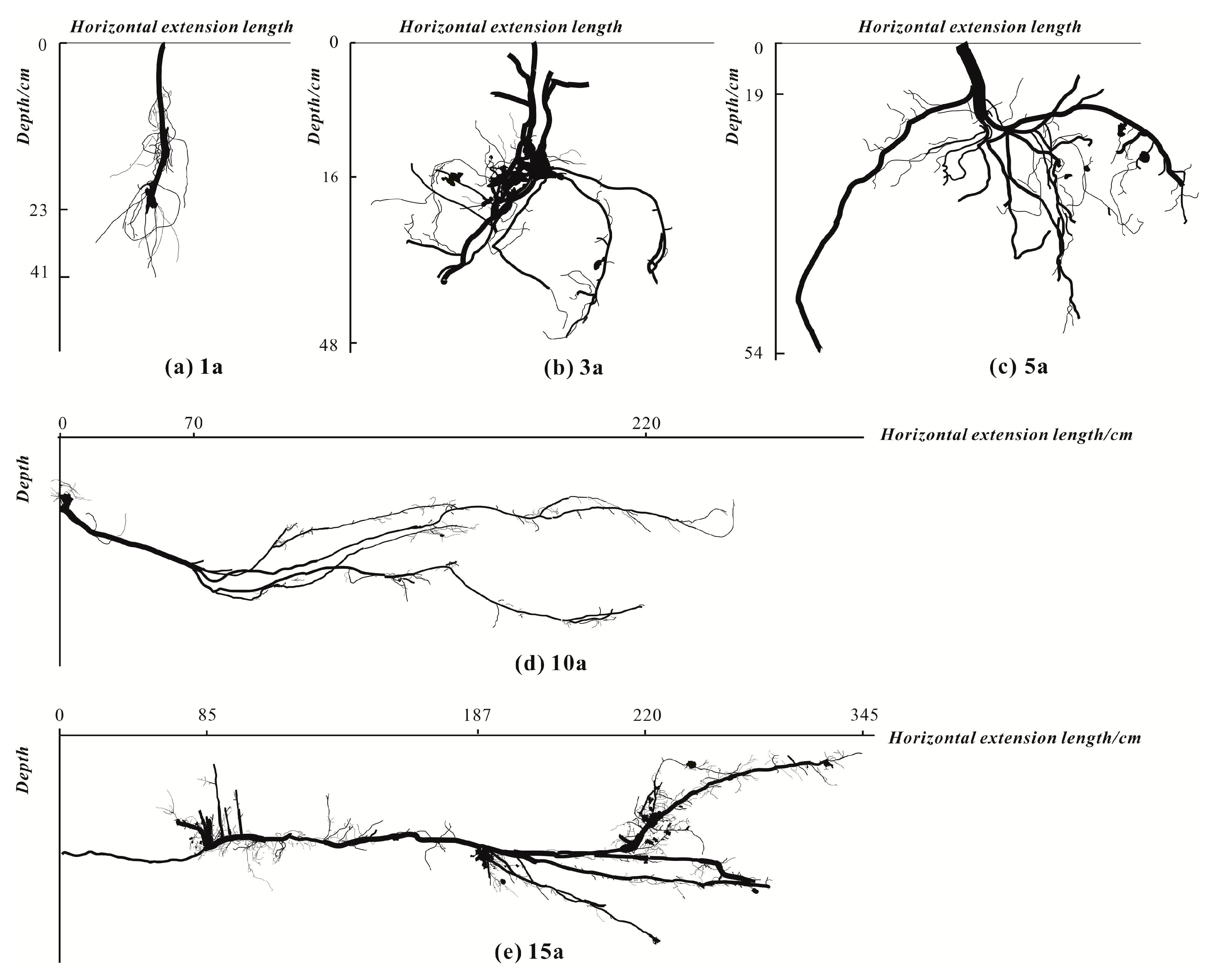
3.1.4. Community Biomass and Root Distribution
3.2. Aeolian Activities of H. rhamnoides Communities
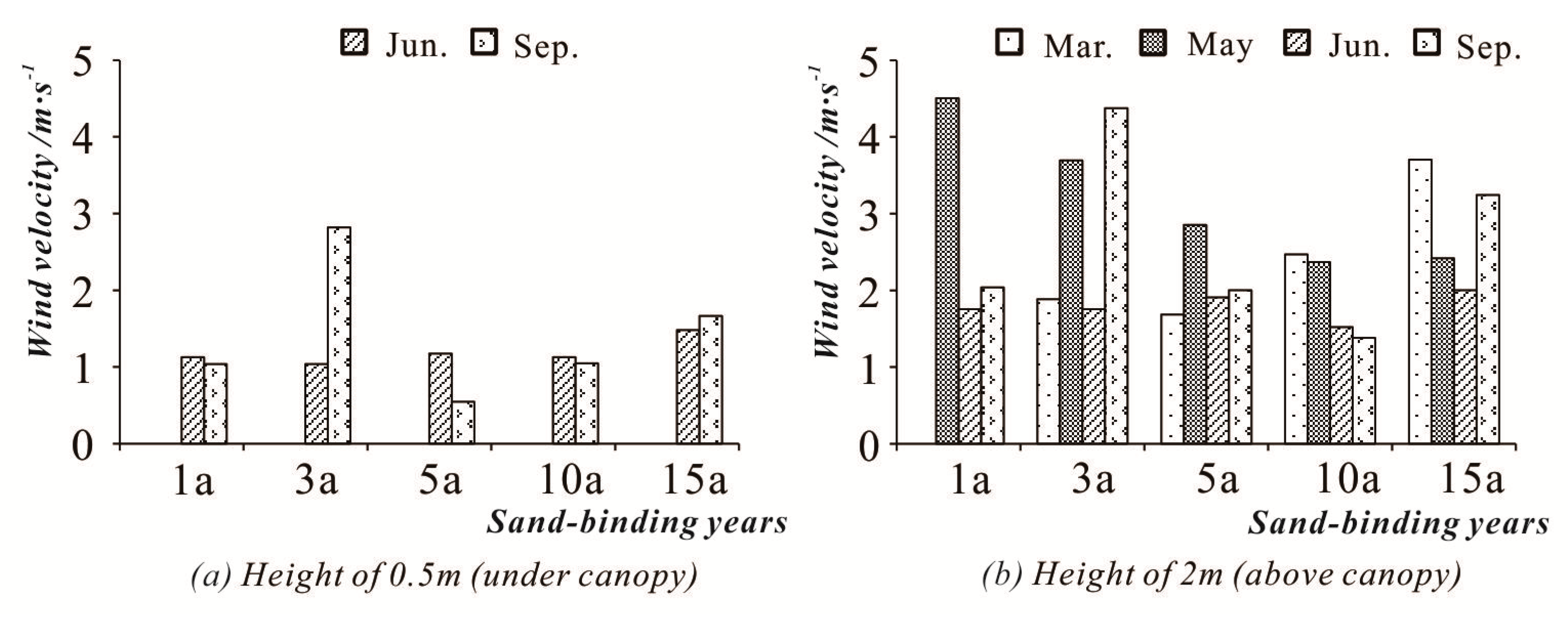
3.2.1. Wind Speed Characteristics of the Study Sites
3.2.2. Vertical Wind Increase Rate (R)
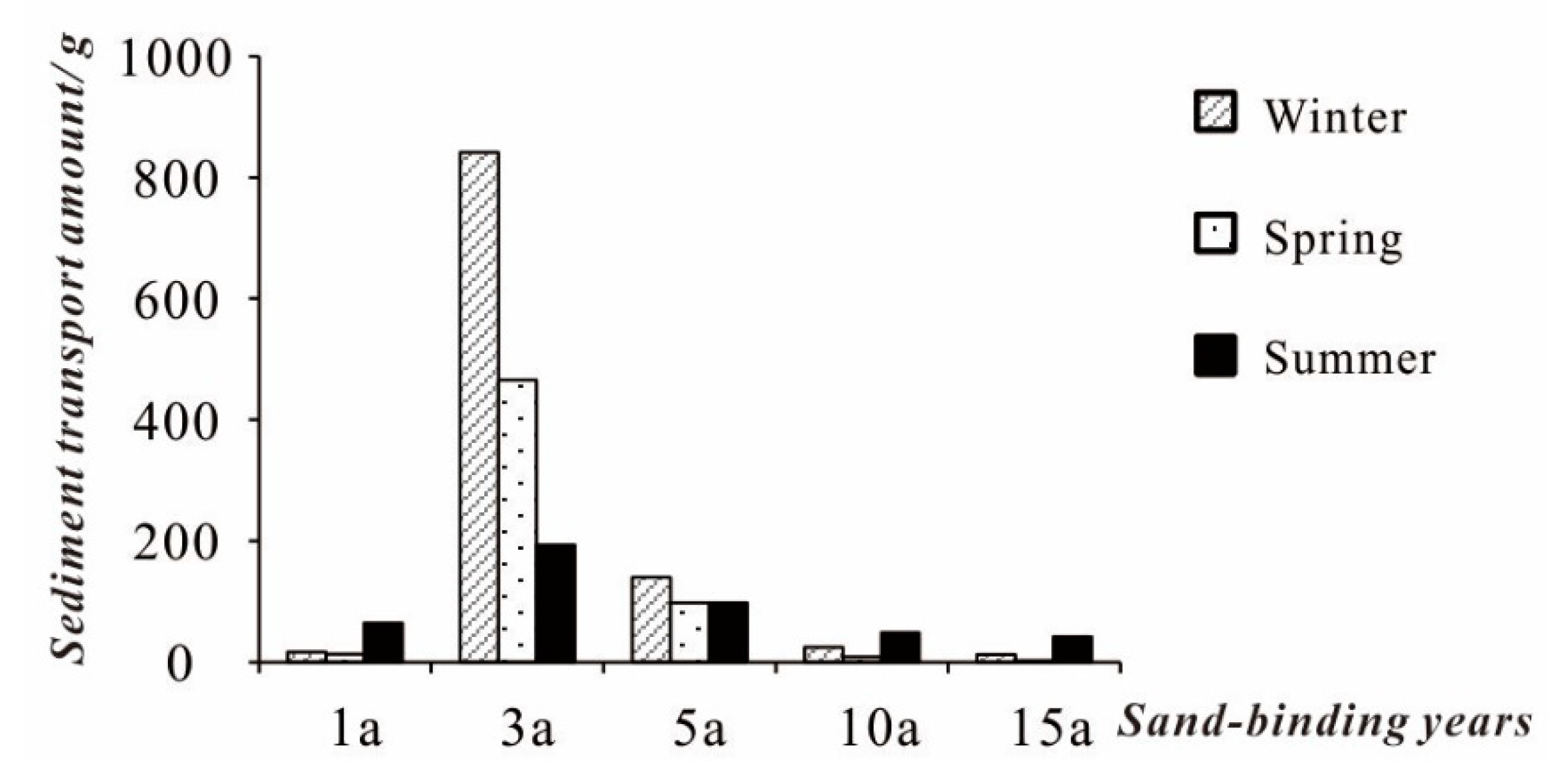
3.2.3. Sediment Transport
4. Discussion
4.1. H. rhamnoides Community Evolution
4.2. Effects of Aeolian Activities on H. rhamnoides Communities
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sperry, J.S.; Hacke, U.G. Desert shrub water relations with respect to soil characteristics and plant functional type. Funct. Ecol. 2002, 16, 367–378. [Google Scholar] [CrossRef]
- D’Odorico, P.; Bhattachan, A.; Davis, K.F.; Ravi, S.; Runyan, C.W. Global desertification: Drivers and feedbacks. Adv. Water Resour. 2013, 51, 326–344. [Google Scholar] [CrossRef]
- Qi, Y.; Chen, T.; Pu, J.; Yang, F.; Shukla, M.K.; Chang, Q. Response of soil physical, chemical and microbial biomass properties to land use changes in fixed desertified land. Catena 2018, 160, 339–344. [Google Scholar] [CrossRef]
- Yu, Y.; Jia, Z.Q. Changes in soil organic carbon and nitrogen capacities of Salix cheilophila Schneid. along a revegetation chronosequence in semi-arid degraded sandy land of the Gonghe Basin, Tibetan Plateau. Solid Earth 2014, 5, 1045–1054. [Google Scholar] [CrossRef]
- Li, X.R.; Zhou, H.Y.; Wang, X.P.; Liu, L.C.; Zhang, J.G.; Chen, G.X.; Zhang, Z.S.; Liu, Y.B.; Tan, H.J.; Gao, Y.H. Ecological restoration and recovery in arid desert regions of China: A review for 60-year research progresses of Shapotou Desert Research and Experiment Station, Chinese Academy of Sciences. J. Desert Res. 2016, 36, 247–264. [Google Scholar]
- Pearson, M.; Rogers, J. Hippophae rhamnoides L. J. Ecol. 1962, 50, 501–513. [Google Scholar] [CrossRef]
- Richards, E.; Burningham, H. Hippophae rhamnoides on a coastal dune system: A thorny issue? J. Coast. Conserv. 2011, 15, 73–85. [Google Scholar] [CrossRef]
- Chen, B.B.; Zeng, C.; Gao, H.Y.; Liu, C.H.; Li, G.Q.; Dai, G.H. Response of persistence of Hippophae rhamnoides L. to soil water contents. J. Cent. South Univ. For. Technol. 2016, 36, 67–71. [Google Scholar]
- Zuazo, V.H.D.; Pleguezuelo, C.R.R. Soil-erosion and runoff prevention by plant covers: A review. Sustain. Agric. 2008, 28, 65–86. [Google Scholar]
- Brown, J.F. Effects of experimental burial on survival, growth and resource allocation of three species of dune plants. J. Ecol. 1997, 85, 151. [Google Scholar] [CrossRef]
- Li, X.R.; Zhang, Z.S.; Huang, L.; Wang, X. Review of the ecohydrological processes and feedback mechanisms controlling sand-binding vegetation systems in sandy desert regions of China. Chin. Sci. Bull. 2013, 58, 1483–1496. [Google Scholar] [CrossRef]
- Yu, Y.J.; Shi, P.J.; He, L.P. Research on the effects of wind-sand current on the plant growth. Adv. Earth Sci. 2002, 17, 262–267. [Google Scholar]
- Maun, M.A.; Lapierre, J. The effects of burial by sand on Ammophila breviligulata. J. Ecol. 1984, 72, 827–839. [Google Scholar] [CrossRef]
- Zhang, J.H.; Maun, M.A. Effects of sand burial on seed germination, seedling emergence, survival, and growth of Agropyronp sammophilum. Can. J. Bot. 1990, 68, 304–310. [Google Scholar] [CrossRef]
- Wu, W.Y.; Zhang, D.S.; Tian, L.H.; Zhang, M.Y.; Zhou, X. Ecological responses of Hippophae rhamnoides to wind-sand hazard in alpine sand land. Bull. Soil Water Conserv. 2018, 38, 1–8. [Google Scholar]
- Zhang, D.S.; Tian, L.H.; Lu, R.J.; Tang, Q.L.; Shi, H. Grain-size features of aeolian deposits in the eastern shore of Qinghai Lake. Arid Land Geogr. 2013, 36, 203–211. [Google Scholar]
- Wu, W.Y.; Zhang, D.S.; Tian, L.H.; Zhang, M.Y.; Zhou, X. Features of artificial plant communities from the east sand region of the Qinghai Lake over the last 10 years. Acta Ecol. Sin. 2019, 39, 2109–2121. [Google Scholar]
- Wu, H.W.; Li, X.Y.; Jiang, Z.Y.; Chen, H.Y.; Zhang, C.C.; Xiao, X. Contrasting water use pattern of introduced and native plants in an alpine desert ecosystem, northeast Qinghai-Tibet Plateau, China. Sci. Total Environ. 2016, 542, 182–191. [Google Scholar] [CrossRef]
- Wu, Z. Geomorphology of Wind-Drift Sands and Their Controlled Engineering; China Science Publishing & Media Ltd.: Beijing, China, 2003; pp. 61–86. [Google Scholar]
- Odum, E.P.; Barrett, G.W. Fundementals of Ecology, 5th ed.; Brooks/Cole: New York, NY, USA, 2005; pp. 33–35. [Google Scholar]
- Liu, H.T. Study on eco-physiological characteristics of Salix Cheilophila and Process of Vegetation Restoration in Alpine-Cold Sandy Land; Chinese Academy of Forestry: Beijing, China, 2012. [Google Scholar]
- Ma, Q.L.; Lu, Q.; Zhang, D.K.; Liu, H.J.; Jin, H.J. Wind prevention and sand resistance effects of Artemisia sphaerocephala and A. ordosica. Chin. J. Ecol. 2012, 31, 1639–1645. [Google Scholar]
- Grimm, V.; Wissel, C. The ecological stability discussions: An inventory and analysis of terminology and a guide for avoiding confusion. Ecology 1997, 109, 323–334. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity–stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y. Redundancy theory and its application to Ecology. J. Nantong Univ. 2006, 5, 55–59. [Google Scholar]
- Cao, C.; Zhu, L.; Jiang, D.; Fan, S. Discussion on Mechanism of Stability of Sand-fixing Phytocoenosium. J. Desert Res. 2004, 24, 461–466. [Google Scholar]
- Hill, A.R. Ecosystem stability in relation to stresses caused by human activities. Can. Geogr. 1975, 19, 206–220. [Google Scholar] [CrossRef]
- Li, R.X.; Xue, Q.H.; Yang, S.Y.; Feng, L.X.; Zhou, X.S. Improving effect of Seabuckthorn and Black Locust artificial forests to soil fertility in Loess Plateau and its model. J. Soil Eros. Soil Water Conserv. 1998, 4, 14–21. [Google Scholar]
- Li, G.Q.; Tang, R.D.; Zhao, Y.Q. The Biological property and ecological habit of Hippophae. Acta Bot. Boreali Occident. Sin. 2000, 20, 892–897. [Google Scholar]
- Wang, L.; Zhang, X.M.; Lin, M.Z.; Zhang, J.K. Study on root and function of Hippophae I—Root differentiation and division. Hippophae 2005, 4, 13–19. [Google Scholar]
- Song, C.Y.; Guo, K.; Liu, G.H. Relationships between plant community’s species diversity and soil factors on otingdag sandy land. Chin. J. Ecol. 2008, 27, 8–13. [Google Scholar]
- Li, Y.H.; Luo, T.X.; Lu, Q.; Tian, X.Y.; Wu, B.; Yang, H.H. Comparisons of leaf traits among 17 major plant species in shazhuyu sand control experimental station of Qinghai Province. Acta Ecol. Sin. 2005, 25, 994–999. [Google Scholar]
- Yang, Z.W. Discussion on three modes of desertification control in Qinghai alpine arid area. Sci. Technol. Qinghai Agric. For. 2004, 3, 42–45. [Google Scholar]
- Lu, Z.X.; Meng, H.J.; Yan, C.M. Study on the growth of Seabuckthorn and its improvement of soil properties. Gansu Sci. Technol. 2016, 10, 226–227. [Google Scholar]
- Walter, B.; Voegeli, C.; Horender, S. Estimating sediment mass fluxes on surfaces sheltered by live vegetation. Bound. Layer Meteorol. 2017, 163, 273–286. [Google Scholar] [CrossRef]
- Li, S.L.; Werger, M.J.; Zuidema, P.A.; Yu, F.H.; Dong, M. Seedlings of the Semi-shrub Artemisia ordosica are Resistant to Moderate Wind Denudation and Sand Burial in Mu Us Sandland, China. Trees 2010, 24, 515–521. [Google Scholar] [CrossRef]
- Danjon, F.; Fourcaud, T.; Bert, D. Root architecture and wind-firmness of mature Pinus pinaster. New Phytol. 2005, 168, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, B.C.; Ray, D. Adaptive growth of tree root systems in response to wind action and site conditions. Tree Physiol. 1996, 16, 891–898. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Planting Time | Original Topography | Existing Topography | Dune Height /m | Mechanical Barriers | Afforestation Mode |
|---|---|---|---|---|---|---|
| 1a | 2016 | Mobile dune | Mobile dune | 10.3 | Barley-oat checkerboards | Seedlings |
| 3a | 2014 | Mobile dune | Mobile dune | 8.7 | Straw checkerboards | Seedlings |
| 5a | 2012 | Mobile dune | Semi-fixed dune | 7.2 | Straw checkerboards | Seedlings |
| 10a | 2008 | Mobile dune | Fixed dune | 10.6 | Straw checkerboards | Seedlings |
| 15a | 2003 | Mobile dune | Fixed dune | 5.9 | Straw checkerboards | Seedlings |
| LSD | Natural | Mobile dune | Mobile dune | — | — | — |
| Family | Species |
|---|---|
| Aphididae | H. rhamnoides |
| Compositae | Artemisia desertorum; Helianthus tuberosus; Sonchus oleraceus |
| Willow | Populus simonii; Salix cheilophila |
| Matsuko | Pinus sylvestrisvar |
| Legume | Caragana intermedia; Oxytropis imbricata |
| Gramineae | Agropyron cristatum; Poa annua; Leymus secalinus; Elymus nutans Avena sativa |
| Diversity Index | 1a | 3a | 5a | 10a | 15a |
|---|---|---|---|---|---|
| H’ | 0.368a | 0.367a | 0.365a | 0.358a | 0.368a |
| D | 0.597a | 0.658a | 0.584a | 0.541a | 0.634a |
| J | 0.189a | 0.205a | 0.175a | 0.172a | 0.177a |
| Community | Total Coverage | Shrubs | Coverage | Herbs |
|---|---|---|---|---|
| 1a | 19.33 | H. rhamnoides | 9.33 | A. sativa |
| A. desertorum | 4.00 | |||
| P. simonii | 2.00 | |||
| S. cheilophila | 1.00 | |||
| H. tuberosus | 3.00 | |||
| 3a | 21.00 | H. rhamnoides | 14.33 | A.cristatum, O. imbricate, E.nutans |
| S. cheilophila | 1.00 | |||
| A. desertorum | 5.67 | |||
| 5a | 45.00 | H. rhamnoides | 22.00 | A.cristatum, O. imbricate, L. secalinus |
| P. simonii | 15.00 | |||
| S. cheilophila | 2.00 | |||
| P. sylvestrisvar | 4.00 | |||
| A. desertorum | 2.00 | |||
| 10a | 60.33 | H. rhamnoides | 45.33 | A.cristatum, O. imbricate, L. secalinus, P. annua |
| P. sylvestrisvar | 11.00 | |||
| C. intermedia | 2.00 | |||
| A. desertorum | 2.00 | |||
| 15a | 65.04 | H. rhamnoides | 36.70 | A.cristatum, O. imbricate, L. secalinus, P. annua |
| P. sylvestrisvar | 2.67 | |||
| A. desertorum | 25.67 |
| Aged | Height/cm | Canopy/cm | Root Height/cm | Branch Roots | Depth of Roots/cm | Above Biomass/g | Under Biomass/g | Root-Shoot Ratio | |
|---|---|---|---|---|---|---|---|---|---|
| 1a | 40 | 32 | 25 | - | 1 | 30–50 | 71.30 | 28.01 | 0.39 |
| 3a | 46 | 45 | 50 | 55 | 4 | 30–80 | 285.33 | 175.00 | 0.61 |
| 5a | 65 | 110 | 90 | 90 | 5 | 90–150 | 275.89 | 242.76 | 0.88 |
| 10a | 63 | 69 | 90 | 80 | 8 | 90–260 | 453.96 | 885.26 | 1.95 |
| 15a | 54 | 103 | 78 | 55 | 6 | 100–340 | 865.22 | 826.97 | 0.96 |
| Height Layer/m | LSD 1 | 1a 2 | 3a 2 | 5a 2 | 10a 2 | 15a 2 |
|---|---|---|---|---|---|---|
| 0–1 | 0.62 | 1.66 | 2.39 | 0.04 | 0.61 | 1.26 |
| 1–2 | 2.00 | 0.29 | 0.36 | 0.97 | 0.45 | 1.62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, L.; Wu, W.; Zhou, X.; Zhang, D.; Yu, Y.; Wang, H.; Wang, Q. The Ecosystem Effects of Sand-Binding Shrub Hippophae rhamnoides in Alpine Semi-Arid Desert in the Northeastern Qinghai–Tibet Plateau. Land 2019, 8, 183. https://doi.org/10.3390/land8120183
Tian L, Wu W, Zhou X, Zhang D, Yu Y, Wang H, Wang Q. The Ecosystem Effects of Sand-Binding Shrub Hippophae rhamnoides in Alpine Semi-Arid Desert in the Northeastern Qinghai–Tibet Plateau. Land. 2019; 8(12):183. https://doi.org/10.3390/land8120183
Chicago/Turabian StyleTian, Lihui, Wangyang Wu, Xin Zhou, Dengshan Zhang, Yang Yu, Haijiao Wang, and Qiaoyu Wang. 2019. "The Ecosystem Effects of Sand-Binding Shrub Hippophae rhamnoides in Alpine Semi-Arid Desert in the Northeastern Qinghai–Tibet Plateau" Land 8, no. 12: 183. https://doi.org/10.3390/land8120183
APA StyleTian, L., Wu, W., Zhou, X., Zhang, D., Yu, Y., Wang, H., & Wang, Q. (2019). The Ecosystem Effects of Sand-Binding Shrub Hippophae rhamnoides in Alpine Semi-Arid Desert in the Northeastern Qinghai–Tibet Plateau. Land, 8(12), 183. https://doi.org/10.3390/land8120183