
Enhancement of Linear Agricultural Areas to Provide Invertebrates as Potential Food for Breeding Birds

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
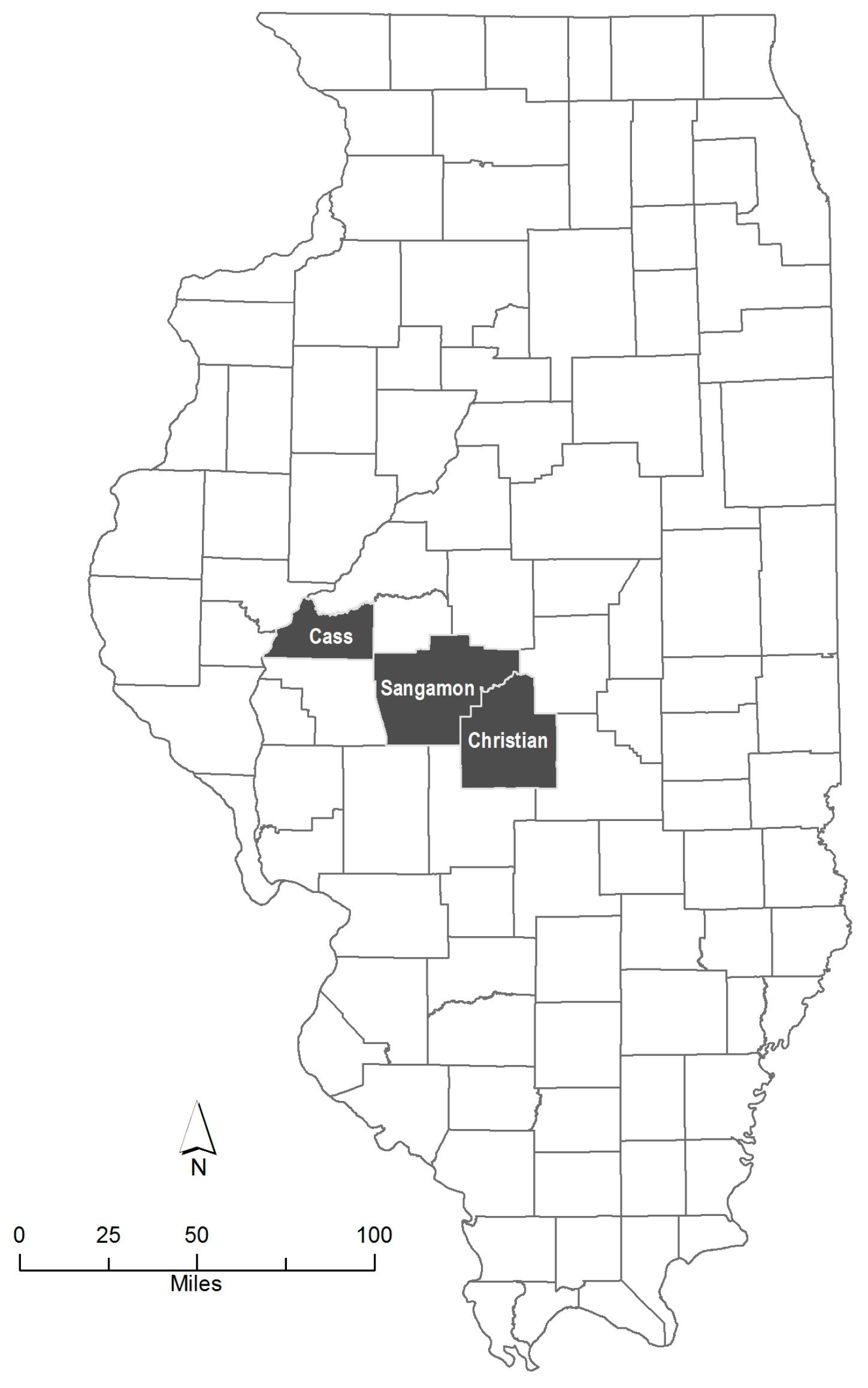
2.1. Study Area
2.2. Sampling Methods
2.3. Identification of Invertebrates
2.4. Data Analysis
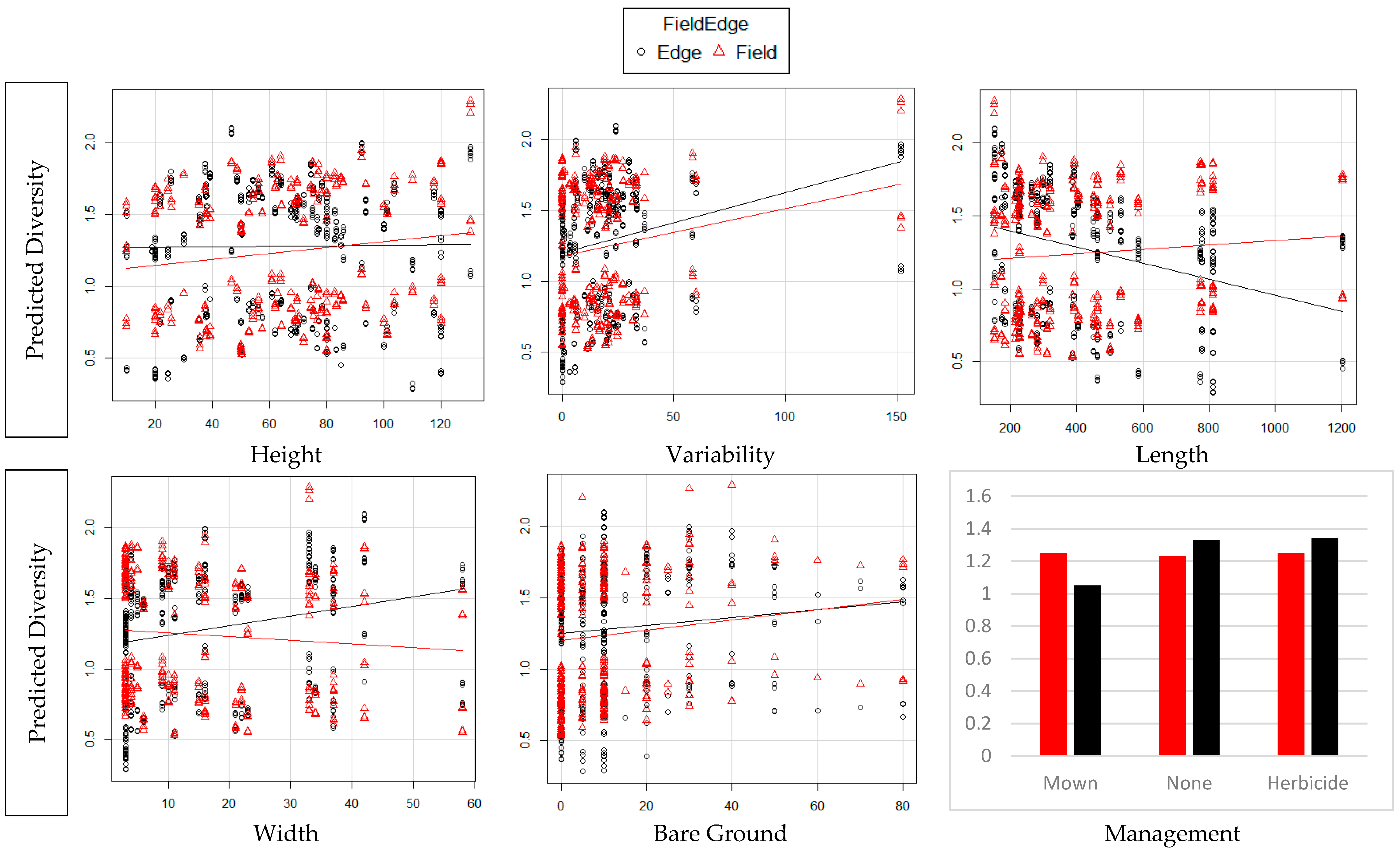
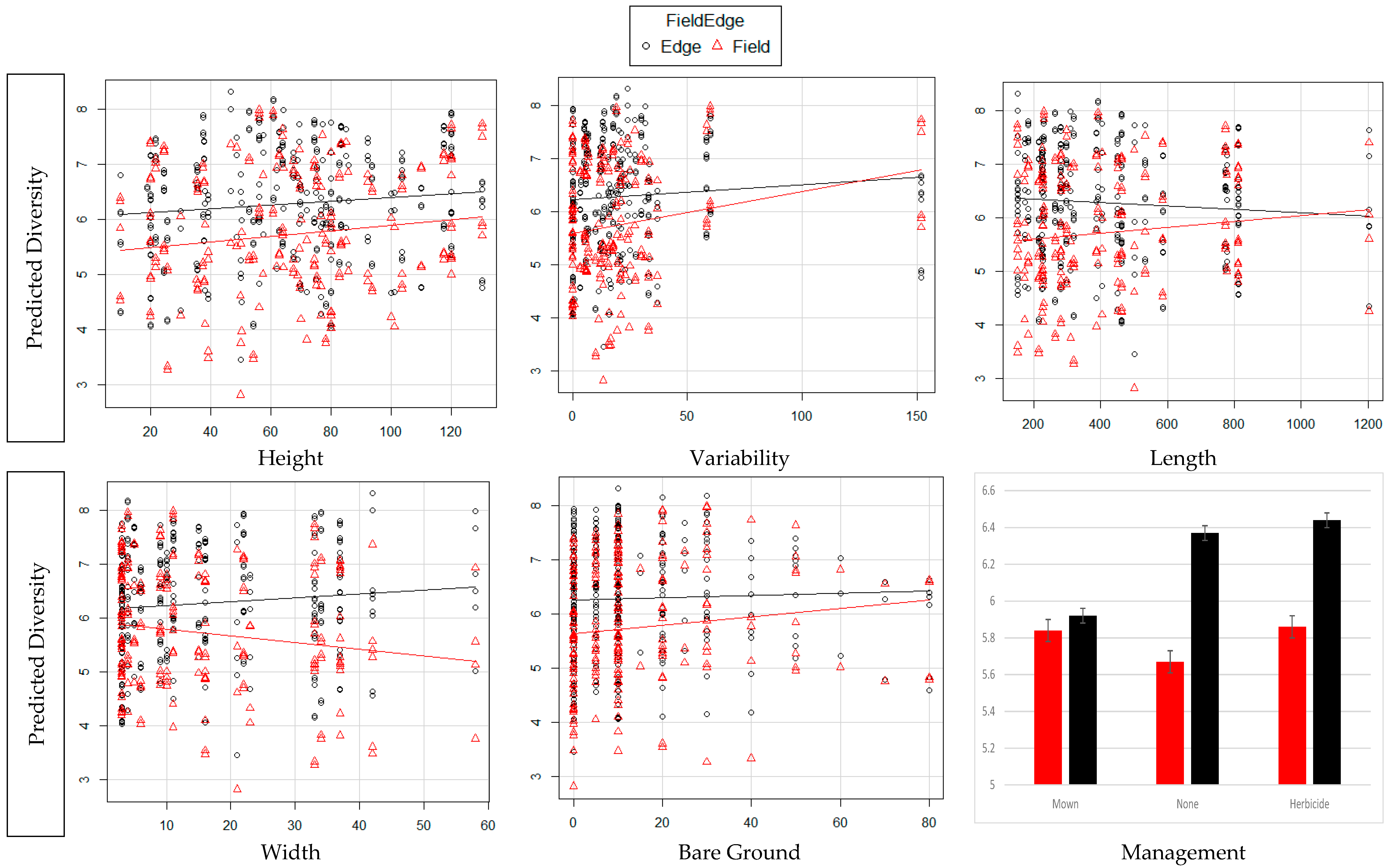
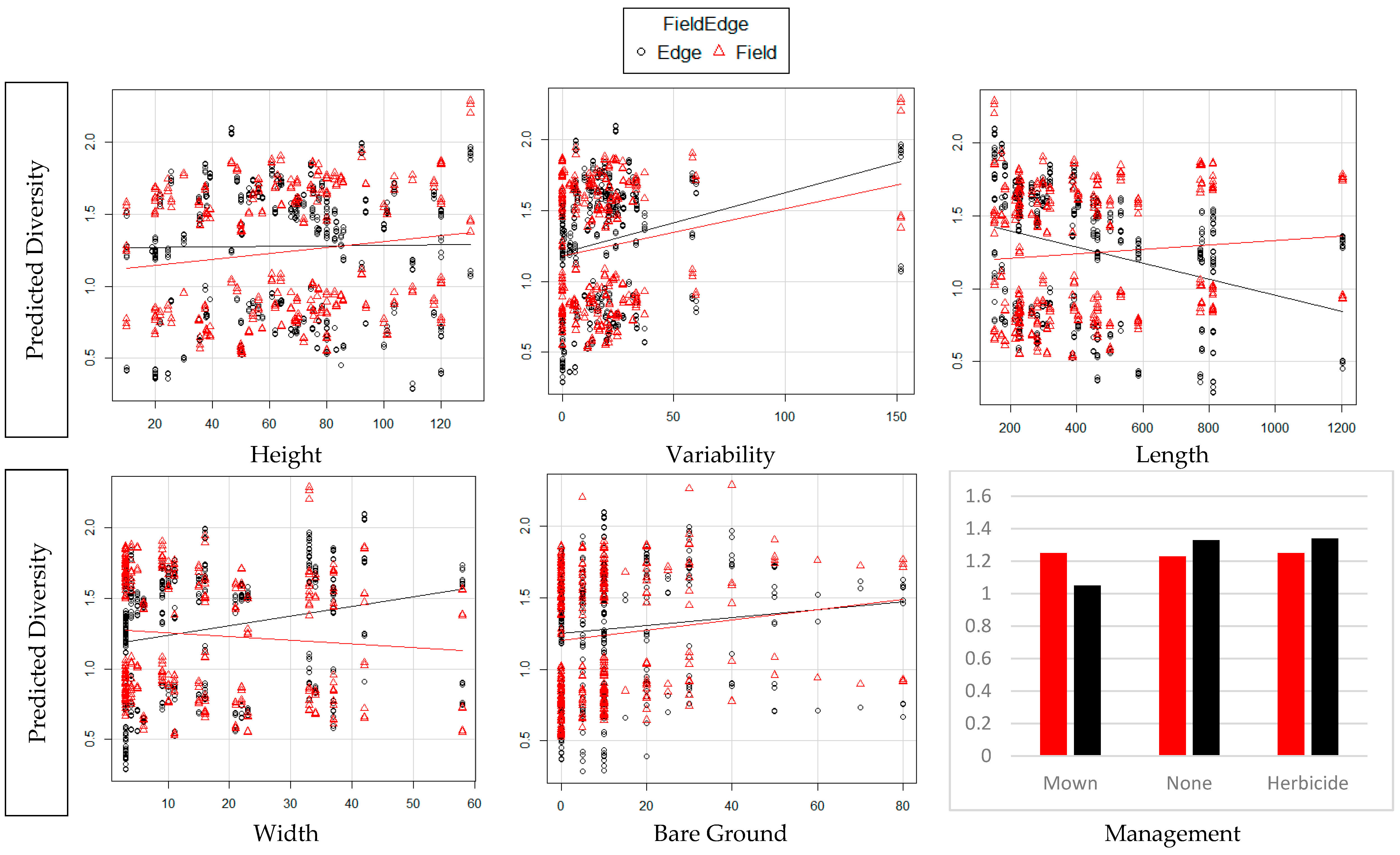
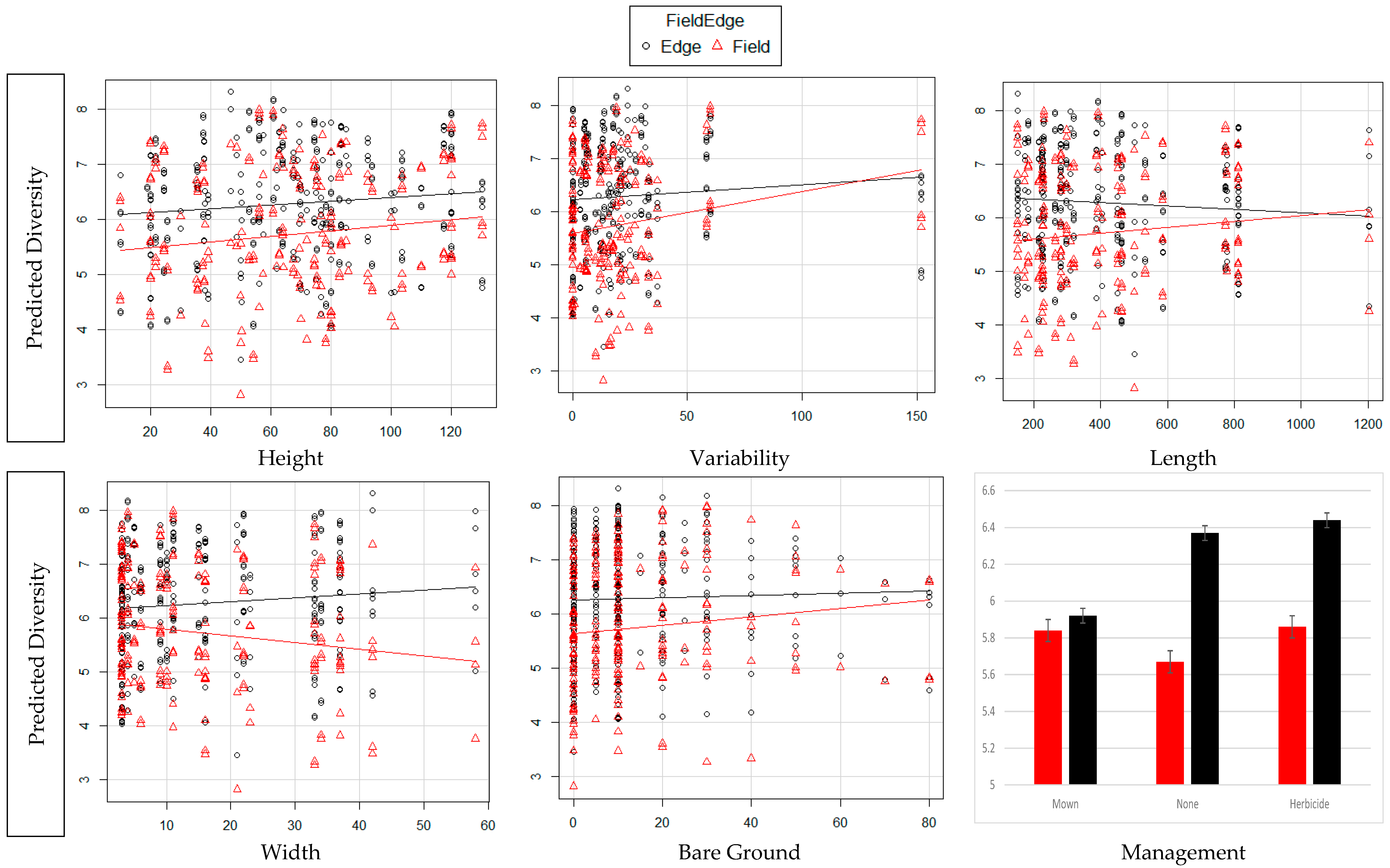
3. Results
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Walk, J.W.; Warwick, C. Illinois Birds: A Century of Change; Illinois Natural History Survey: Champaign, IL, USA, 2010. [Google Scholar]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C. Persistent negative effects of pesticides on biodiversity and biological control potential on european farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Phalan, B.; Onial, M.; Balmford, A.; Green, R.E. Reconciling food production and biodiversity conservation: Land sharing and land sparing compared. Science 2011, 333, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Grau, R.; Kuemmerle, T.; Macchi, L. Beyond ‘land sparing versus land sharing’: Environmental heterogeneity, globalization and the balance between agricultural production and nature conservation. Curr. Opin. Environ. Sustain. 2013, 5, 477–483. [Google Scholar] [CrossRef]
- Donald, P.F.; Evans, A.D. Habitat connectivity and matrix restoration: The wider implications of agri-environment schemes. J. Appl. Ecol. 2006, 43, 209–218. [Google Scholar] [CrossRef]
- Vickery, J.A.; Bradbury, R.B.; Henderson, I.G.; Eaton, M.A.; Grice, P.V. The role of agri-environment schemes and farm management practices in reversing the decline of farmland birds in england. Biol. Conserv. 2004, 119, 19–39. [Google Scholar] [CrossRef]
- Carvell, C.; Meek, W.R.; Pywell, R.F.; Goulson, D.; Nowakowski, M. Comparing the efficacy of agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins. J. Appl. Ecol. 2007, 44, 29–40. [Google Scholar] [CrossRef]
- Anderson, R.C. Prairies in the prairie state. Trans. Ill. State Acad. Sci. 1970, 63, 214–221. [Google Scholar]
- Ries, L.; Fletcher, R.J., Jr.; Battin, J.; Sisk, T.D. Ecological responses to habitat edges: Mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 491–522. [Google Scholar] [CrossRef]
- Cavitt, J.F.; Thompson, C.F. Mass loss in breeding house wrens: Effects of food supplements. Ecology 1997, 78, 2512–2523. [Google Scholar] [CrossRef]
- Bell, G.P. Birds and mammals on an insect diet: A primer on diet composition analysis in relation to ecological energetics. Stud. Avian Biol. 1990, 13, 416–422. [Google Scholar]
- Batáry, P.; Báldi, A.; Szél, G.; Podlussány, A.; Rozner, I.; Erdős, S. Responses of grassland specialist and generalist beetles to management and landscape complexity. Divers. Distrib. 2007, 13, 196–202. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Tscharntke, T. Insect communities and biotic interactions on fragmented calcareous grasslands—A mini review. Biol. Conserv. 2002, 104, 275–284. [Google Scholar] [CrossRef]
- Westerman, P.; Hofman, A.; Vet, L.; van der Werf, W. Relative importance of vertebrates and invertebrates in epigeaic weed seed predation in organic cereal fields. Agric. Ecosyst. Environ. 2003, 95, 417–425. [Google Scholar] [CrossRef]
- Marvier, M.; McCreedy, C.; Regetz, J.; Kareiva, P. A meta-analysis of effects of Bt cotton and maize on nontarget invertebrates. Science 2007, 316, 1475–1477. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.D.; Morris, A.J.; Arroyo, B.E.; Clark, S.C.; Bradbury, R.B. A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern europe in relation to agricultural change. Agric. Ecosyst. Environ. 1999, 75, 13–30. [Google Scholar] [CrossRef]
- Stinner, B.R.; House, G. Arthropods and other invertebrates in conservation-tillage agriculture. Annu. Rev. Entomol. 1990, 35, 299–318. [Google Scholar] [CrossRef]
- Schwegman, J.E. The Natural Divisions of Illinois; Illinois Nature Preserves Commission: Springfield, IL, USA, 1973. [Google Scholar]
- Eymann, J. Manual on field recording techniques and protocols for all taxa biodiversity inventories and monitoring. Abc Taxa 2010, 8. Part 1. Available online: http://www.abctaxa.be/ (accessed on 21 October 2011). [Google Scholar]
- Hendron, L.M. Do Arthropod Abundance and Diversity Differ between Grass Habitats Varying in Height? Master’s Thesis, University of Illinois, Urbana, IL, USA, 2010. [Google Scholar]
- Graber, R.R.; Graber, J.W. A comparative study of bird populations in Illinois, 1906–1909 and 1956–1958. Ill. Nat. Hist. Surv. Bull. 1963, 28. no. 03. [Google Scholar]
- Luman, D.; Joselyn, M.; Suloway, L. Critical Trends Assessment Project: Landcover Database; Illinois Natural History Survey: Champaign, IL, USA, 2009. [Google Scholar]
- Kennedy, P.L.; DeBano, S.J.; Bartuszevige, A.M.; Lueders, A.S. Effects of native and non-native grassland plant communities on breeding passerine birds: Implications for restoration of northwest bunchgrass prairie. Restor. Ecol. 2009, 17, 515–525. [Google Scholar] [CrossRef]
- Triplehorn, C.A.; Johnson, N.F. Borror and Delong’s Introduction to the Study of Insects; Thomson Brooks/Cole: Belmont, CA, USA, 2005. [Google Scholar]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.E.; Hinds, W.; Buschbom, R.L. A general weight vs. Length relationship for insects. Ann. Entomol. Soc. Am. 1976, 69, 387–389. [Google Scholar] [CrossRef]
- Burnham, K.; Anderson, D. Model Selection and Inference: A Practical Informationtheoretic Approach: 60–64; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1998. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. Aic model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Elston, D.; Moss, R.; Boulinier, T.; Arrowsmith, C.; Lambin, X. Analysis of aggregation, a worked example: Numbers of ticks on red grouse chicks. Parasitology 2001, 122, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.R.; Mahoney, M.J.; Cashatt, E.D.; Noordijk, J.; de Snoo, G.; Musters, C. The impact of landscape complexity on invertebrate diversity in edges and fields in an agricultural area. Insects 2016, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P.; Dobkin, D.S.; Wheye, D. Birder’s Handbook; Simon and Schuster: New York, NY, USA, 1988. [Google Scholar]
- Kleen, V.M.; Cordle, L.; Montgomery, R.A. The Illinois Breeding Bird Atlas; Illinois Natural History Survey: Champaign, IL, USA, 2004; Volume 26. [Google Scholar]
- Kang, W.; Hoffmeister, M.; Martin, E.A.; Steffan-Dewenter, I.; Han, D.; Lee, D. Effects of management and structural connectivity on the plant communities of organic vegetable field margins in south korea. Ecol. Res. 2013, 28, 991–1002. [Google Scholar] [CrossRef]
- Noordijk, J.; Musters, C.; van Dijk, J.; de Snoo, G.R. Invertebrates in field margins: Taxonomic group diversity and functional group abundance in relation to age. Biodivers. Conserv. 2010, 19, 3255–3268. [Google Scholar] [CrossRef]
- Scheffer, M.; Achterberg, A.A.; Beltman, B. Distribution of macro-invertebrates in a ditch in relation to the vegetation. Freshw. Biol. 1984, 14, 367–370. [Google Scholar] [CrossRef]
- Healy, W.M. Turkey poult feeding activity, invertebrate abundance, and vegetation structure. J. Wildl. Manag. 1985, 49, 466–472. [Google Scholar] [CrossRef]
- Pryke, J.S.; Samways, M.J. Differential resilience of invertebrates to fire. Austral Ecol. 2012, 37, 460–469. [Google Scholar] [CrossRef]
- De Snoo, G.; Poll, R.v.d.; Bertels, J. Butterflies in sprayed and unsprayed field margins. J. Appl. Entomol. 1998, 122, 157–161. [Google Scholar] [CrossRef]
- Frampton, G.K.; Dorne, J.L.C.M. The effects on terrestrial invertebrates of reducing pesticide inputs in arable crop edges: A meta-analysis. J. Appl. Ecol. 2007, 44, 362–373. [Google Scholar] [CrossRef]
- Muskett, C.; Jones, M. The dispersal of lead, cadmium and nickel from motor vehicles and effects on roadside invertebrate macrofauna. Environ. Pollut. Ser. A Ecol. Biol. 1980, 23, 231–242. [Google Scholar] [CrossRef]
- Forman, R.T. Road ecology: A solution for the giant embracing us. Landsc. Ecol. 1998, 13. [Google Scholar] [CrossRef]
- Noordijk, J.; Schaffers, A.P.; Heijerman, T.; Boer, P.; Gleichman, M.; Sýkora, K.V. Effects of vegetation management by mowing on ground-dwelling arthropods. Ecol. Eng. 2010, 36, 740–750. [Google Scholar] [CrossRef]
- Parr, T.; Way, J. Management of roadside vegetation: The long-term effects of cutting. J. Appl. Ecol. 1988, 1073–1087. [Google Scholar] [CrossRef]
- Morris, M. Resoneses of grassland invertebrates to mangement by cutting: IV positive responses of auchenorhyncha. J. Appl. Ecol. 1981, 18, 763–771. [Google Scholar] [CrossRef]
- Padoa-Schioppa, E.; Baietto, M.; Massa, R.; Bottoni, L. Bird communities as bioindicators: The focal species concept in agricultural landscapes. Ecol. Indic. 2006, 6, 83–93. [Google Scholar] [CrossRef]
- Temple, S.A.; Wiens, J.A. Bird populations and environmental changes: Can birds be bio-indicators. Am. Birds 1989, 43, 260–270. [Google Scholar]
- Furness, R. Birds as monitors of pollutants. In Birds as Monitors of Environmental Change; Springer: Dordrecht, The Netherlands, 1993; pp. 86–143. [Google Scholar]
- Holmes, R.T.; Schultz, J.C.; Nothnagle, P. Bird predation on forest insects: An exclosure experiment. Science 1979, 206, 462–463. [Google Scholar] [CrossRef] [PubMed]
- Hammers, M.; Müskens, G.J.; van Kats, R.J.; Teunissen, W.A.; Kleijn, D. Ecological contrasts drive responses of wintering farmland birds to conservation management. Ecography 2015, 38, 813–821. [Google Scholar] [CrossRef]
- Leopold, A. A Sand County Almanac, 1949; Ballantine: New York, NY, USA, 1970. [Google Scholar]
- Smith, A.C.; Fahrig, L.; Francis, C.M. Landscape size affects the relative importance of habitat amount, habitat fragmentation, and matrix quality on forest birds. Ecography 2011, 34, 103–113. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Variable Increase | TD Edge | TD Field | WT Edge | WT Field |
|---|---|---|---|---|
| Edge Height | Decreased | Increased | Increased | Increased |
| Edge Variation | Increased | Increased | Increased | Increased |
| Edge Length | Decreased | Increased | Decreased | Increased |
| Edge Depth | Increased | Decreased | Increased | Decreased |
| Bare ground % | Increased | Increased | Increased | Increased |
| Management | Mown (least) | No Impact | Mown (least) | No Impact |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, T.R.; Mahoney, M.J.; Cashatt, E.D.; De Snoo, G.; Musters, C.J.M. Enhancement of Linear Agricultural Areas to Provide Invertebrates as Potential Food for Breeding Birds. Land 2016, 5, 26. https://doi.org/10.3390/land5030026
Evans TR, Mahoney MJ, Cashatt ED, De Snoo G, Musters CJM. Enhancement of Linear Agricultural Areas to Provide Invertebrates as Potential Food for Breeding Birds. Land. 2016; 5(3):26. https://doi.org/10.3390/land5030026
Chicago/Turabian StyleEvans, Tracy R., Meredith J. Mahoney, Everett D. Cashatt, Geert De Snoo, and C.J.M. Musters. 2016. "Enhancement of Linear Agricultural Areas to Provide Invertebrates as Potential Food for Breeding Birds" Land 5, no. 3: 26. https://doi.org/10.3390/land5030026
APA StyleEvans, T. R., Mahoney, M. J., Cashatt, E. D., De Snoo, G., & Musters, C. J. M. (2016). Enhancement of Linear Agricultural Areas to Provide Invertebrates as Potential Food for Breeding Birds. Land, 5(3), 26. https://doi.org/10.3390/land5030026