
Human Impacts on Holocene Vegetation and Wetland Degradation in the Lower Pearl River, Southern China
 , and
, and
Abstract
1. Introduction
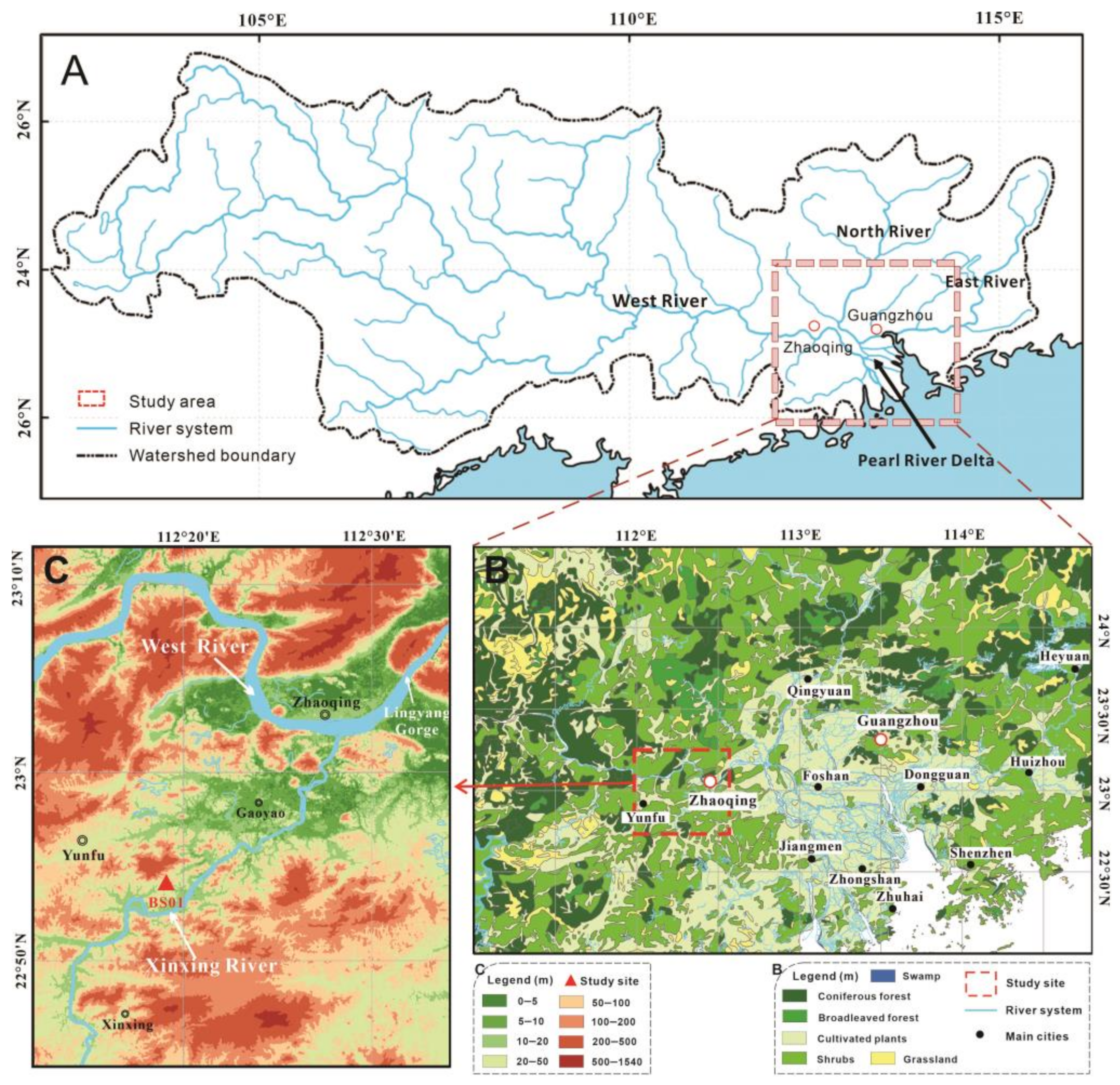
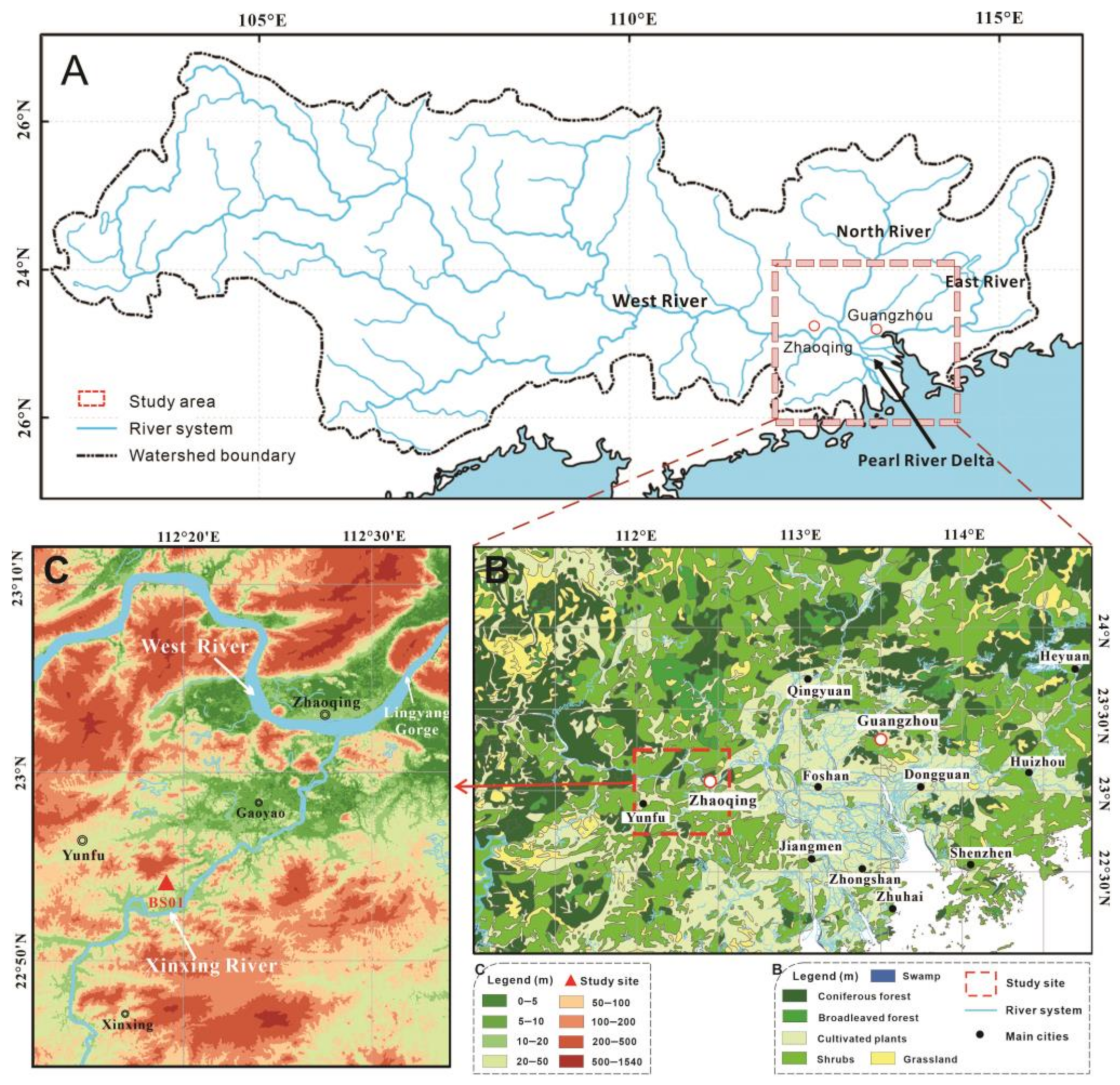
2. Geographic Settings
3. Material and Methods
4. Results
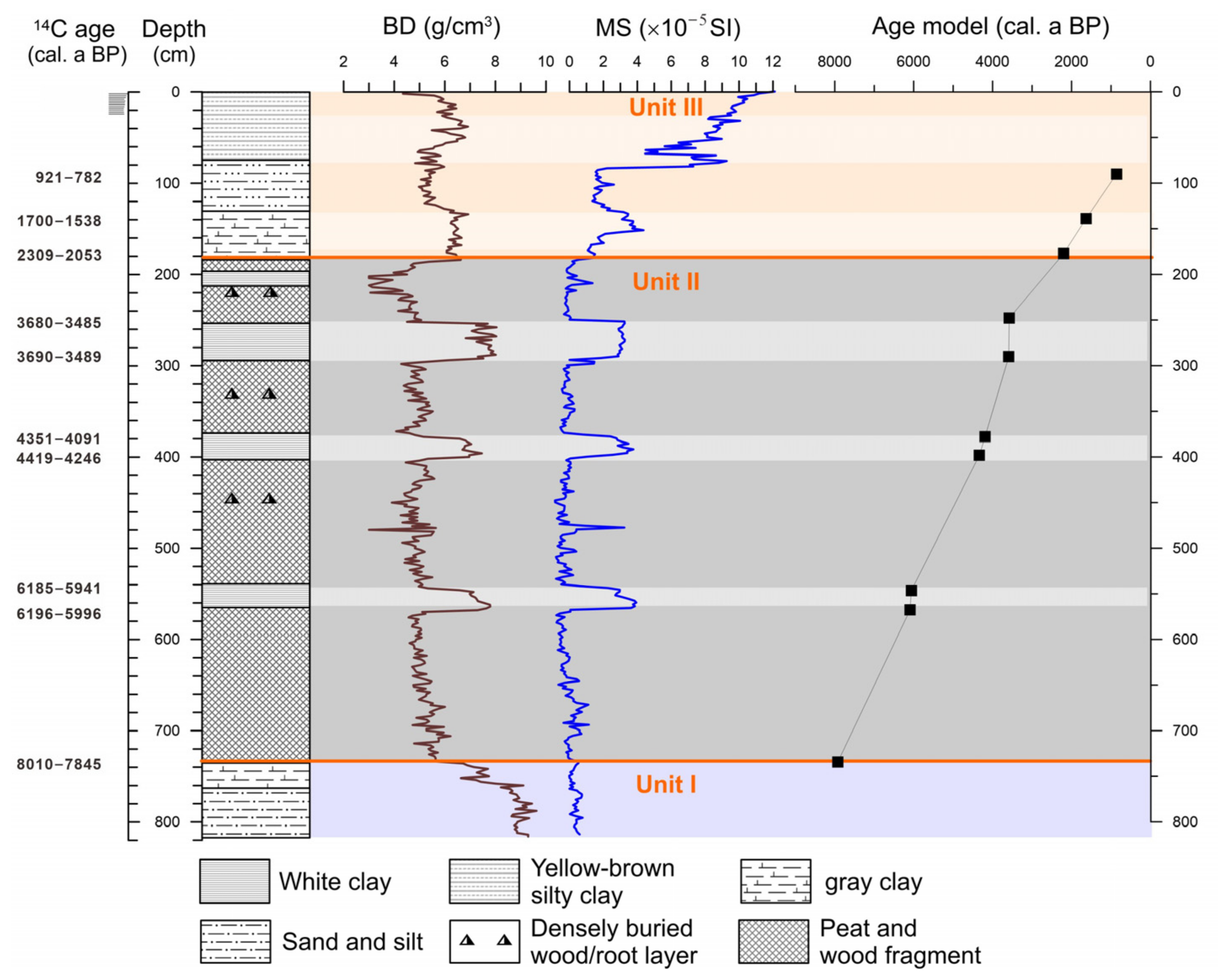
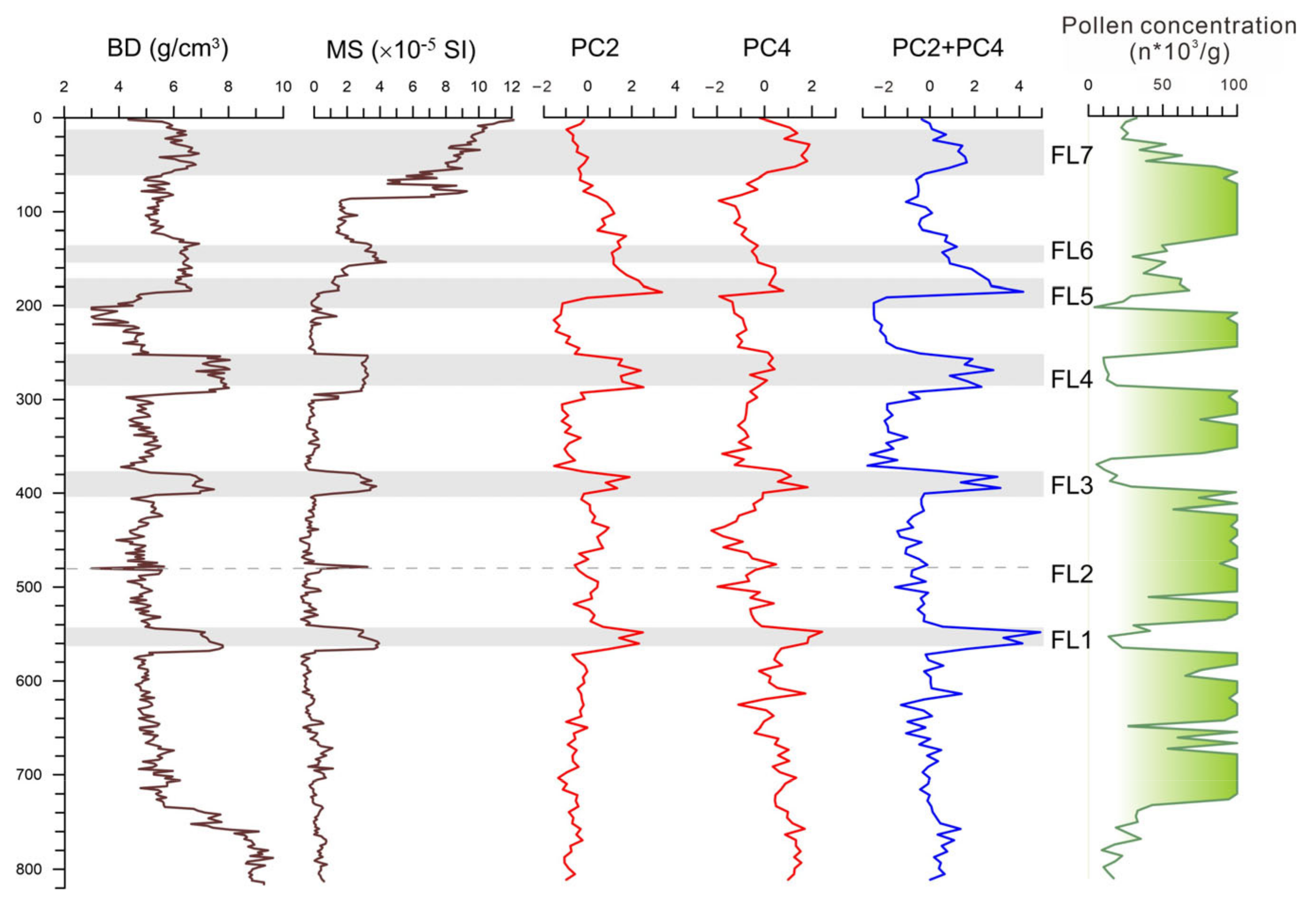
4.1. Lithology and Chronology
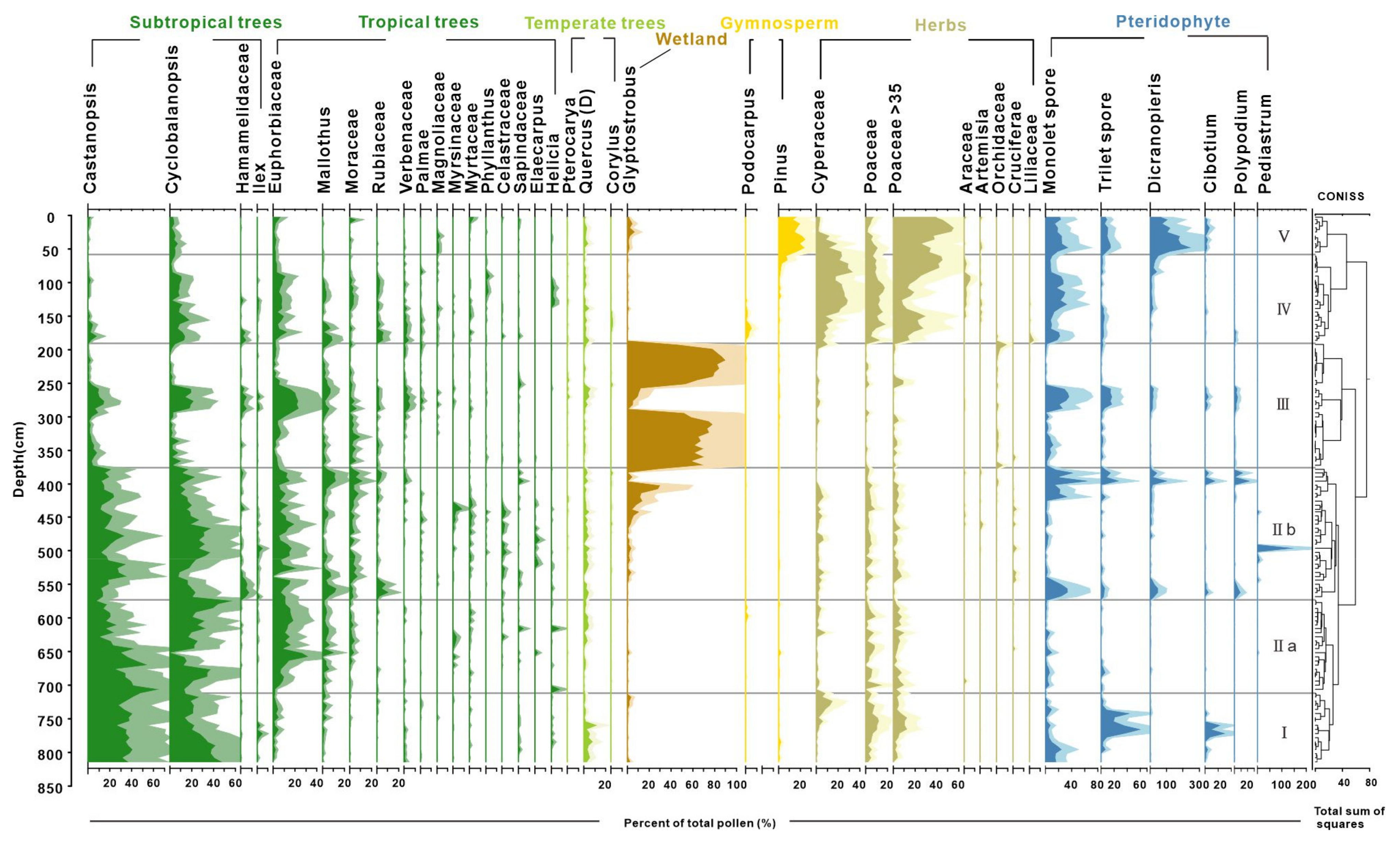
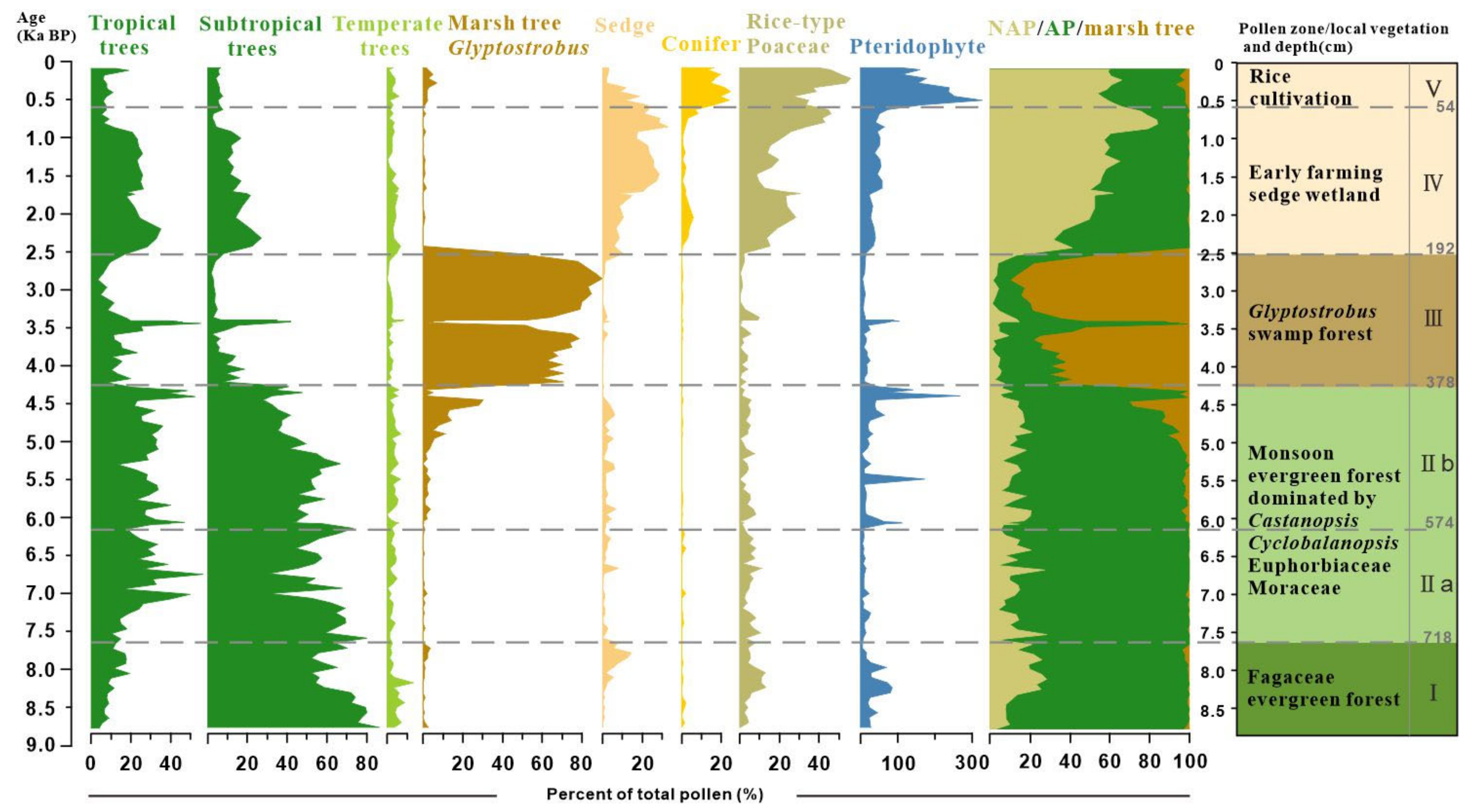
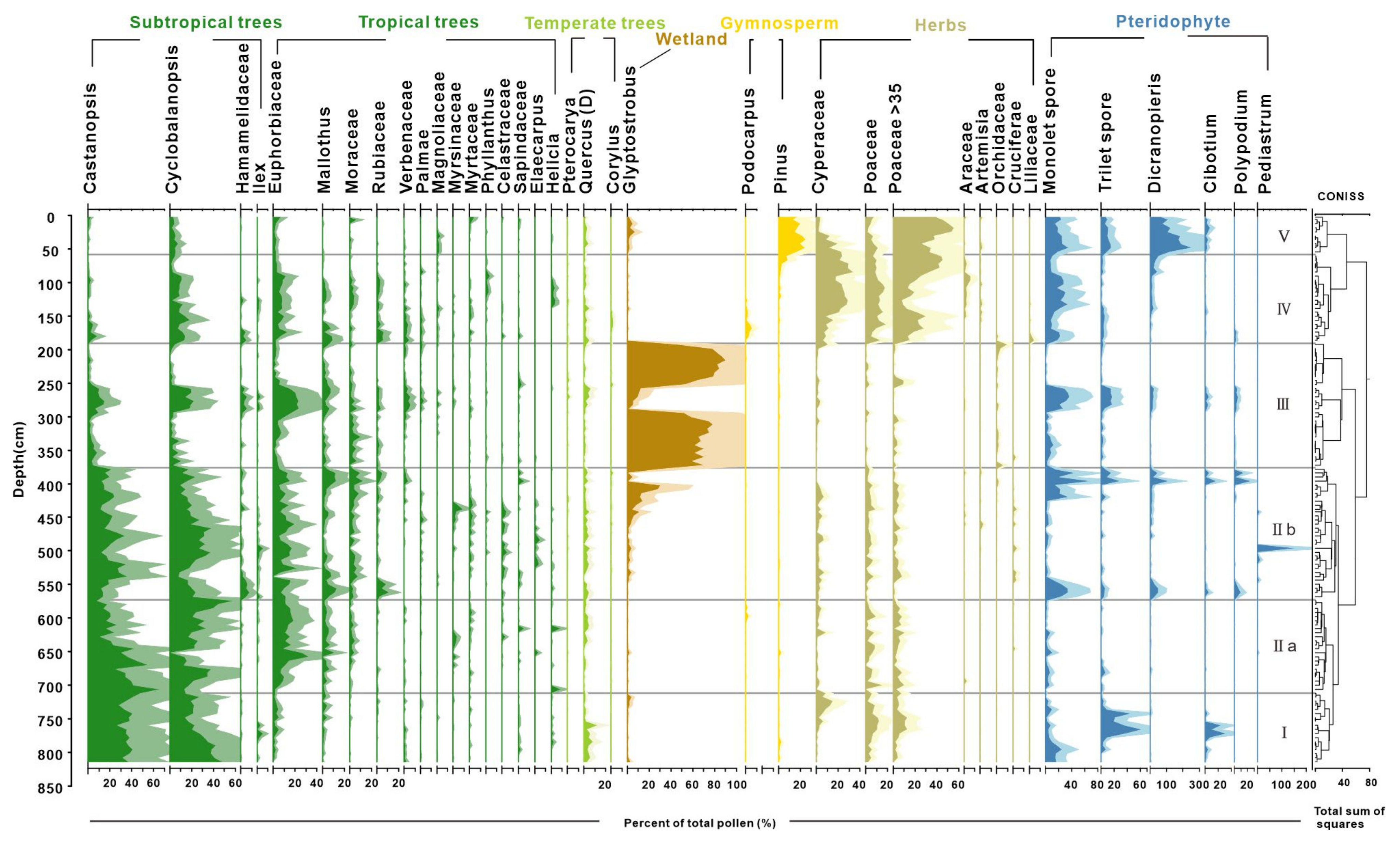
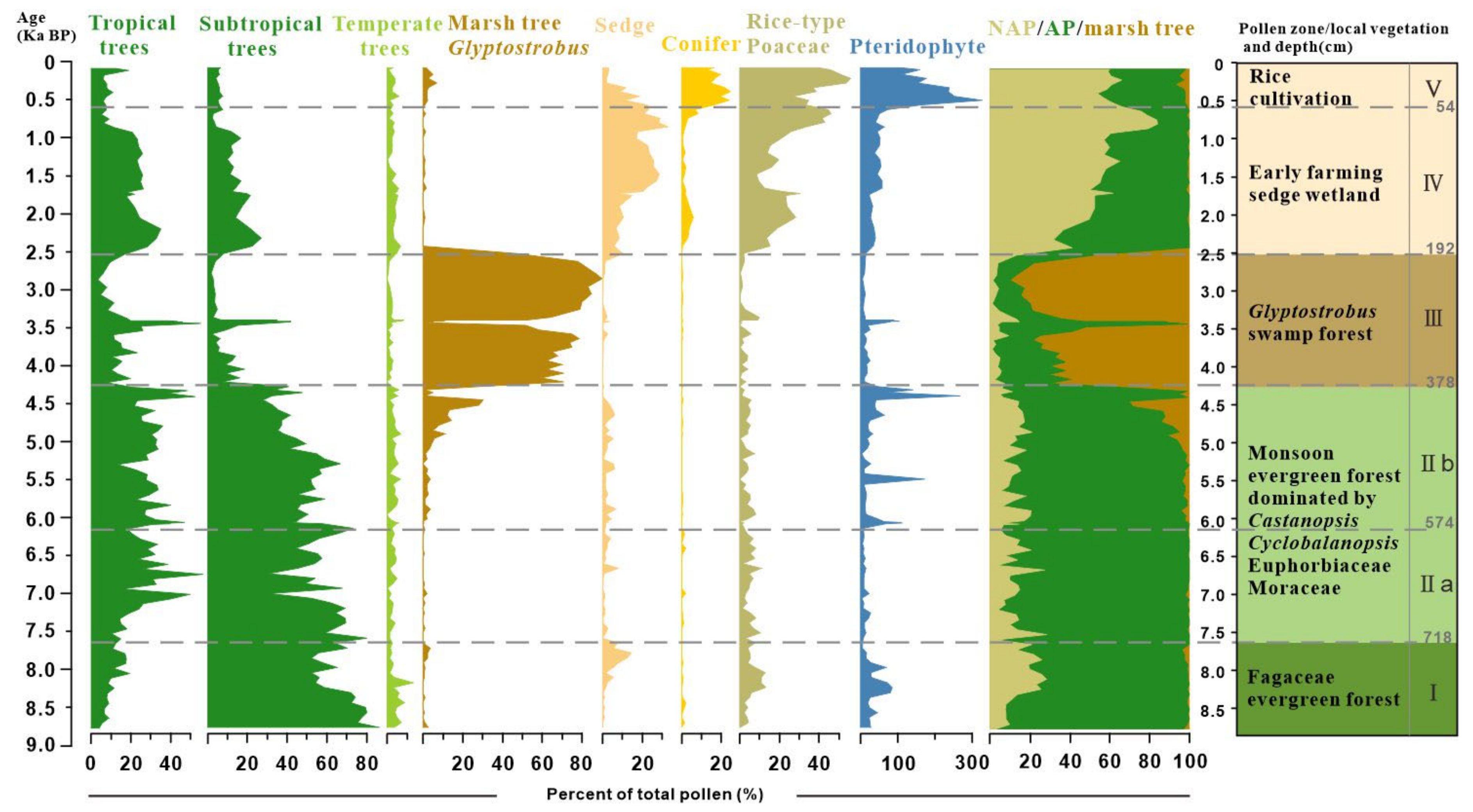
4.2. Result of Pollen Analysis
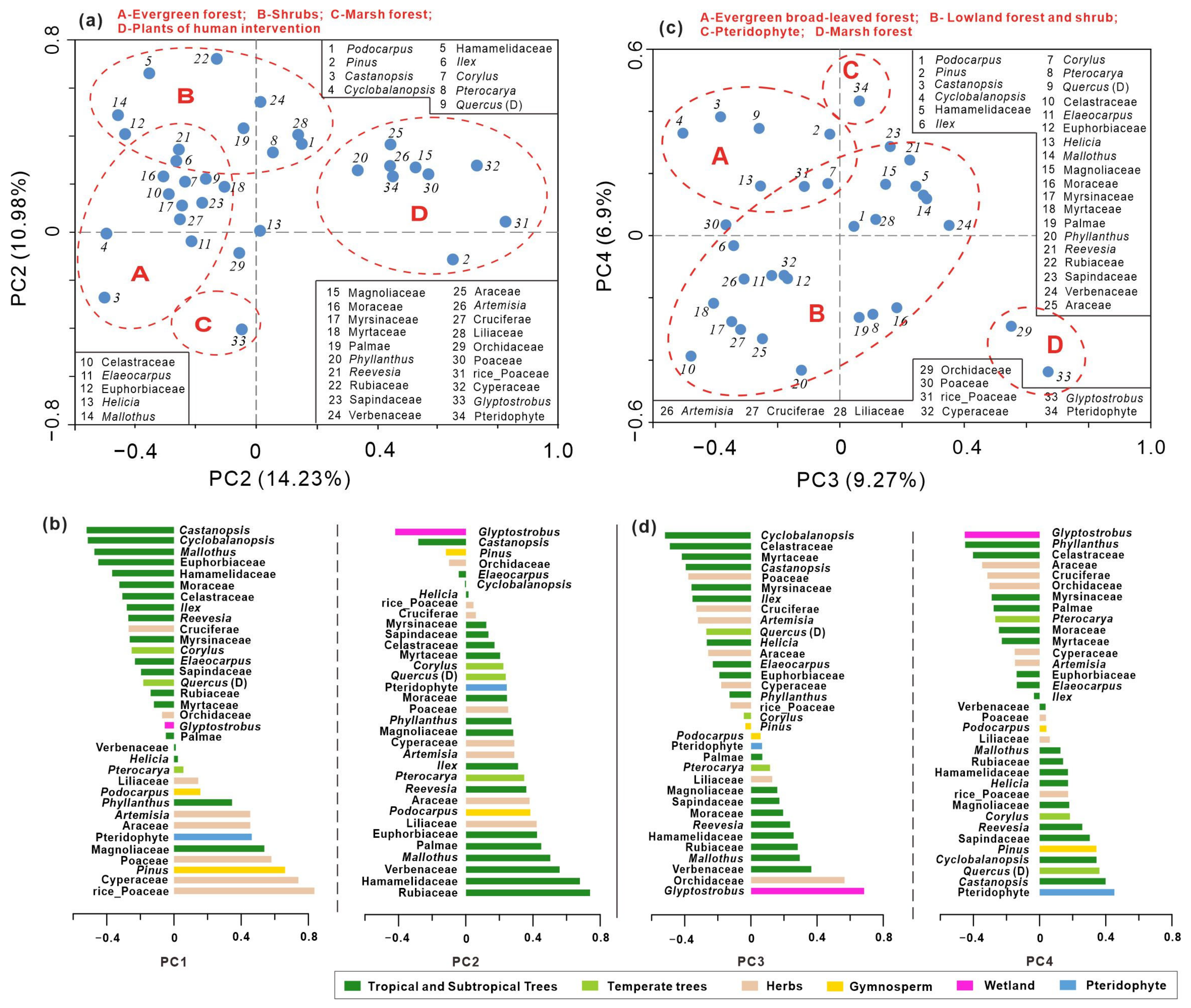
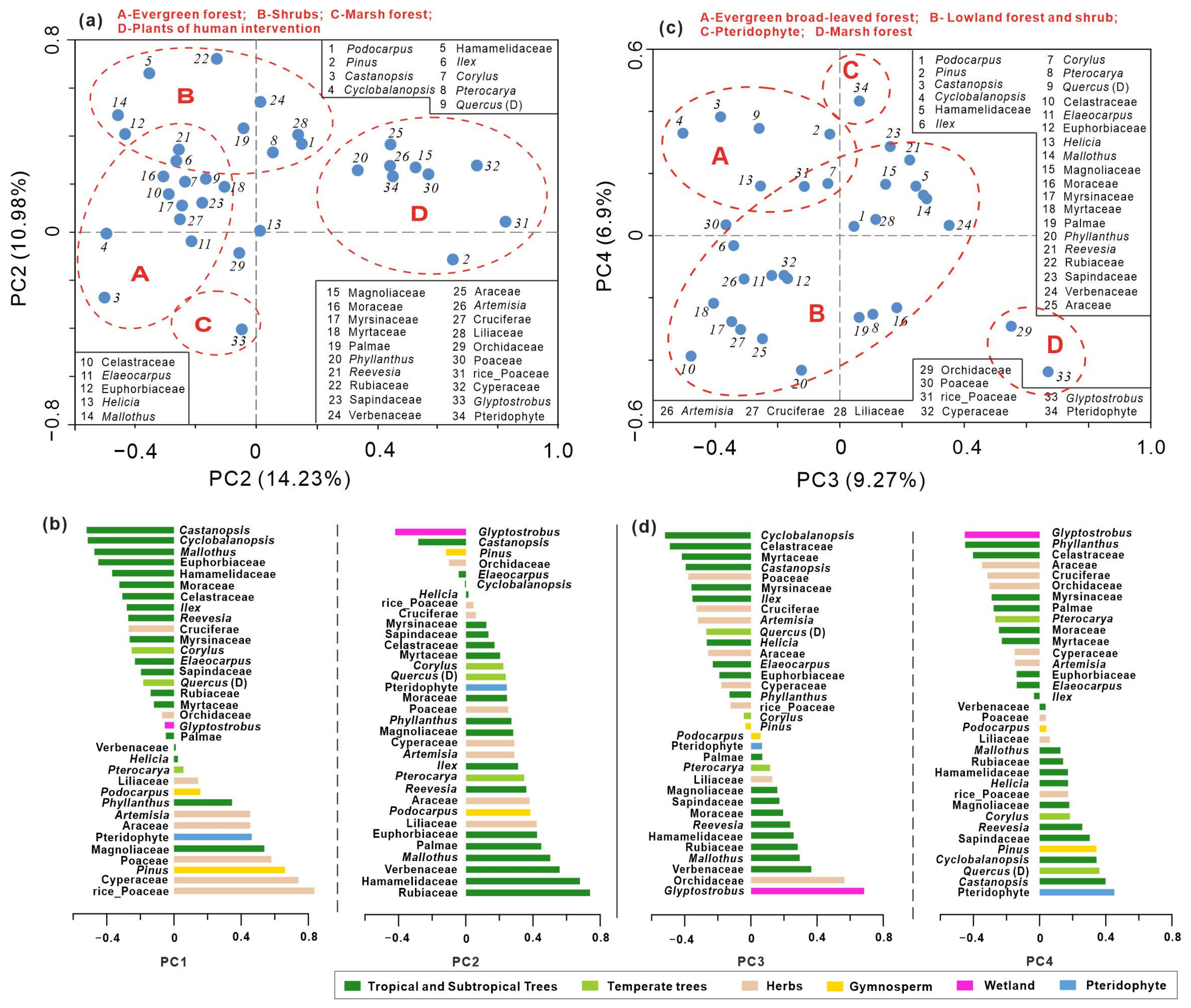
4.3. Principal Component Analysis of Pollen Data
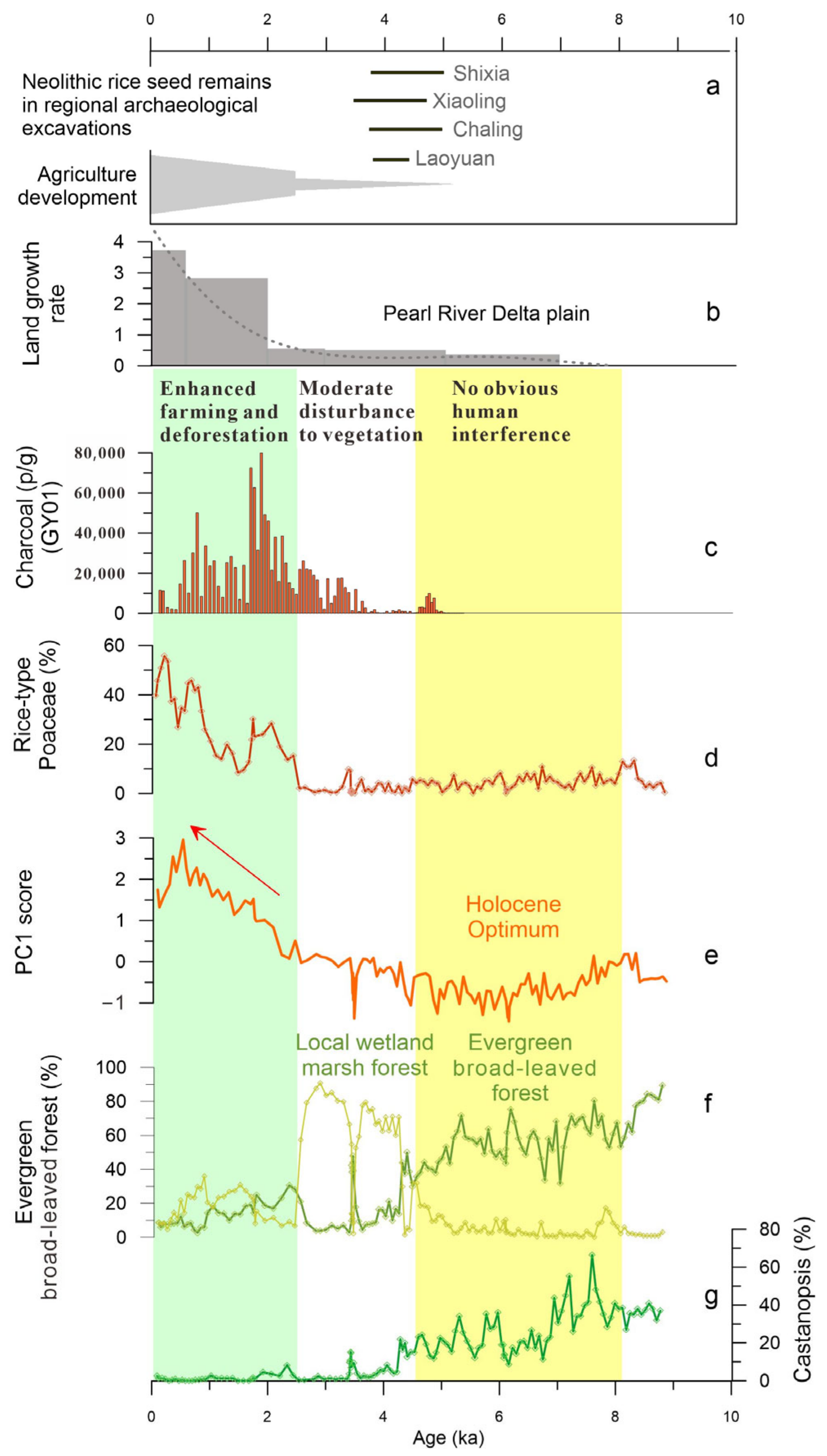
5. Discussion
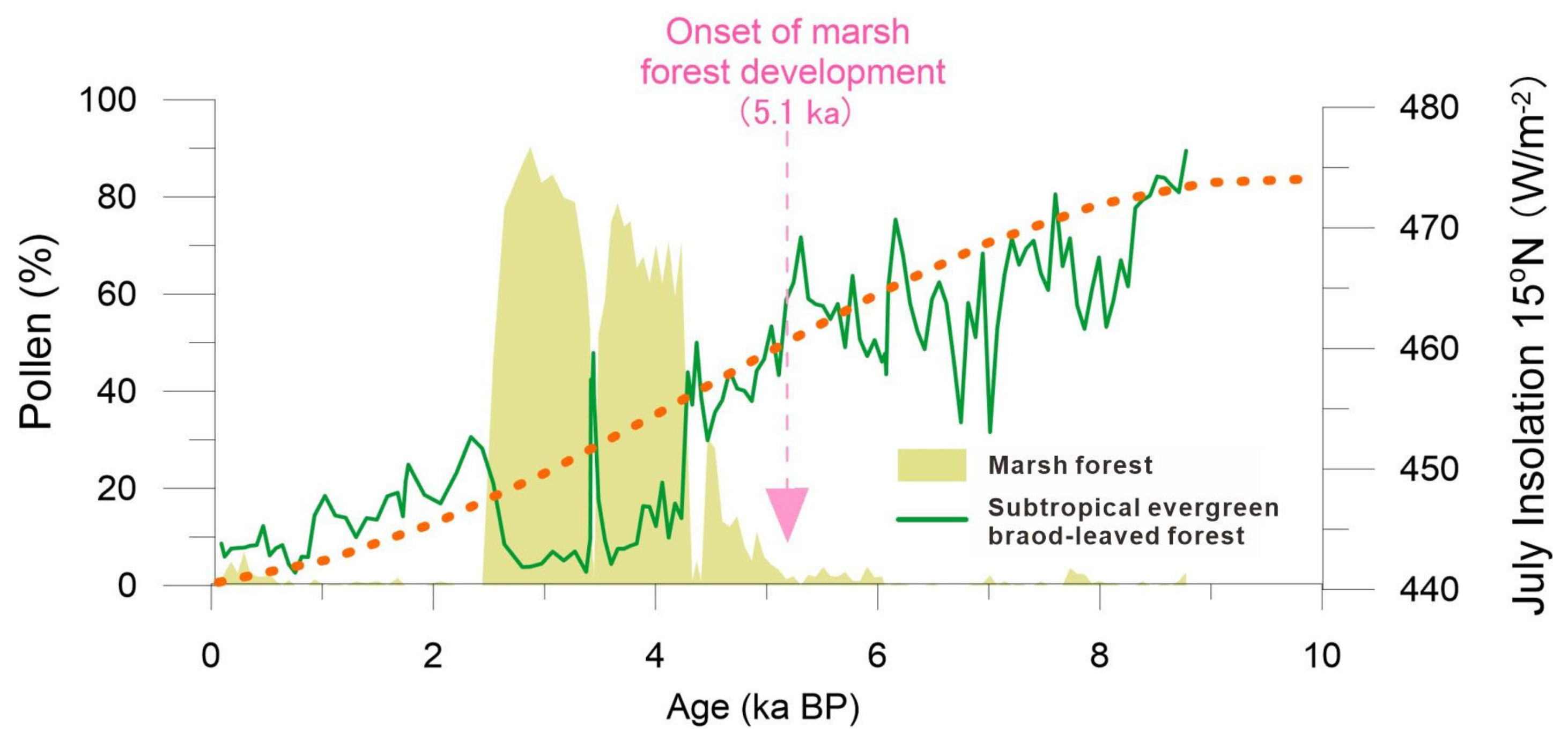
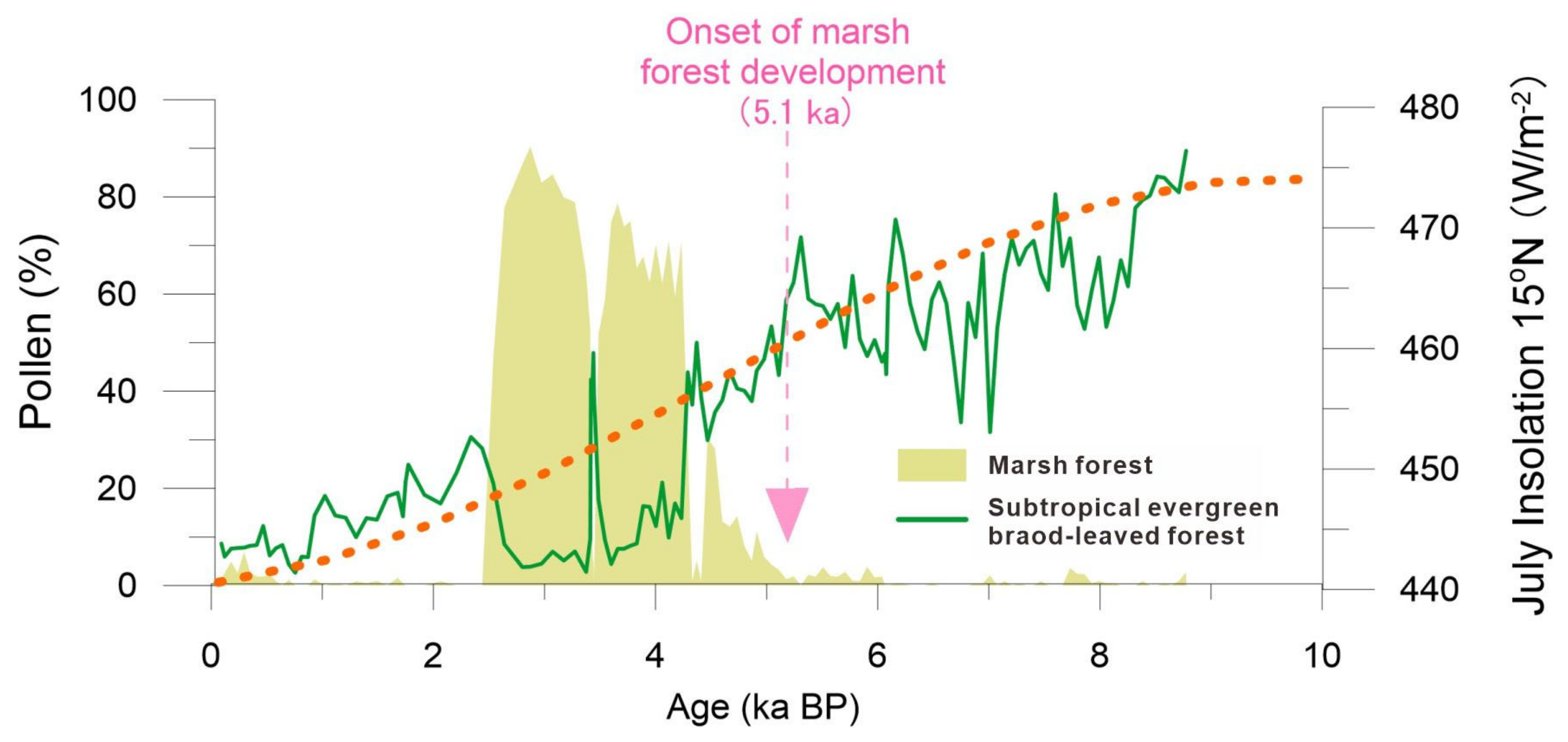
5.1. Holocene Predominance of Evergreen Broad-Leaved Forest and the Development of a Marsh Forest
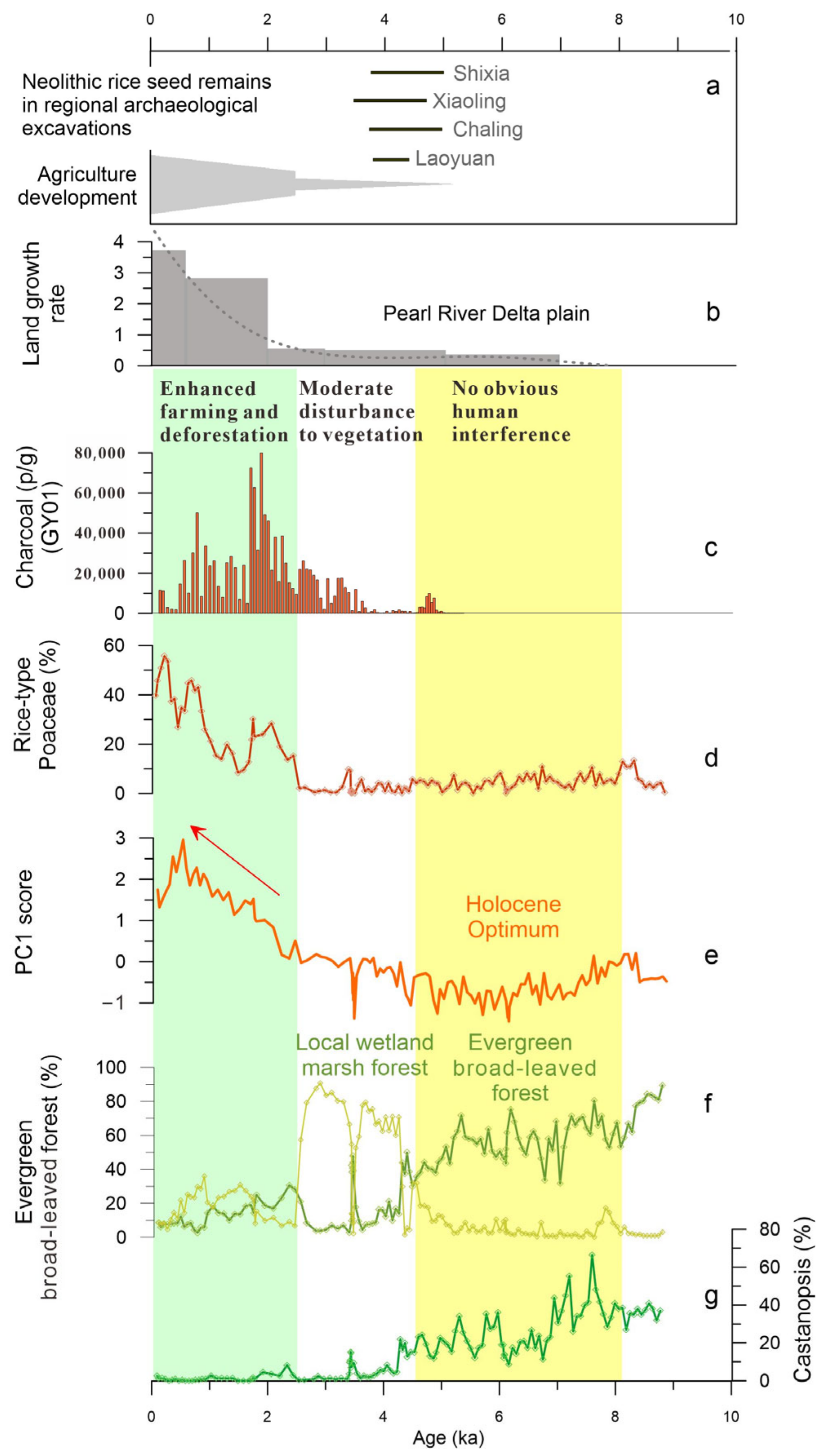
5.2. History of Human Activity and Degradation of Marsh Forest Ecosystem
5.3. Pollen Data as an Indicator of Megaflood Deposition
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dykoski, C.A.; Edwards, R.L.; Cheng, H.; Yuan, D.; Cai, Y.; Zhang, M.; Lin, Y.; Qing, J.; An, Z.; Revenaugh, J. A high-resolution, absolute-dated Holocene and deglacial Asian monsoon record from Dongge Cave, China. Earth Planet. Sci. Lett. 2005, 233, 71–86. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, H.; Edwards, R.L.; He, Y.; Kong, X.; An, Z.; Wu, J.; Kelly, M.J.; Dykoski, C.A.; Li, X. The Holocene Asian monsoon: Links to solar changes and North Atlantic climate. Science 2005, 308, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Henderson, G.M.; Huang, J.; Xie, S.; Sun, Y.; Johnson, K.R. Quantification of Holocene Asian monsoon rainfall from spatially separated cave records. Earth Planet. Sci. Lett. 2008, 266, 221–232. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, H.; Edwards, R.L.; Kong, X.; Shao, X.; Chen, S.; Wu, J.; Jiang, X.; Wang, X.; An, Z. Millennial- and orbital-scale changes in the East Asian monsoon over the past 224,000 years. Nature 2008, 451, 1090–1093. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Chen, C.; Huang, K.; Zhang, X.; Kershaw, P.; Cheng, J.; Li, J.; Yue, Y.; Wan, Q.; Zhang, Y.; et al. Holocene warming and evergreen/deciduous forest replacement across eastern China. Quat. Sci. Rev. 2023, 307, 108057. [Google Scholar] [CrossRef]
- An, Z.; Porter, S.C.; Kutzbach, J.E.; Wu, X.; Wang, S.; Liu, X.; Li, X.; Zhou, W. Asynchronous Holocene optimum of the East Asian monsoon. Quat. Sci. Rev. 2000, 19, 743–762. [Google Scholar] [CrossRef]
- Herzschuh, U.; Winter, K.; Wünnemann, B.; Li, S. A general cooling trend on the central Tibetan Plateau throughout the Holocene recorded by the Lake Zigetang pollen spectra. Quat. Int. 2006, 154–155, 113–121. [Google Scholar] [CrossRef]
- Huang, Z.; Li, P.; Zhang, Z.; Li, K. Wood-rich peat layers of Quaternary system in Pearl River Delta. Trop. Geogr. 1981, 2, 50–55. (In Chinese) [Google Scholar]
- Huang, Z.; Li, P.; Zhang, Z.; Li, K.; Qiao, P. Formation, Development and Evolution of the Pearl River Delta; Popular Science Press, Guangzhou Branch: Guangzhou, China, 1982. (In Chinese) [Google Scholar]
- Li, P.; Cui, H.; Tan, H.; Dai, J.; Shen, C.; Sun, Y. A study on Holocene buried timbers in Guangdong. Trop. Geogr. 2001, 21, 195–197. (In Chinese) [Google Scholar]
- Zheng, Z.; Tang, Y.; Zheng, Y.; Huang, K.; Han, Z.; Zong, Y.; Li, P.; Tan, H. Environmental Changes Inferred from Spatial-temporal Distribution of Holocene Buried Peat Layers in Lower Reaches of the Xijiang and Beijiang and the River Confluence of Pearl River Delta. Trop. Geogr. 2016, 36, 313–325. (In Chinese) [Google Scholar]
- Chen, C.; Zheng, Y.; Zheng, Z.; Zong, Y.; Huang, K.; Rolett, B.V.; Peng, H.; Zhang, X.; Tang, Y.; Wan, Q.; et al. Holocene millennial-scale megaflood events point to ENSO-driven extreme climate changes. Sci. China Earth Sci. 2023, 66, 2530–2545. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, W.; Xia, Y.; Gu, Q.; Yu, W.; Zhang, Y.; Wu, D.; Liu, J.; Zhang, X. History of human disturbance to vegetation in the Southeast Hills of China over the last 2900 years: Evidence from a high resolution pollen record. Palaeogeogr. Palaeoclimat. Palaeoecol. 2022, 598, 111028. [Google Scholar]
- Ma, T.; Rolett, B.V.; Zheng, Z.; Zong, Y. Holocene coastal evolution preceded the expansion of paddy field rice farming. Proc. Natl. Acad. Sci. USA 2020, 117, 24138–24143. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Harvey, E.; Qin, L. Presumed domestication? Evidence for wild rice cultivation and domestication in the fifth millennium BC of the Lower Yangtze region. Antiquity 2007, 81, 316–331. [Google Scholar] [CrossRef]
- Fuller, D.; Qin, L.; Zheng, Y.; Zhao, Z.; Chen, X.; Hosoya, L.; Sun, G. The domestication process and domestication rate in rice: Spikelet bases from the Lower Yangtze. Science 2009, 323, 1607–1610. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D. Pathways to Asian civilizations: Tracing the origins and spread of rice and rice cultures. Rice 2011, 4, 78–92. [Google Scholar] [CrossRef]
- Lu, H.Y. New methods and progress in research on the origins and evolution of prehistoric agriculture in China. Sci. China Earth Sci. 2017, 60, 2141–2159. [Google Scholar] [CrossRef]
- Lipson, M.; Cheronet, O.; Mallick, S.; Rohland, N.; Oxenham, M.; Pietrusewsky, M.; Pryce, T.O.; Willis, A.; Matsumura, H.; Buckley, H.; et al. Ancient genomes document multiple waves of migration in Southeast Asian prehistory. Science 2018, 361, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, W.; Zhuang, Y.; Li, Z.; Ma, Z.; Ma, Y.; Cui, Y.; Wei, J.; Fuller, D.Q. New radiocarbon evidence on early rice consumption and farming in South China. Holocene 2017, 27, 1045–1051. [Google Scholar] [CrossRef]
- Tong, J. The Generalizations and Hydrological Character Analysis of Pearl River Basin. Water Conserv. Sci. Technol. Econ. 2007, 13, 31–33. (In Chinese) [Google Scholar]
- Peng, H.; Zheng, Z.; Zheng, Y.; Huang, K.; Wei, J. Holocene vegetation changes and human activities revealed by a peat sediment core in Gaoyao, Zhaoqing. Quat. Sci. 2015, 35, 742–754. (In Chinese) [Google Scholar]
- Wu, Z.Y. Vegetation of China; Science Press: Beijing, China, 1980. [Google Scholar]
- Zou, S.; Zhou, G.Y.; Zhang, Q.M.; Meng, Z.; Chu, G.W.; Xu, S.; Xia, Y.J. Long-term (1992–2015) dynamics of interspecific associations among tree species in a monsoon evergreen broad-leaved forest in Dinghushan Biosphere Reserve. Acta Ecol. Sin. 2019, 39, 6362–6371. (In Chinese) [Google Scholar] [CrossRef]
- Shen, C.; Ding, P.; Wang, N.; Yi, W.; Ding, X.; Fu, D.; Liu, K.; Zhou, L. Buried ancient forest and implications for paleoclimate since the mid-Holocene in South China. Radiocarbon 2010, 52, 1411–1421. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.; Reimer, R. CALIB rev. 8.2. Available online: http://calib.org/ (accessed on 4 April 2022).
- Nakagawa, T.; Brugiapaglia, E.; Digerfeldt, G.; Reille, M.; Beaulieu, J.; Yasuda, Y. Dense-media separation as a more efficient pollen extraction method for use with organic sediment/deposit samples: Comparison with the conventional method. Boreas 1998, 27, 15–24. [Google Scholar] [CrossRef]
- Stockmarr, J. Tablets with Spores used in Absolute Pollen Analysis. Pollen Spores 1971, 13, 615–621. [Google Scholar]
- Grimm, E.C. CONISS: A FORTRAN 77 Program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Frey, D.G.; Deevey, E.S. Review 1: Numerical tools in palaeolimnology—Progress, potentialities, and problems. J. Paleolimnol. 1998, 20, 307–332. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Zhou, X.; Sun, L.; Zhan, T.; Huang, W.; Zhou, X.; Hao, Q.; Wang, Y.; He, X.; Zhao, C.; Zhang, J.; et al. Time-transgressive onset of the Holocene Optimum in the East Asian monsoon region. Earth Planet. Sci. Lett. 2016, 456, 39–46. [Google Scholar] [CrossRef]
- Renssen, H.; Seppä, H.; Crosta, X.; Goosse, H.; Roche, D.M. Global characterization of the Holocene Thermal Maximum. Quat. Sci. Rev. 2012, 48, 7–19. [Google Scholar] [CrossRef]
- Zong, Y.; Huang, G.; Switzer, A.D.; Yu, F.; Yim, W.W.S. An evolutionary model for the Holocene formation of the Pearl River delta, China. Holocene 2009, 19, 129–142. [Google Scholar] [CrossRef]
- Tang, Y.; Zheng, Z.; Huang, K.; Chen, C.; Chen, Z.; Lu, H.; Wu, W.; Lin, X.; Zhang, X.; Li, H. Holocene Evolution of the Pearl River Delta: Mapping Integral Isobaths and Delta Progradation. J. Mar. Sci. Eng. 2023, 11, 1986. [Google Scholar] [CrossRef]
- Qu, X.; Huang, C.; Rao, Z.; Wu, L.; Luo, Y.; Chen, F.; Li, Y.; Zhao, L.; Liu, L.; Song, Z.; et al. Natural and anthropogenic controls on environmental change during the Holocene based on a multiproxy record obtained from subalpine peatland in southern China. Sci. Total Environ. 2024, 912, 169446. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, H.; Wei, L.; Ji, Y.; Wu, S.; Xu, Q.; Zhao, W.; Zhang, X.; Zhao, Y. Rate of vegetation change in southeast China during the Holocene and its potential drivers. Rev. Palaeobot. Palynol. 2024, 322, 105066. [Google Scholar] [CrossRef]
- Xiao, J.; Lu, H.; Zhou, W.; Zhao, Z.; Hao, R. Evolution of vegetation and climate since the last glacial maximum recorded at Dahu peat site, south China. Sci. China Earth Sci. 2007, 50, 1209–1217. [Google Scholar] [CrossRef]
- Li, P.; Cui, H.; Tan, H.; Feng, Y.; Liu, H.; Mu, Z.; Sun, Y.; Song, L. Climate and Environmental Changes Reflected by Buried Ancient trees in Guangdong. Geomorphology, Environment and Development. In Proceedings of the 2004 Danxia Mountain Conference; The Geographical Society of China: Nanjing, China, 2004. [Google Scholar]
- Zong, Y.; Zheng, Z.; Huang, K.; Sun, Y.; Wang, N.; Tang, M.; Huang, G. Changes in sea level, water salinity and wetland habitat linked to the late agricultural development in the Pearl River delta plain of China. Quat. Sci. Rev. 2013, 70, 145–157. [Google Scholar] [CrossRef]
- Zheng, Z. Study of human disturbance to the vegetation in recent thousands of years in the coastal areas of southeast China. Ecol. Sci. 1998, 17, 30–36. [Google Scholar]
- Yang, X.; Chen, Q.; Ma, Y.; Li, Z.; Hung, H.; Zhang, Q.; Jin, Z.; Liu, S.; Zhou, Z.; Fu, X. New radiocarbon and archaeobotanical evidence reveal the timing and route of southward dispersal of rice farming in south China. Sci. Bull. 2018, 63, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Huang, G. Neolithic culture and paleogeographic environment in the Zhujiang Delta. Acta Geogr. Sinica 1996, 51, 508–517. (In Chinese) [Google Scholar]
- Zhao, S. Environmental change and cultural evolution of the Pearl River Delta in Pre-Qin period. Huaxia Archaeol. 2007, 2, 90–97. (In Chinese) [Google Scholar]
- Xia, X.; Zhang, P.; Wu, Y. The analysis of rice remains from the Chaling site in the Pearl River Delta, Guangdong Province. Quat. Sci. 2019, 39, 24–36. (In Chinese) [Google Scholar]
- Guo, X.; Zhao, W.; Sun, J.; Li, F.; Zhang, K.; Zhao, Y. Advances of Charcoal Study for Paleoenvironment in China. J. Glaciol. Geocryol. 2011, 33, 342–348. [Google Scholar]
- Xiong, H.; Zong, Y.; Huang, G.; Fu, S. Human drivers accelerated the advance of Pearl River deltaic shoreline in the past 7500 years. Quat. Sci. Rev. 2020, 246, 106545. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, Q. Vegetation, climate, and sea level in the past 55,000 years, Hanjiang Delta, Southeastern China. Quat. Res. 2000, 53, 330–340. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (cm) | Sample | Lab. No | Age (ka BP) | ±2σ (cal. ka BP) | Median (cal. ka BP) | Median (BC/AD) |
|---|---|---|---|---|---|---|
| 90 | BS01-3 | BETA369885 | 940 ± 30 | 921–782 | 851 | 1099 AD |
| 139 | BS01-4 | BETA389737 | 1720 ± 30 | 1700–1538 | 1606 | 344 AD |
| 177 | BS01-5 | BETA389738 | 2170 ± 30 | 2309–2053 | 2179 | 229 BC |
| 248 | BS01-6 | BETA369886 | 3350 ± 30 | 3686–3485 | 3547 | 1624 BC |
| 290 | BS01-7 | BETA369887 | 3360 ± 30 | 3689–3489 | 3592 | 1642 BC |
| 378 | BS01-8 | BETA389739 | 3810 ± 30 | 4351–4091 | 4198 | 2248 BC |
| 398 | BS01-9 | BETA389740 | 3910 ± 30 | 4419–4246 | 4346 | 2396 BC |
| 546.5 | BS01-10 | BETA369888 | 5280 ± 30 | 6185–5941 | 6079 | 4129 BC |
| 567.5 | BS01-11 | BETA369889 | 5310 ± 30 | 6196–5996 | 6085 | 4135 BC |
| 734.5 | BS01-12 | BETA369890 | 7100 ± 40 | 8010–7845 | 7932 | 5982 BC |
| Tropical Trees | Subtropical Trees | Wetland |
|---|---|---|
| Apocynaceae | Altingia [Altingiaceae] | Alismataceae |
| Bignoniaceae | Anacardiaceae | Glyptostrobus [Cupressaceae] |
| Caesalpiniaceae | Aralia [Araliaceae] | Iridaceae |
| Celastraceae | Caprifoliaceae | Lobeliaceae |
| Dipterocarpaceae | Castanopsis [Fagaceae] | Nuphar [Nymphaeaceae] |
| Elaecarpus [Elaeocarpaceae] | Cyclobalanopsis [Fagaceae] | Onagraceae |
| Phyllanthus [Euphorbiaceae] | Combretaceae | Polygonum [Polygonaceae] |
| Mallothus [Euphorbiaceae] | Crypteroniaceae | Typha [Typhaceae] |
| Euphorbiaceae | Elaeagnaceae | |
| Flacourtiaceae | Hamamelidaceae | Herbs |
| Guttiferae | Ilex [Aquifoliaceae] | Araceae |
| Helicia [Proteaceae] | Myrica [Myricaceae] | Artemisia [Asteraceae] |
| Loranthaceae | Rhamnus [Rhamnaceae] | Chenopodiaceae |
| Lythraceae | Rosaceae | Commelinaceae |
| Magnoliaceae | Scrophulariaceae | Convolvulaceae |
| Melatomataceae | Theaceae | Cruciferae |
| Melia [Meliaceae] | Thymelaeaceae | Impatiens [Balsaminaceae] |
| Mimosaceae | Ulmus [Ulmaceae] | Lamiaceae |
| Moraceae | Liliaceae | |
| Myrsinaceae | Temperate Trees | Orchidaceae |
| Myrtaceae | Alnus [Betulaceae] | Poaceae |
| Nyssa [Nyssaceae] | Betula [Betulaceae] | Ranunculaceae |
| Oleaceae | Corylus [Betulaceae] | Saururaceae |
| Palmae | Pterocarya [Juglandaceae] | Cyperaceae |
| Papilionaceae | Quercus (D) [Fagaceae] | |
| Passifloraceae | Ulmaceae | Pteridophyte |
| Reevesia [Sterculiaceae] | Cibotium [Cibotiaceae] | |
| Rubiaceae | Dicranopteris [Gleicheniaceae] | |
| Rutaceae | Gymnosperm | Diplopterygium [Gleicheniaceae] |
| Sapindaceae | Podocarpus [Podocarpaceae] | Lycopodium [Lycopodiaceae] |
| Sapotaceae | Pinus [Pinaceae] | Lygodium [Lygodiaceae] |
| Symplocos [Symplocaceae] | Polypodium [Polypodiaceae] | |
| Tiliaceae | Pediastrum [Pediastraceae] | |
| Verbenaceae | Concentricytes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zheng, Y.; Gong, Q.; Fu, S.; Chen, C.; Tang, Y.; Zhang, X.; Wan, Q.; Huang, K.; Zheng, Z. Human Impacts on Holocene Vegetation and Wetland Degradation in the Lower Pearl River, Southern China. Land 2024, 13, 530. https://doi.org/10.3390/land13040530
Zhang Y, Zheng Y, Gong Q, Fu S, Chen C, Tang Y, Zhang X, Wan Q, Huang K, Zheng Z. Human Impacts on Holocene Vegetation and Wetland Degradation in the Lower Pearl River, Southern China. Land. 2024; 13(4):530. https://doi.org/10.3390/land13040530
Chicago/Turabian StyleZhang, Yaze, Yanwei Zheng, Qinghua Gong, Shuqing Fu, Cong Chen, Yongjie Tang, Xiao Zhang, Qiuchi Wan, Kangyou Huang, and Zhuo Zheng. 2024. "Human Impacts on Holocene Vegetation and Wetland Degradation in the Lower Pearl River, Southern China" Land 13, no. 4: 530. https://doi.org/10.3390/land13040530
APA StyleZhang, Y., Zheng, Y., Gong, Q., Fu, S., Chen, C., Tang, Y., Zhang, X., Wan, Q., Huang, K., & Zheng, Z. (2024). Human Impacts on Holocene Vegetation and Wetland Degradation in the Lower Pearl River, Southern China. Land, 13(4), 530. https://doi.org/10.3390/land13040530