Abstract
This study examines whether there is a biodiversity benefit (“dividend”) associated with the existence and management of conservation reserves in the extensive and largely natural landscape of northern Australia. Species richness and abundance of vertebrate fauna and the intensity of a range of disturbance factors were compared across a set of 967 sampled quadrats, located either in pastoral lands, Indigenous lands or conservation reserves, with all sampled quadrats within a single vegetation type (open forests and savannah woodlands dominated by Eucalyptus miniata and/or E. tetrodonta). The relationships with land tenure varied between major taxonomic groups, but generally (and particularly for threatened species) values were highest for conservation reserves. This “biodiversity dividend” associated with conservation reserves is considered to be due to the effects of management rather than because conservation reserves were established on lands supporting atypically high conservation values. The impact of weeds and (unsurprisingly) livestock was greatest on pastoral lands, and pig impact was greatest in conservation reserves. Although pastoral and Indigenous lands supported lower biodiversity tallies than reserved lands, the conservation values of reserved lands in this region are probably substantially supported by the maintenance of relatively intact ecological systems across all lands.
1. Introduction
Across much of the world, land tenure markedly delineates and influences biodiversity conservation values, typically with residual intact biotic communities most persistent in conservation reserves, whereas lands of contrasting use and tenure have biotic communities of substantially modified composition. In much-transformed landscapes, biodiversity conservation may be largely dependent upon the network of conservation reserves [1]. In landscapes characterised by far less transformation, such as much of the world’s tropical savannahs, other land tenure types may make a contribution to biodiversity conservation that more nearly equals (and/or provides a necessary buffer and complement to) that provided by conservation reserves. Notwithstanding their recognised or assumed significance for biodiversity conservation and some obligation to demonstrate that reserve management investments produce conservation outcomes, there has been relatively little systematic consideration of the effectiveness of reserves compared with lands of other tenures [2,3,4,5,6].
Northern Australia provides an example of an extensive largely intact landscape (<5% cleared or significantly transformed) [7], managed under a range of land tenures, particularly low-intensity pastoralism (comprising 75% of the total land area), Indigenous lands (15%) and conservation reserves (6%), all characterised by general retention of native vegetation [8]. Unsurprisingly, given the relatively minor levels of modification of their lands, some landholder groups have asserted that their management provides good conservation outcomes and, hence, that there is little need for further reservation. Indeed, the maintenance of biodiversity is an explicit condition of the regulation of the pastoral industry in the Northern Territory, Australia. With respect to Indigenous lands, it has been asserted that traditional management practices are likely to be particularly effective for the maintenance of biodiversity [9]. This latter claim has fuelled substantial development of policies and practice that equate biodiversity conservation with Indigenous land management and its resourcing [10,11,12] and the recent development of Indigenous Protected Areas as significant components of Australia’s national reserve system [13,14]. However, the most substantial supportive study [9] may be an insecure basis for such a claim, given that it focused on one relatively small area and lacked controlled comparison with lands subjected to contrasting management.
Here, we undertake a systematic and very substantial assessment of vertebrate biodiversity in different land tenure types across an extensive area (ca. 300,000 km2) of the monsoonal tropics of the Northern Territory. To focus the assessment particularly on land tenure and its related management and reduce extraneous environmental variability, we restrict the survey and analysis to one vegetation type, forests dominated by Eucalyptus miniata and/or E. tetrodonta. This forest type is the characteristic vegetation across much of northern Australia, with a total area of about 450,000 km2 [7]. Our objective is to compare, between different land tenure types, the biodiversity (and also the intensity of possible threatening factors) across lands supporting this vegetation type.
The land tenure types we consider are pastoral lands, Indigenous lands and conservation reserves. We recognise that in this region, as elsewhere, there may not be a close match between land tenure and the intensity and form of management. However, in this region, conservation reserves are typically managed to reduce the incidence of feral animals (principally pigs Sus scrofa, cattle Bos indicus and water buffalo Bubalus bubalis) and weeds and to manage fire regimes in a manner that aims to reduce the incidence of high intensity uncontrolled late dry season fire [15]. Pastoral lands are managed with extensive (typical property sizes 1,000 to 10,000 km2), but typically low-intensity (stocking rates typically <20 cattle/km2) ranching for beef cattle, with occasional areas of more intensified manipulation (some clearing and planting of exotic pasture grasses, but little or no additions of nutrients). The management of Indigenous lands varies along a gradient from traditional intensive manipulation of fire to absence of any management (largely due to depopulation or abandonment). Note that in this region, some conservation reserves are Aboriginal lands now jointly managed; these were treated here as conservation reserves.
A related broad-based assessment of land tenure impacts upon biodiversity and environmental management in this region was undertaken recently [16]. The current study differs from that work in that our study is (purposefully) restricted to one vegetation type, provides a direct survey-based assessment of faunal biodiversity and relates disturbance measures and biodiversity assessed together for a large set of individual quadrats.
We note that two other study designs may provide more incisive assessments of the efficacy of conservation reserves—matched cross-fence comparisons of contrasting land tenure (or management) types (e.g., [17]) and controlled time-series studies that seek to measure divergence in conservation values between a recently-established conservation reserve and a nearby unreserved area. Using the first approach, recent studies have demonstrated significant differences between pastoral lands and ungrazed lands in the richness and composition of both their vertebrate and invertebrate faunas in forests of tropical north-eastern Australia [18,19], greater abundance of native mammals in conservation reserves compared with paired sites on pastoral lands in tropical north-eastern Australia [20] and significant divergence in vegetation and vertebrate faunas across adjacent sites of markedly contrasting fire history [21]. In northern Australia, two recent studies have chronicled change over time in fauna assemblages following de-stocking (change from pastoral areas to conservation reserves), although the magnitude of the resulting increased abundance in native mammals varied appreciably between these two studies [22,23]. Furthermore, some monitoring studies have demonstrated responses over time of vegetation and vertebrates in relationship to management inputs on conservation lands [24,25,26,27].
In contextualising this study, we note that our focus is solely on vertebrate fauna and that patterns may differ for other components of biodiversity. For example, there are now well-established demonstrations that some fire-sensitive plant species may prosper on lands subjected to some forms of traditional Aboriginal fire management, but decline in lands subjected to less intensive fire management [28,29,30,31,32,33,34,35,36,37]. We recognise the limitation in our ambit, but consider it a necessary counterpoint to the bulk of those previous related studies that have asserted the benefits to biodiversity of Indigenous management mainly using evidence from a restricted (but nonetheless important) set of fire-sensitive plant species. Furthermore, we consider that the inclusion here of birds, mammals, reptiles and frogs (totalling >400 species) is likely to provide a relatively broad mix of ecologies and environmental responses to land tenure and management and, hence, a reasonably robust baseline for ecological generalizations.
2. Methods
2.1. Sampling
A consistent protocol for the survey of vertebrate fauna has been established in the Northern Territory over the period 1988 to the present [38], during which more than 5,000 quadrats have been sampled. Over the course of this systematic sampling, the selection of locations of quadrats was generally related to providing regional inventory, rather than the consideration on which the present study focuses.
Briefly, the sampling provides a measure of the abundance of frog, reptile, bird and mammal species over a 72 h period for a marked 50 m × 50 m quadrat (for mammals, reptiles and frogs), nestled within a 1 ha quadrat (for birds). For each individual species, the abundance measure was derived from live-trapping results (for frogs, reptiles and mammals; using 20 Elliott traps, four cage traps and two pitfall buckets) and timed searches (for birds—eight instantaneous counts; and for other taxa—three 10 min day-time searches and two 10 min spotlight searches at night).
A set of environmental variables was recorded for every quadrat, including basal area for all woody plant species, rock cover and a five-class scale (from 0 (no apparent impact) to 5 (severe impact)) for each of a range of disturbance variables (including weeds, pigs, fire and livestock or feral stock): see [39] for more details of scoring for these disturbance values. The average annual rainfall of every quadrat was estimated from modelled surfaces in ANUCLIM [40].
From the large collated data base of sampled quadrats, we selected all forest or woodland quadrats for which E. miniata and/or E. tetrodonta contributed at least 20% of the total woody basal area and which had not been grossly disturbed by clearing. This resulted in a selected set of 967 quadrats, widely spaced across the range of this forest type in the Northern Territory (Figure 1). The land tenure of each quadrat, at the time of sampling, was then identified by reference to cadastral mapping and information on land use and type recorded when sampled. The land tenure tallies of samples comprised 386 quadrats on Aboriginal lands, 487 on conservation reserves (including 14 different conservation reserves) and 94 on pastoral lands. Some (285) of these quadrats were on islands and, as such, location alone may influence biodiversity composition and conservation value [41]; we repeated all analyses with these island quadrats excluded.
Over the geographic and environmental range occupied by forests dominated by E. miniata and/or E. tetrodonta, fauna assemblages have previously been shown to vary with annual rainfall and rock cover [42,43,44], and so these variables were explicitly considered within analyses (see below). In this region, fauna assemblages may also vary with soil texture [44], but forests of E. miniata and/or E. tetrodonta grow on only a narrow range of textures (typically sandy-loams), so this variable was not considered in analysis. Given that our sampling spanned the period 1988-2008 and that this period coincided with a regional-scale decline in native mammals [45], we also included year within modelling.
For each quadrat, we tallied the species richness and total abundance of native frogs, reptiles, birds and mammals and the richness and total abundance of threatened species (as listed in [46]). Because of difficulties in consistent sampling, we do not include bats within our mammal tallies.
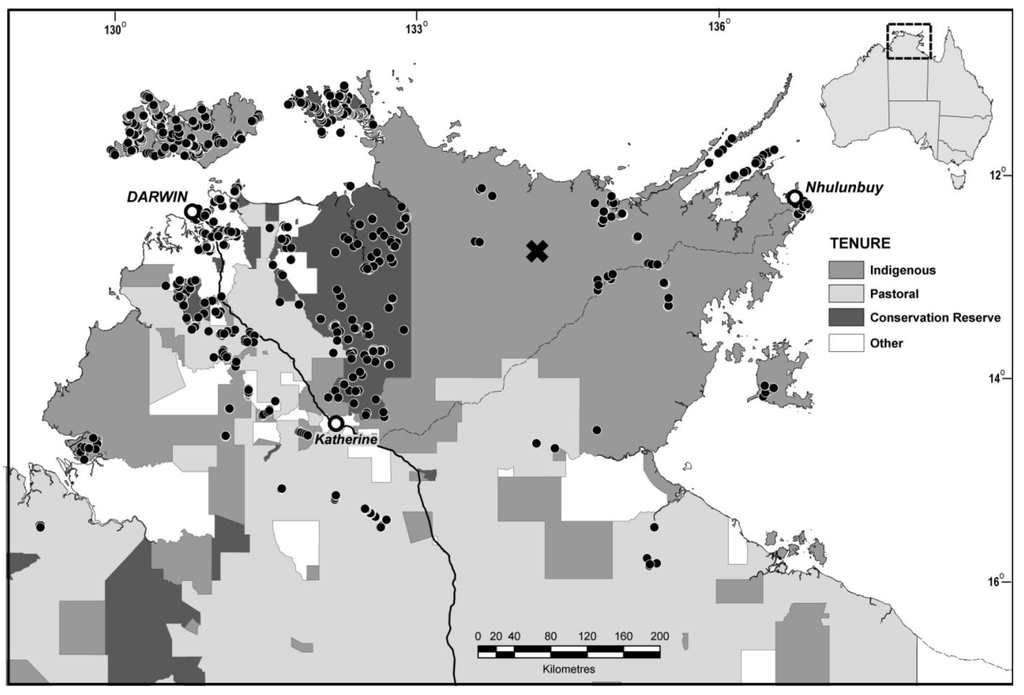
Figure 1.
Location of study area, land tenure and sampled quadrats. The location of the Cadell River study of [9] is indicated with a cross.
2.2. Analysis
We estimated the relationship between land tenure and biodiversity using generalised linear models [47], where the species richness is modelled with a Poisson distribution. The count of species (y) observed was related to the covariates (x = land tenure, rainfall, rockiness, year), and we used this model to estimate coefficients (β- measures of effect) for n covariates:
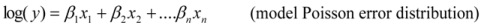
We conducted this Poisson regression in log space to limit the estimates to positive values.
We conducted model averaging to parsimoniously estimate the effect of land tenure and other covariates (rock cover, rainfall and year), from the set of candidate models. Model averaging combines the coefficient estimates (β) according to their Akaike weight:
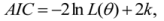
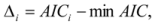
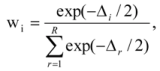
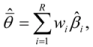 where L is the likelihood of the model, k is the number of parameters, Δi is the change in Akaike Information Criteria, wi is the Akaike weight for the model i and
where L is the likelihood of the model, k is the number of parameters, Δi is the change in Akaike Information Criteria, wi is the Akaike weight for the model i and 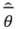 is the model averaged estimate for a set of R models [48]. We conducted the Poisson regression, model averaging and predictions (except those previously mentioned) in program R [49].
is the model averaged estimate for a set of R models [48]. We conducted the Poisson regression, model averaging and predictions (except those previously mentioned) in program R [49].
is the model averaged estimate for a set of R models [48]. We conducted the Poisson regression, model averaging and predictions (except those previously mentioned) in program R [49].We compared the level of disturbance recorded in quadrats of contrasting land tenure, using Kruskal-Wallis ANOVA, for impacts of fire, weeds, feral pigs and livestock (including feral and non-feral buffalo and cattle).
3. Results: Impacts of Land Tenure and Other Variables
3.1. Fauna
For most of the fauna parameters considered, the model incorporating all four terms (rock cover, rainfall, land tenure and year) was overwhelmingly the best model (Table 1). There were two exceptions. For reptile species richness, no single model was particularly good, but the best model included only the terms rock cover and land tenure. For richness of threatened species, the best model contained the terms rainfall, land tenure and year. The direction of relationships with rainfall, rock cover and year varied between vertebrate groupings, and these relationships are not considered further here. For all but two vertebrate groupings, the land tenure relationship was consistent, with the highest value in conservation reserves. The exceptions were for mammal richness and amphibian richness, both of which were marginally higher in pastoral lands than in conservation reserves (Figure 2).
The exclusion of island quadrats did not change the terms included in any models or the direction of relationship or provide any substantial change in the explanatory power, and accordingly, the models excluding island quadrats are not presented here.

Table 1.
Summary of model performance for vertebrate species richness and abundance. “Full” model includes all of the terms: rock cover, land tenure, year and rainfall.
| Fauna Group Attribute | Model | Deviance | AICc | Delta | Weight |
|---|---|---|---|---|---|
| Vertebrate richness | full | 2,616.0 | 6,982.4 | 0 | 0.98 |
| rock, tenure, year | 2,626.3 | 6,990.7 | 8.3 | 0.02 | |
| Vertebrate abundance | full | 2,0531 | 2,594.8 | 0 | 1 |
| rock, rainfall, tenure | 2,0619 | 2,603.5 | 86.2 | 0 | |
| Amphibian richness | full | 1,238.7 | 1,952.1 | 0 | 0.79 |
| rainfall, tenure, year | 1,243.5 | 1,954.8 | 2.7 | 0.21 | |
| Amphibian abundance | full | 4,887.5 | 5,796.7 | 0 | 1 |
| rainfall, tenure, year | 4,930.6 | 5,837.8 | 41.1 | 0 | |
| Reptile richness | rock, tenure | 1,056.5 | 4,039.0 | 0 | 0.23 |
| tenure | 1,059.7 | 4,040.1 | 1.12 | 0.13 | |
| rock, tenure, year | 1,055.8 | 4,040.2 | 1.24 | 0.12 | |
| rainfall, rock, tenure | 1,056.5 | 4,040.9 | 1.96 | 0.08 | |
| rock | 1,063.0 | 4,041.4 | 2.40 | 0.07 | |
| Reptile abundance | full | 5,785.4 | 9,525.2 | 0 | 0.74 |
| rainfall, tenure, year | 5,789.4 | 9,527.3 | 2.1 | 0.26 | |
| Bird richness | full | 2,876.7 | 6,877.5 | 0 | 1 |
| rainfall, rock, year | 2,913.4 | 6,910.1 | 32.6 | 0 | |
| Bird abundance | full | 23,250.9 | 28,366.0 | 0 | 1 |
| rainfall, rock, tenure | 23,320.5 | 28,433.3 | 67.4 | 0 | |
| Mammal richness | full | 1,248.2 | 3,091.4 | 0 | 0.99 |
| rainfall, tenure, year | 1,260.3 | 3,101.6 | 10.1 | 0.01 | |
| Mammal abundance | full | 5,387.1 | 7,677.3 | 0 | 1 |
| rainfall, tenure, year | 5,453.1 | 7,741.2 | 63.9 | 0 | |
| Threatened species richness | rainfall, tenure, year | 760.8 | 1,433.9 | 0 | 0.70 |
| full | 760.6 | 1,435.7 | 1.79 | 0.29 | |
| Threatened species abundance | full | 2,120.9 | 2,953.7 | 0 | 0.66 |
| rainfall, tenure, year | 2,124.2 | 2,955.0 | 1.35 | 0.34 |
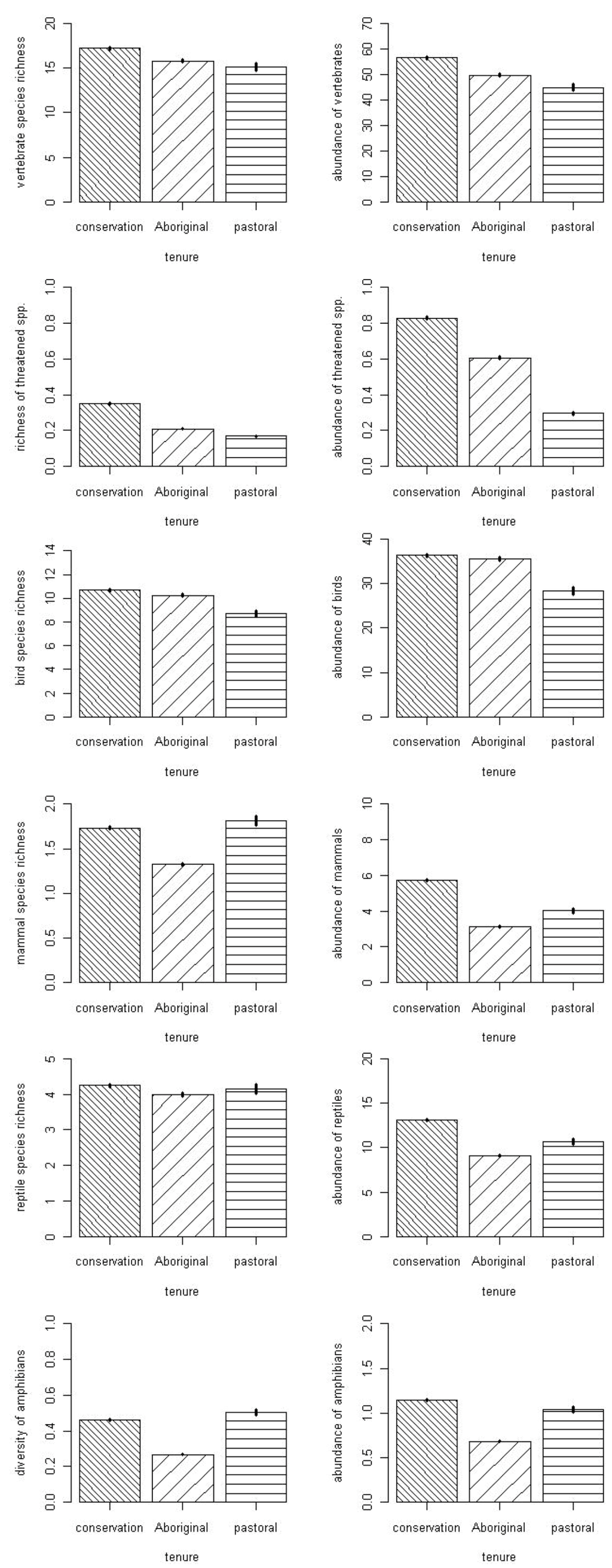
Figure 2.
Variation amongst land tenures (conservation reserves, Indigenous (Aboriginal) lands and pastoral lands) in the richness and abundance of vertebrate groups. Bars show means and whiskers standard errors, based on model averaging with all covariates included.
3.2. Threats
Impact scores for weeds, livestock and pigs, but not fire, varied significantly between land tenure types (Table 2). Weed impact was greatest on pastoral lands and least on Aboriginal lands. Unsurprisingly, livestock (cattle) impact was also greatest on pastoral lands. Pig impact was greatest on conservation lands.
Table 2.
Summary table of environmental disturbance variables and their relationship to land tenure. Values in body of table are means (and relate to a scale from 0 (no impact) to 5 (severe impact)). Significance levels: * p < 0.05; ** p < 0.01; *** p < 0.001.
| Variable | Conservation Reserves | Pastoral | Aboriginal Lands | H |
|---|---|---|---|---|
| Fire impact | 1.75 | 1.64 | 1.58 | 5.9 ns |
| Weed impact | 0.15 | 0.85 | 0.02 | 120.2 *** |
| Cattle impact | 0.41 | 0.70 | 0.41 | 10.6 ** |
| Pig impact | 0.42 | 0.19 | 0.14 | 41.7 *** |
4. Interpretation and Discussion
Across a very large data set of quadrats sampled consistently for vertebrate fauna and a single (extensive) vegetation type, we attempted to distil the influence of land tenure (and its partly matched associate of land management) on biodiversity. Although the resulting models indicate that species richness and abundance is associated also with some other environmental factors, the results demonstrate that there is a significant influence of land tenure on vertebrate species richness and abundance. Of the three land tenures (pastoral, conservation reserves and Indigenous lands), conservation reserves supported highest species richness and total abundance for most fauna groups considered. There was some variation amongst vertebrate groups in the magnitude of this response to land tenure, with the most marked response being the markedly reduced abundance of threatened species on pastoral lands. Our study does not provide a definitive explanation of this particular response, but we speculate that it may be because many now threatened species have an association with relatively dense ground-layer cover, which would be most depleted (by grazing) on pastoral lands.
The trend suggests that management on conservation reserves may be having some beneficial impacts upon (at least vertebrate) biodiversity relative to other land types. We note that the findings of this study relate to vertebrate fauna only and may not apply comparably to plants and invertebrates; however, some previous studies in the region have indicated some broad commonality between vertebrate and invertebrate faunas in responses to land use [18,19], and we note also that sampling across birds, mammals, reptiles and frogs provides a reasonably broad scope for ecological response.
This “biodiversity dividend” associated with conservation reserves appears to be small, but significant. In part, the limited effect of conservation reserve tenure may be due to the relatively recent (mostly post-1960s) establishment of many reserves, typically from areas formerly exposed to a substantial history of pastoralism. In part, the limited effect of conservation reserve tenure may be due also to the limited resourcing for management that characterizes most conservation reserves in this region and, consequently, the limited extent to which detrimental disturbance factors are being controlled in conservation reserves [50]. In part, the limited dividend may also be because, in this region, the non-reserve land tenures are also relatively benign for biodiversity (although the results indicate that this assumption may be invalid for threatened species). Note that our use of the term “biodiversity dividend” should not imply that there has been a bonus above and beyond background rates of biodiversity. Indeed, given some broad-scale declines of biodiversity in this region across all land tenures [45], the biodiversity dividend for conservation reserves may be better interpreted as a biodiversity loss that is relatively less than the losses on other land tenures.
It may be argued that the higher values reported here for vertebrate biodiversity on conservation reserves than on other extensive land tenures may be due to deliberate establishment of those reserves on areas known to have supported high biodiversity value: that is, that the high biodiversity value preceded the conservation management. We suggest that this is not the case, because the location of conservation reserves in this region has been driven mostly by opportunity, by scenic value or by recreational potential [51].
The results also demonstrate a contrasting constellation of disturbance regimes operating across these different land tenures, with such differences in the incidence or impacts of threats presumably contributing to the observed differences in vertebrate species richness and abundance. The differences between land tenures in disturbance regimes observed in this study are consistent with some previous observations in this region, including a study restricted to the small rainforest patches embedded within the extensive savannah woodlands [52]. Both studies reported that weed impacts were highest on pastoral lands, then conservation reserves, then Indigenous lands; and both studies reported that pig impacts were greatest in conservation reserves, then pastoral lands, then Indigenous lands. However, the previous rainforest study reported that fire impacts were most severe for rainforests on pastoral lands, whereas no such trend was evident in our data [52]. Two other studies also reported an incidence of weed impacts that was higher on pastoral lands than in conservation reserves or Indigenous lands, with the relatively low incidence of weeds on Indigenous lands, most likely to be due to remoteness from major development rather than a consequence of intrinsic management [16,53].
This study provides no substantial support for previous assertions (notably [9]) of the superior value of Indigenous land management for fostering biodiversity. The broad regional analysis reported here indicates that, when corrected for variation in rainfall and rockiness, quadrats on Indigenous lands typically supported fewer frogs, reptiles, birds and mammals than comparably-sampled quadrats in the same environment on conservation reserves. However, we note that the management of Indigenous lands considered in this study varied substantially across a gradient from lands where traditional intensive management was being continued to lands that had been largely depopulated and neglected. Hence, our results here reflect land tenure per se rather than management.
Across northern Australia, there is now substantial investment by governments and a substantial policy framework for supporting environmental management actions by Indigenous people on Indigenous lands. This support is clearly providing important socio-economic contributions to disadvantaged communities, and there is evidence that involvement in management is producing health benefits [54,55]. However, while there may be some reasonable supposition that such management may produce some biodiversity benefit, particularly for some fire-sensitive plant species [9,13,14,56], there is as yet no compelling evidence to support such happy conflation more generally. The results reported here suggest that a far more rigorous evidence-based assessment of the environmental outcomes of this investment is required [57]. In considering such supporting evidence, we note the bluntness of the data we report here (one-off inventory, of species richness and total abundance) and recognise that a more sensitive and targeted measurement (and comparison) of management efficacy would be provided by monitoring of biodiversity trends comparably across lands of different tenure and exposed to contrasting management objectives, methods and resource investment.
We also recognise that there is increasing blending of Indigenous land management skills and conventional park management (the “two tool-box” approach [14,58]) on conservation reserves and Indigenous lands across much of northern Australia and increasing resources available for conservation management, particularly on Indigenous lands [14]. Such a recent development is likely to provide some increases in biodiversity outcomes, particularly for lands that have until recently been largely depopulated and unmanaged [30].
For pastoral lands, our results demonstrate that biodiversity values are being reasonably well maintained, albeit with values generally less than for conservation reserves. However, this is not the case for the set of threatened species, where the values for pastoral land were substantially (>50%) less than for conservation reserves. This contrast reinforces the point that conservation value may be poorly measured by simply the richness and abundance of broad taxonomic groups. The result here for threatened species suggests that pastoral lands may provide relatively little conservation benefit for these species and, indeed, that pastoralism across most of these north Australian landscapes may have contributed to the currently threatened status of some species. This conclusion, from broad-scale comparison, complements some findings of highly localised (typically cross-fence contrasts) that have demonstrated localised detrimental impacts of pastoralism on biodiversity [18,20,22].
This study provides correlative evidence of a response to land tenure and a foundation for subsequent research. Such research may provide stronger and more direct evidence if it involves ongoing monitoring of biodiversity at sites where land tenure (and/or management) has changed [22] and/or more intensive studies of habitat quality and ecology for species for which abundance varies between land tenures.
5. Conclusions
These results demonstrate a biodiversity dividend, albeit relatively small, that is attributable to the existence of conservation reserves, even within extensive largely intact landscapes. This conclusion, from our extensive regional-scale sampling, is broadly consistent with results from the few other studies (whose sampling has mostly involved other approaches (notably cross-fence comparisons) at smaller geographic scales) [18,19,20,21].
In this region, the continued existence of natural landscapes across the far more extensive matrix in which these reserves are embedded may be a major reason why these reserves maintain their biodiversity values [59]. Although less than for conservation reserves, Indigenous lands and pastoral lands are here shown to have substantial biodiversity value, notwithstanding their contrasting management regimes, intensity and objectives. Given the relatively small proportion of lands currently devoted to conservation reserves in this area, these unreserved lands make a very substantial contribution to regional biodiversity conservation. This conclusion suggests that there is substantial justification for investing in biodiversity management for lands of all tenures and managing this ecologically continuous land for biodiversity outcomes without excessive concern for cadastral boundaries. This is a markedly different situation to many other parts of the world, where broad-scale land-use impacts outside reserves result in a more marked contrast in biodiversity values between reserved and unreserved lands and probably ultimately lead to erosion of park values through fragmentation effects [60,61,62].
In this extensive natural landscape, lands of all tenures may contribute substantially to biodiversity conservation, but conservation reserves make the most substantial contribution. However, even in the largest and best-resourced conservation reserve in this region (Kakadu National Park), some components of biodiversity are currently undergoing considerable decline, suggesting that the management of threats even in conservation reserves is insufficient to meet the conservation need; that the biodiversity dividend reported here for conservation reserves may instead be simply a biodiversity detriment that happens to be less than on other land tenures [27,45].
Acknowledgments
This study was funded in part by an Australia Research Council Linkage Grant (LP0455163). The data considered here derive from many years of fauna surveys undertaken by the Northern Territory Department of Natural Resources Environment The Arts and Sport, with many survey staff, but with particular mention of Martin Armstrong, Kym Brennan and Damian Milne. We thank Simon Davies, Kamil Barton and the R core team for statistical R functions. We thank Graham Phelps, Stephen Garnett, Peter Whitehead and three anonymous referees for their helpful comments. We thank very many landholders for permission to work on their lands.
References
- Jackson, S.F.; Gaston, K.J. Land use change and the dependence of national priority species on protected areas. Glob. Change Biol. 2008, 14, 2132–2138. [Google Scholar] [CrossRef]
- Parrish, J.D.; Braun, D.P.; Unnasch, R.S. Are we conserving what we say we are? Measuring ecological integrity within protected areas. BioScience 2003, 53, 851–860. [Google Scholar] [CrossRef]
- Chape, S.; Harrison, J.; Spalding, M.; Lysenko, I. Measuring the extent and effectiveness of protected areas as an indicator for meeting global biodiversity targets. Phil. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 443–455. [Google Scholar] [CrossRef]
- Devictor, V.; Godet, L.; Julliard, R.; Couvet, D.; Jiguet, F. Can common species benefit from protected areas? Biol. Conserv. 2007, 139, 29–36. [Google Scholar] [CrossRef]
- Gaston, K.J.; Charman, K.; Jackson, S.F.; Armsworth, P.R.; Bonn, A.; Briers, R.A.; Callaghan, C.S.Q.; Catchpole, R.; Hopkins, J.; Kunin, W.E.; et al. The ecological effectiveness of protected areas: the United Kingdom. Biol. Conserv. 2006, 132, 76–87. [Google Scholar] [CrossRef]
- Rannestad, O.T.; Danielsen, T.; Moe, S.R.; Stokke, S. Adjacent pastoral areas support higher densities of wild ungulates during the wet season than the Lake Mburo National Park in Uganda. J. Trop. Ecol. 2006, 22, 675–683. [Google Scholar] [CrossRef]
- Woinarski, J.; Mackey, B.; Nix, H.; Traill, B. The Nature of Northern Australia: Natural Values, Ecological Processes and Future Prospects; ANU E-Press: Canberra, ACT, Australia, 2007. [Google Scholar]
- Garnett, S.T.; Woinarski, J.C.Z.; Crowley, G.M.; Kutt, A.S. Biodiversity conservation in Australian tropical rangelands. In Wild Rangelands: Conserving Wildlife While Maintaining Livestock in Semi-Arid Ecosystems; du Toit, J., Kock, R., Deutsch, J., Eds.; Wiley-Blackwell: Chichester, UK, 2010; pp. 191–234. [Google Scholar]
- Yibarbuk, D.; Whitehead, P.J.; Russell-Smith, J.; Jackson, D.; Godjuwa, C.; Fisher, A.; Cooke, P.; Choquenot, D.; Bowman, D.M.J.S. Fire ecology and Aboriginal land management in central Arnhem Land, northern Australia: a tradition of ecosystem management. J. Biogeogr. 2001, 28, 325–343. [Google Scholar]
- Altman, J.C.; Whitehead, P.J. Caring for Country and Sustainable Indigenous Development: Opportunities, Constraints and Innovation; Australian National University Centre for Aboriginal Economic Policy Research Working Paper No. 20/2003; ANU: Canberra, ACT, Australia, 2003. [Google Scholar]
- Luckert, M.K.; Campbell, B.M.; Gorman, J.T.; Garnett, S.T. Investing in Indigenous Natural Resource Management; Charles Darwin University: Darwin, NT, Australia, 2007. [Google Scholar]
- Whitehead, P.J.; Bowman, D.M.J.S.; Preece, N.; Fraser, F.; Cooke, P. Customary use of fire by Indigenous peoples in northern Australia: its contemporary role in savannah management. Int. J. Wildland Fire 2003, 12, 415–425. [Google Scholar] [CrossRef]
- Altman, J.C.; Buchanan, G.J.; Larsen, L. The Environmental Significance of the Indigenous Estate: Natural Resource Management as Economic Development in Remote Australia; CAEPR Discussion Paper 286/2007; ANU College of Arts & Social Sciences: Canberra, ACT, Australia, 2007. [Google Scholar]
- Gilligan, B. The Indigenous Protected Area Programme; Department of the Environment and Heritage: Canberra, ACT, Australia, 2006. [Google Scholar]
- Director of National Parks. In Kakadu National Park. Management Plan 2007–2014; Australian Government Director of National Parks: Canberra, ACT, Australia, 2007.
- Franklin, D.C.; Petty, A.M.; Williamson, G.J.; Brook, B.W.; Bowman, D.M.J.S. Monitoring contrasting land management in the savannah landscapes of northern Australia. Environ. Manage. 2008, 41, 501–515. [Google Scholar] [CrossRef]
- Caro, T.M. Species richness and abundance of small mammals inside and outside and African national park. Biol. Conserv. 2001, 98, 251–257. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Ash, A.J. Responses of vertebrates to pastoralism, military land use and landscape position in an Australian tropical savannah. Austral. Ecol. 2002, 27, 311–323. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Andersen, A.N.; Churchill, T.; Ash, A.J. Response of ant and terrestrial spider assemblages to pastoral and military land use and to landscape position, in a tropical savannah woodland in northern Australia. Austral. Ecol. 2002, 27, 324–333. [Google Scholar] [CrossRef]
- Kutt, A.S.; Gordon, I.J. Variation in terrestrial mammal abundance on pastoral and conservation land tenures in north-eastern Australian tropical savannas. Anim. Conserv. 2012, 15, 416–425. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Risler, J.; Kean, L. The response of vegetation and vertebrate fauna to 23 years of fire exclusion in a tropical Eucalyptus open forest, Northern Territory, Australia. Austral. Ecol. 2004, 29, 156–176. [Google Scholar] [CrossRef]
- Legge, S.; Kennedy, M.S.; Lloyd, R.; Murphy, S.A.; Fisher, A. Rapid recovery of mammal fauna in the central Kimberley, northern Australia, following the removal of introduced herbivores. Austral. Ecol. 2011, 36, 791–799. [Google Scholar] [CrossRef]
- Kutt, A.S.; Vanderduys, E.P.; Perry, J.J.; Perkins, G.C.; Kemp, J.E.; Bateman, B.L.; Kanowski, J.; Jensen, R. Signals of change in tropical savannah woodland vertebrate fauna 5 years after cessation of livestock grazing. Wildlife Res. 2012, 39, 386–396. [Google Scholar]
- Edwards, A.; Kennett, R.; Price, O.; Russell-Smith, J.; Spiers, G.; Woinarski, J. Monitoring the impacts of fire regimes on biodiversity in northern Australia: an example from Kakadu National Park. Int. J. Wildland Fire 2003, 12, 427–440. [Google Scholar]
- Russell-Smith, J.; Edwards, A.C.; Woinarski, J.C.Z.; McCartney, J.; Kerin, S.; Winderlich, S.; Murphy, B.P.; Watt, F. The First Ten Years of the ‘Three Parks’ (Kakadu, Litchfield, Nitmiluk) Fire Regime and Biodiversity Monitoring Program. In Culture, Ecology and Economy of Fire Management in Northern Australia: Rekindling the Wurrk Tradition; Russell-Smith, J., Whitehead, P.J., Cooke, P., Eds.; CSIRO Publications: Melbourne, VIC, Australia, 2009; pp. 257–286. [Google Scholar]
- Woinarski, J.C.Z.; Russell-Smith, J.; Andersen, A.; Brennan, K. Fire management and biodiversity of the western Arnhem Land plateau. In Culture, Ecology and Economy of Fire Management in Northern Australia: Rekindling the Wurrk Tradition; Russell-Smith, J., Whitehead, P.J., Cooke, P., Eds.; CSIRO Publications: Melbourne, VIC, Australia, 2009; pp. 201–228. [Google Scholar]
- Woinarski, J.C.Z.; Armstrong, M.; Brennan, K.; Fisher, A.; Griffiths, A.D.; Hill, B.; Milne, D.J.; Palmer, C.; Ward, S.; Watson, M.; et al. Monitoring indicates rapid and severe decline of native small mammals in Kakadu National Park, northern Australia. Wildlife Res. 2010, 37, 116–126. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Panton, W.J. Decline of Callitris intratropica R.T. Baker & H.G. Smith in the Northern Territory: implications for pre- and post-European colonization ire regimes. J. Biogeogr. 1993, 20, 373–381. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Prior, L.D. Impact of Aboriginal landscape burning on woody vegetation in Eucalyptus tetrodonta savannah in Arnhem Land, northern Australia. J. Biogeogr. 2004, 31, 807–817. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Price, O.; Whitehead, P.J.; Walsh, A. The ‘wilderness effect’ and the decline of Callitris intratropica on the Arnhem Land Plateau, northern Australia. Aust. J. Bot. 2001, 49, 665–672. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Walsh, A.; Prior, L.D. Landscape analysis of Aboriginal fire management in central Arnhem Land, north Australia. J. Biogeogr. 2004, 31, 207–223. [Google Scholar] [CrossRef]
- Price, O.; Bowman, D.M.J.S. Fire-stick forestry: a matrix model in support of skilful fire management of Callitris intratropica R.T. Baker by north Australian Aborigines. J. Biogeogr. 1994, 21, 573–580. [Google Scholar] [CrossRef]
- Prior, L.D.; Bowman, D.M.J.S.; Brook, B.W. Growth and survival of two north Australian relictual tree species, Allosyncarpia ternata (Myrtaceae) and Callitris intratropica (Cupressaceae). Ecol. Res. 2007, 22, 228–236. [Google Scholar] [CrossRef]
- Russell-Smith, J. Recruitment dynamics of the long-lived obligate seeders Callitris intratropica (Cupressaceae) and Petraeomyrtus punicea (Myrtaceae). Aust. J. Bot. 2006, 54, 479–485. [Google Scholar] [CrossRef]
- Russell-Smith, J.; Lucas, D.; Gapindi, M.; Gunbunuka, B.; Kapirigi, N.; Namingum, G.; Lucas, K.; Giuliani, P.; Chaloupka, G. Aboriginal resource utilization and fire management practice in western Arnhem Land, monsoonal northern Australia: Notes for prehistory, lessons for the future. Hum. Ecol. 1997, 25, 159–195. [Google Scholar] [CrossRef]
- Russell-Smith, J.; Ryan, P.G.; Klessa, D.; Waight, G.; Harwood, R. Fire regimes, fire-sensitive vegetation and fire management of the sandstone Arnhem Plateau, monsoonal northern Australia. J. Appl. Ecol. 1998, 35, 829–846. [Google Scholar]
- Russell-Smith, J.; Ryan, P.G.; Cheal, D.C. Fire regimes and the conservation of sandstone heath in monsoonal northern Australia: frequency, interval, patchiness. Biol. Conserv. 2002, 104, 91–106. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Armstrong, M.; Price, O.; McCartney, J.; Griffiths, T.; Fisher, A. The terrestrial vertebrate fauna of Litchfield National Park, Northern Territory: monitoring over a 6-year period and response to fire history. Wildlife Res. 2004, 31, 1–10. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Rankmore, B.; Hill, B.; Griffiths, A.D.; Stewart, A.; Grace, B. Fauna assemblages in regrowth vegetation in tropical open forests of the Northern Territory, Australia. Wildlife Res. 2010, 36, 675–690. [Google Scholar]
- Houlder, D.J. ANUCLIM (Version 5.1); Centre for Resource and Environmental Studies, Australian National University: Canberra, ACT, Australia, 2000. [Google Scholar]
- Firth, R.S.C.; Woinarski, J.C.Z.; Brennan, K.G.; Hempel, C. Environmental relationships of the brush-tailed rabbit-rat Conilurus penicillatus and other small mammals on the Tiwi Islands, northern Australia. J. Biogeogr. 2006, 33, 1820–1837. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Gambold, N. Gradient analysis of a tropical herpetofauna: distribution patterns of terrestrial reptiles and amphibians in Stage III of Kakadu National Park, Australia. Wildlife Res. 1992, 19, 105–127. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Braithwaite, R.W.; Menkhorst, K.A.; Griffin, S.; Fisher, A.; Preece, N. Gradient analysis of the distribution of mammals in Stage III of Kakadu National Park, with a review of the distribution patterns of mammals across north-western Australia. Wildlife Res. 1992, 19, 233–262. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Fisher, A.; Milne, D. Distribution patterns of vertebrates in relation to an extensive rainfall gradient and soil variation in the tropical savannas of the Northern Territory, Australia. J. Trop. Ecol. 1999, 15, 381–398. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Legge, S.; Fitzsimons, J.A.; Traill, B.J.; Burbidge, A.A.; Fisher, A.; Firth, R.S.C.; Gordon, I.J.; Griffiths, A.D.; Johnson, C.N.; et al. The disappearing mammal fauna of northern Australia: context, cause and response. Conserv. Lett. 2011, 4, 192–201. [Google Scholar] [CrossRef]
- Woinarski, J.; Pavey, C.; Kerrigan, R.; Cowie, I.; Ward, S. Lost from Our Landscape: Threatened Species of the Northern Territory; NT Government Printer: Darwin, NT, Australia, 2007. [Google Scholar]
- Dobson, A.J. An Introduction to Generalized Linear Models; Chapman and Hall: London, UK, 1990. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Development Core Team, R: A Language and Environment for Statistical Computing, Reference Index Version 2.9.2; R Foundation for Statistical Computing: Vienna, Austria, 2009.
- Parr, C.L.; Woinarski, J.C.Z.; Pienaar, D.J. Cornerstones of biodiversity conservation? Comparing the management effectiveness of Kruger and Kakadu National Parks, two key savannah reserves. Biodivers. Conserv. 2009, 18, 3643–3662. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z. A Difficult and Destructive Metamorphosis: Conservation and Land Management in the Northern Territory in the 1950s. In Modern Frontier: Aspects of the 1950s in Australia’s Northern Territory; Wells, J.T., Dewar, M., Parry, S., Eds.; Charles Darwin University Press: Darwin, NT, Australia, 2005; pp. 33–55. [Google Scholar]
- Russell-Smith, J.; Bowman, D.M.J.S. Conservation of monsoon rainforest isolates in the Northern Territory, Australia. Biol. Conserv. 1992, 59, 51–63. [Google Scholar] [CrossRef]
- Preece, N.; Harvey, K.; Hempel, C.; Woinarski, J.C.Z. Uneven distribution of weeds along extensive transects in Australia's Northern Territory points to management solutions. Ecol. Manag. Restor. 2010, 11, 127–134. [Google Scholar] [CrossRef]
- Garnett, S.T.; Sithole, B.; Whitehead, P.J.; Burgess, C.P.; Johnston, F.H.; Lea, T. Healthy Country, Healthy People: policy implications of links between Indigenous human heath and environmental condition in tropical Australia. Aust. J. Publ. Admin. 2009, 68, 53–66. [Google Scholar] [CrossRef]
- Johnston, F.H.; Burgess, P.; Bowman, D.M.J.S. A Case for Indigenous Natural Resource Management and Health. In Investing in Indigenous Natural Resource Management; Luckert, M.K., Campbell, B.M., Gorman, J.T., Garnett, S.T., Eds.; Charles Darwin University Press: Darwin, NT, Australia, 2007; pp. 91–95. [Google Scholar]
- Hill, R.; Harding, E.K.; Edwards, D.; O’Dempsey, J.; Hill, D.; Martin, A.; McIntyre-Tamwoy, S. A Cultural and Conservation Economy for Northern Australia; Australian Conservation Foundation: Melbourne, ACT, Australia, 2007. [Google Scholar]
- Fitzsimons, J.; Russell-Smith, J.; James, G.; Vigilante, T.; Lipsett-Moore, G.; Morrison, J.; Looker, M. Insights into the biodiversity and social benchmarking components of the Northern Australian fire management and carbon abatement programmes. Ecol. Manag. Restor. 2012, 13, 51–57. [Google Scholar] [CrossRef]
- Preuss, K.; Dixon, M. ‘Looking after country two-ways’: insights into Indigenous community-based conservation form the Southern Tanami. Ecol. Manag. Restor. 2012, 13, 2–15. [Google Scholar] [CrossRef]
- Wiens, J. The dangers of black-and-white conservation. Conserv. Biol. 2007, 21, 1371–1372. [Google Scholar] [CrossRef]
- Hansen, A.J.; Rotella, J.J. Biophysical factors, land use and species viability in and around Nature Reserves. Conserv. Biol. 2002, 16, 1112–1122. [Google Scholar] [CrossRef]
- Berger, J. Is it acceptable to let a species go extinct in a National Park? Conserv. Biol. 2003, 17, 1451–1454. [Google Scholar] [CrossRef]
- Struhsaker, T.T.; Struhsaker, P.J.; Siex, K.S. Conserving Africa’s rain forests: Problems in protected areas and possible solutions. Biol. Conserv. 2005, 123, 45–54. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).