
Effects of Long-Term Soil Tillage Practices on Soil Organic C Accumulation Characteristics in Double-Cropped Rice Paddy

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Soil Sampling
2.4. Soil Laboratory Analysis
Soil Chemical Analysis
2.5. Statistical Analysis
3. Results
3.1. Effects on Soil Physical Properties
3.2. Effects on SOC
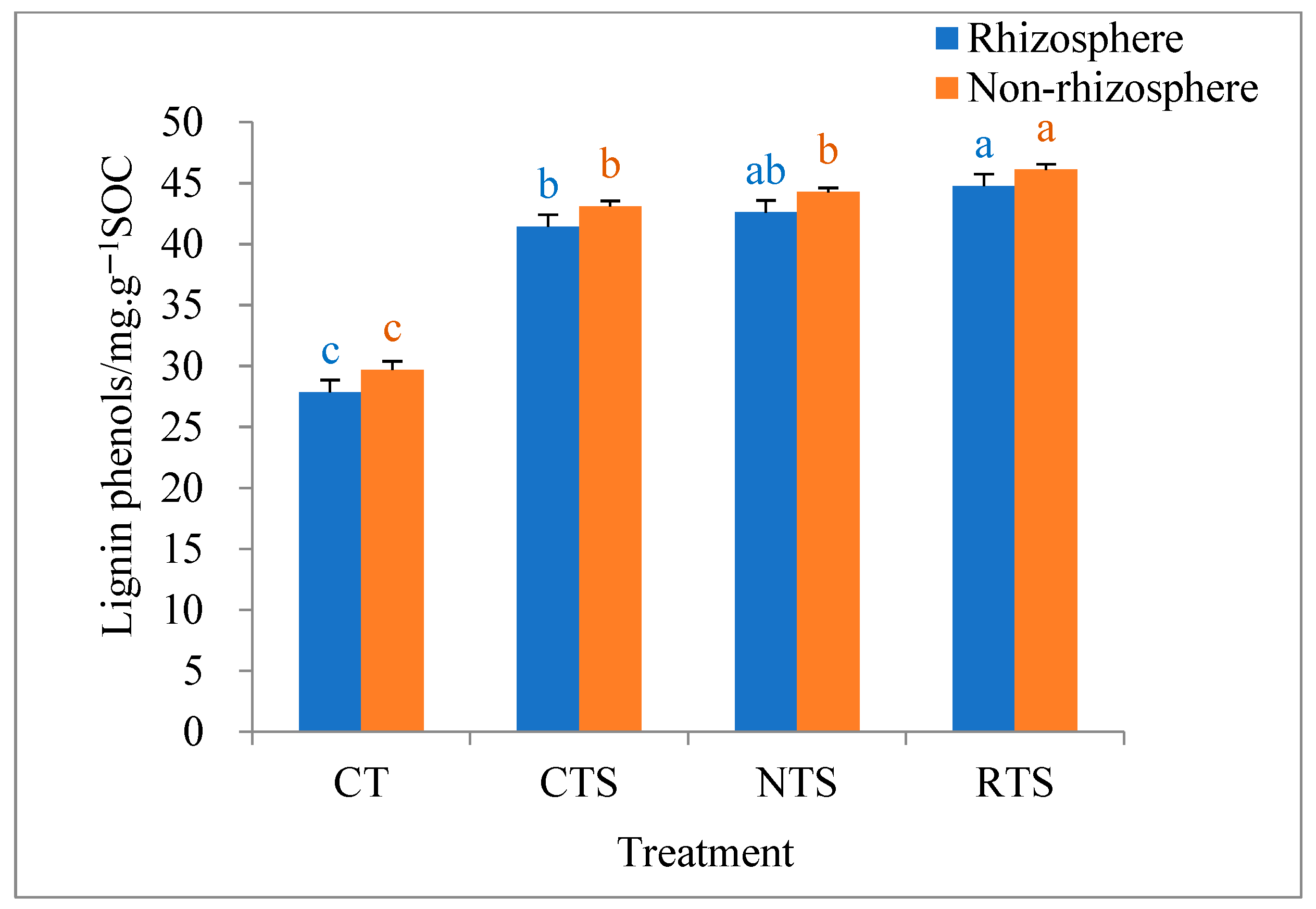
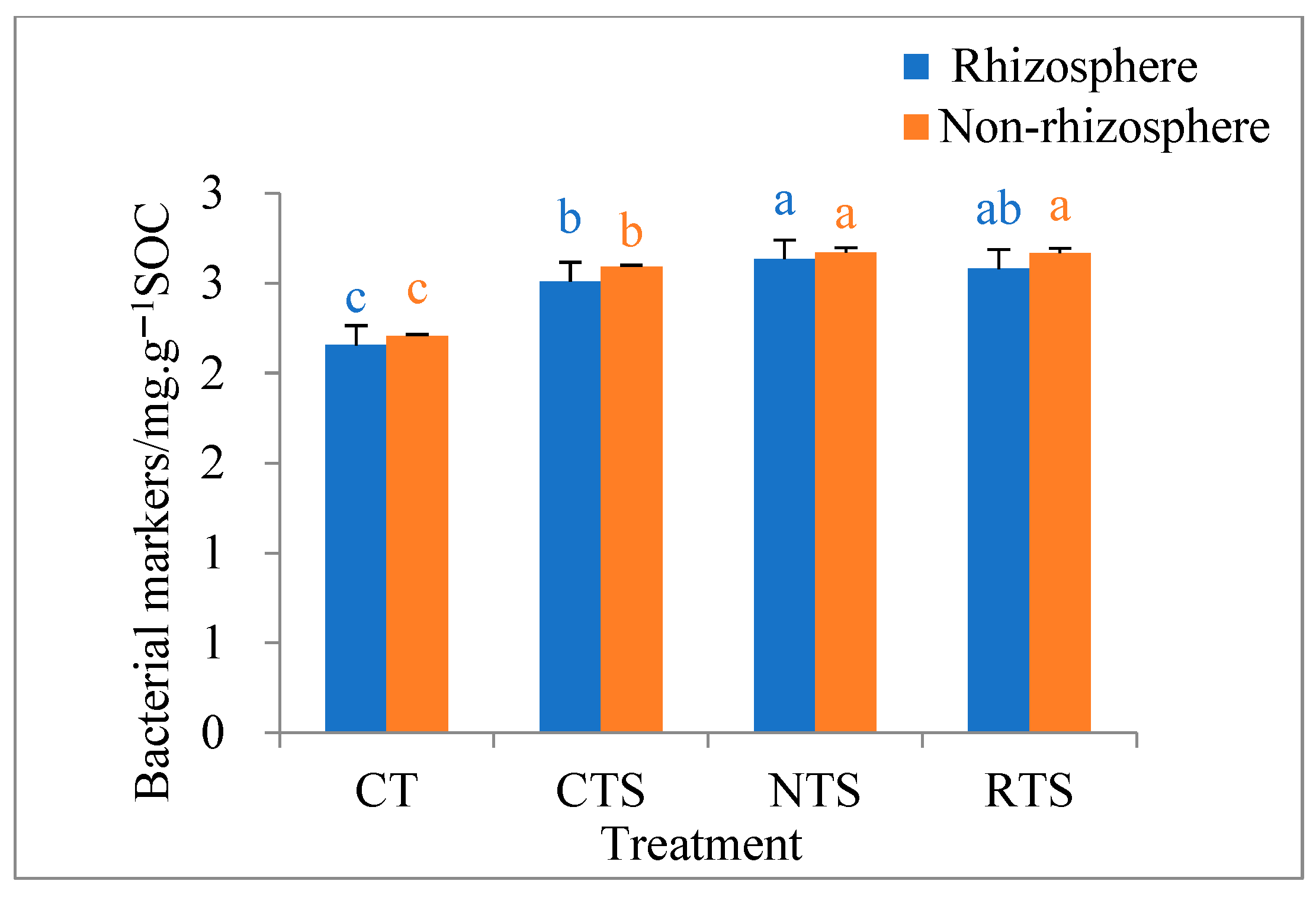
3.3. Effects on Lignin Phenol and Amino Sugar Contents
3.4. Impact on SOC Sequestration Sources in Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, H.M.; Xiao, X.P.; Li, C.; Tang, W.G.; Cheng, K.K.; Pan, X.C.; Wang, K.; Li, W.Y. Effects of different soil tillage systems on soil carbon management index under double-cropping rice paddy field in southern China. Agron. J. 2019, 111, 440–447. [Google Scholar] [CrossRef]
- Ghosh, B.N.; Meena, V.S.; Singh, R.J.; Alam, N.M.; Patra, S.; Bhattacharyya, R.; Sharma, N.K.; Dadhwal, K.S.; Mishra, P.K. Effects of fertilization on soil aggregation, carbon distribution and carbon management index of maize-wheat rotation in the north-western Indian Himalayas. Ecol. Indic. 2019, 105, 415–424. [Google Scholar] [CrossRef]
- Lei, K.J.; Dai, W.X.; Wang, J.; Li, Z.W.; Cheng, Y.; Jiang, Y.J.; Yin, W.Q.; Wang, X.Z.; Song, X.D.; Tang, Q. Biochar and Straw Amendments over a Decade Divergently Alter Soil Organic Carbon Accumulation Pathways. Agronomy 2024, 14, 2176. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jackson, R.D.; DeLucia, E.H.; Tiedje, J.M.; Liang, C. The soil microbial carbon pump: From conceptual insights to empirical assessments. Glob. Chang. Biol. 2020, 26, 6032–6039. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Lehmann, J.; Hansel, C.M.; Kaiser, C.; Kleber, M.; Maher, K.; Manzoni, S.; Nunan, N.; Reichstein, M.; Schimel, J.P.; Torn, M.S. Persistence of soil organic carbon caused by functional complexity. Nat. Geosci. 2020, 13, 529–534. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef]
- Sokol, N.W.; Sanderman, J.; Bradford, M.A. Pathways of mineral-associated soil organic matter formation: Integrating the role of plant carbon source, chemistry, and point of entry. Glob. Chang. Biol. 2019, 25, 12–24. [Google Scholar] [CrossRef]
- Joergensen, R.G. Amino sugars as specific indices for fungal and bacterial residues in soil. Biol. Fert. Soils 2018, 54, 559–568. [Google Scholar] [CrossRef]
- Liu, X.; Ma, T.; Zhang, H.G.; Yan, K.; Zhou, S.W. The accumulation of fungal not bacterial residue carbon is management-dependent under conventional and organic practices in apple-orchard soil. Geoderma 2024, 448, 116968. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, P. The Microbial Eficiency-Matrix S tabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter. Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Chen, X.B.; Xia, Y.H.; Rui, Y.; Ning, Z.; Hu, Y.J.; Tang, H.M.; He, H.B.; Li, H.X.; Kuzyakov, Y.; Ge, T.D.; et al. Microbial carbon use efficiency, biomass turnover, and necromass accumulation in paddy soil depending on fertilization. Agric. Ecosyst. Environ. 2020, 292, 106816. [Google Scholar] [CrossRef]
- Crow, S.E.; Lajtha, K.; Filley, T.R.; Swanston, C.W.; Bowden, R.D.; Caldwell, B.A. Sources of plant-derived carbon and stability of organic matter in soil: Implications for global change. Glob. Chang. Biol. 2009, 15, 2003–2019. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.C.; Luo, J.F.; Lindsey, S.; Zhou, F.; Xie, H.T.; Li, Y.; Zhu, P.; Wang, L.C.; Shi, Y.L.; et al. Differential accumulation of microbial necromass and plant lignin in synthetic versus organic fertilizer-amended soil. Soil Biol. Biochem. 2020, 149, 107967. [Google Scholar] [CrossRef]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef]
- Pan, G.; Li, L.; Wu, L.; Zhang, X. Storage and sequestration potential of topsoil organic carbon in China’s paddy soils. Glob. Chang. Biol. 2004, 10, 79–92. [Google Scholar] [CrossRef]
- Tian, K.; Zhao, Y.; Xu, X.; Hai, N.; Huang, B.; Deng, W. Effects of long-term fertilization and residue management on soil organic carbon changes in paddy soils of China: A meta-analysis. Agric. Ecosyst. Environ. 2015, 204, 40–50. [Google Scholar] [CrossRef]
- Liu, Y.L.; Ge, T.D.; Zhu, Z.K.; Liu, S.L.; Luo, Y.; Li, Y.; Wang, P.; Gavrichkova, O.; Xu, X.L.; Wang, J.K.; et al. Carbon input and allocation by rice into paddy soils: A review. Soil Biol. Biochem. 2019, 133, 97–107. [Google Scholar] [CrossRef]
- Cheng, K.K.; Tang, H.M.; Li, C.; Tang, W.G.; Xiao, X.P.; Yi, Z.Y. Impact of long-term tillage management on utilization of microbial carbon sources in rhizosphere and non-rhizosphere soils under a double-cropping rice paddy field. Environ. Sci. Pollut. Res. 2022, 29, 15205–15214. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.Y.; Cheng, C.; Wang, J.J.; Luo, K.Z.; Zeng, Y.G.; Yang, X.X. Net global warming potential, greenhouse gas intensity and carbon footprint as affected by different tillage systems from Chinese double-cropping paddy fields. Soil Till. Res. 2021, 209, 104947. [Google Scholar] [CrossRef]
- Wang, X.; Jing, Z.H.; He, C.; Liu, Q.Y.; Qi, J.Y.; Zhao, X.; Xiao, X.P.; Zhang, H.L. Temporal variation of SOC storage and crop yield and its relationship-A fourteen year field trial about tillage practices in a double paddy cropping system, China. Sci. Total Environ. 2021, 759, 143494. [Google Scholar] [CrossRef]
- Choudhary, S.; Gajanand, M.C.; Choudhary, M.S. Conservation Agriculture and its Impact on Physical, Chemical and Biological Properties of Soil: A Review. IJBS 2021, 8, 113–122. [Google Scholar] [CrossRef]
- Crittenden, S.J.; Poot, N.; Heinen, M. Soil physical quality in contrasting tillage systems in organic and conventional tillage. Soil Till. Res. 2015, 154, 136–144. [Google Scholar] [CrossRef]
- Govaerts, B.; Mezzalama, M.; Sayre, K.D.; Crossa, J.; Nicol, J.; Deckers, J. Long-term consequences of tillage, residue management, and crop rotation on maize/wheat root rot and nematode populations in subtropical highlands. Appl. Soil Ecol. 2006, 32, 305–315. [Google Scholar] [CrossRef]
- Van Groenigen, K.J.; Bloem, J.; Bååth, E.; Boeckx, P.; Rousk, J.; Bodé, S.; Forristal, D.; Jones, M.B. Abundance, production and stabilization of microbial biomass under conventional and reduced tillage. Soil Biol. Biochem. 2010, 42, 48–55. [Google Scholar] [CrossRef]
- Qi, J.Y.; Wang, X.; Zhao, X.; Pu, C.; Kan, Z.R.; Li, C.; Liu, P.; Xiao, X.P.; Rattan, L.; Zhang, H.L. Temporal variability of soil organic C in paddies during 13-year conservation tillage. Land Degrad. Dev. 2019, 30, 1840–1850. [Google Scholar] [CrossRef]
- Klute, A. Physical and mineralogical methods. Am. Soc. Agron. Soil Sci. Soc. Am. J. 1986, 2, 463–478. [Google Scholar]
- Regalado, C.M.; Carpena, R.M.; Socorro, A.R.; Moreno, J.M.H. Time domain reflectometry models as a tool to understand the dielectric response of volcanic soils. Geoderma 2003, 117, 313–330. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Chen, X. Economic potential of biomass supply from crop residues in China. Appl. Energy 2016, 166, 141–149. [Google Scholar] [CrossRef]
- Ye, G.; Lin, Y.; Kuzyakov, Y.; Liu, D.; Ding, W. Manure over crop residues increases soil organic matter but decreases microbial necromass relative contribution in upland Ultisols: Results of a 27-year field experiment. Soil Biol. Biochem. 2019, 134, 15–24. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Kögel, I. Estimation and decomposition pattern of the lignin component in forest humus layers. Soil Biol. Biochem. 1986, 18, 589–594. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Volumetric Water Content (g·kg−1) | Permeability Resistance (MPa) | Bulk Density (g·cm−3) | |||
|---|---|---|---|---|---|---|
| Early Rice | Late Rice | Early Rice | Late Rice | Early Rice | Late Rice | |
| CT | 186.2 ± 6.9 c | 191.8 ± 5.2 c | 2.68 ± 0.05 a | 2.74 ± 0.06 a | 1.10 ± 0.02 b | 1.12 ± 0.03 b |
| CTS | 253.2 ± 6.2 b | 263.6 ± 7.4 b | 2.51 ± 0.04 ab | 2.62 ± 0.05 ab | 1.07 ± 0.03 b | 1.08 ± 0.03 b |
| NTS | 281.4 ± 8.6 a | 283.3 ± 6.4 a | 1.82 ± 0.04 c | 1.91 ± 0.04 c | 1.38 ± 0.03 a | 1.38 ± 0.03 a |
| RTS | 273.2 ± 7.4 ab | 277.7 ± 6.5 ab | 2.33 ± 0.06 b | 2.39 ± 0.05 b | 1.17 ± 0.03 b | 1.18 ± 0.03 b |
| Treatment | Early Rice (SOC) g·kg−1 | Late Rice (SOC) g·kg−1 | ||||
|---|---|---|---|---|---|---|
| 0–5 cm | 5–10 cm | 10–20 cm | 0–5 cm | 5–10 cm | 10–20 cm | |
| CT | 31.5 c | 28.7 ab | 21.3 b | 30.5 c | 29.9 ab | 23.4 b |
| CTS | 33.0 b | 31.5 a | 25.2 a | 33.2 b | 32.5 a | 26.4 a |
| NTS | 40.4 a | 25.4 c | 21.4 b | 41.1 a | 26.1 c | 20.8 c |
| RTS | 33.6 b | 28.8 b | 21.8 b | 34.7 b | 29.2 b | 21.3 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, K.; Peng, S.; Li, C.; Wen, L.; Liu, L.; Luo, H.; Liu, J.; Tang, H. Effects of Long-Term Soil Tillage Practices on Soil Organic C Accumulation Characteristics in Double-Cropped Rice Paddy. Land 2024, 13, 2074. https://doi.org/10.3390/land13122074
Cheng K, Peng S, Li C, Wen L, Liu L, Luo H, Liu J, Tang H. Effects of Long-Term Soil Tillage Practices on Soil Organic C Accumulation Characteristics in Double-Cropped Rice Paddy. Land. 2024; 13(12):2074. https://doi.org/10.3390/land13122074
Chicago/Turabian StyleCheng, Kaikai, Shuting Peng, Chao Li, Li Wen, Lingling Liu, Hanfang Luo, Jie Liu, and Haiming Tang. 2024. "Effects of Long-Term Soil Tillage Practices on Soil Organic C Accumulation Characteristics in Double-Cropped Rice Paddy" Land 13, no. 12: 2074. https://doi.org/10.3390/land13122074
APA StyleCheng, K., Peng, S., Li, C., Wen, L., Liu, L., Luo, H., Liu, J., & Tang, H. (2024). Effects of Long-Term Soil Tillage Practices on Soil Organic C Accumulation Characteristics in Double-Cropped Rice Paddy. Land, 13(12), 2074. https://doi.org/10.3390/land13122074