4.1. Influence of Grazing Intensity on Herbaceous and Woody Species Composition
Grazing intensity significantly influenced the composition and abundance of herbaceous and woody species across the grazing zones. Heavily grazed areas exhibited a reduction in herbaceous species alongside an increase in less palatable woody species like
Dichrostachys cinerea and
Senegalia mellifera, corroborating findings from [
4,
21]. This shift toward woody dominance under heavy grazing reduces high-quality forage availability, impacting rangeland productivity. Moderately grazed areas displayed a mix of herbaceous and woody species, supporting greater species diversity and aligning with the intermediate disturbance hypothesis [
17]. Lightly grazed zones, with the highest species richness and palatable vegetation, promote forage sustainability and ecosystem stability, as supported by [
16]. Meanwhile, riparian areas hosted abundant deep-rooted woody species like
Vachellia tortilis, which aid in soil stabilization but may limit herbaceous cover and access to water.
4.3. Seasonal Biomass Dynamics and Grazing Pressure
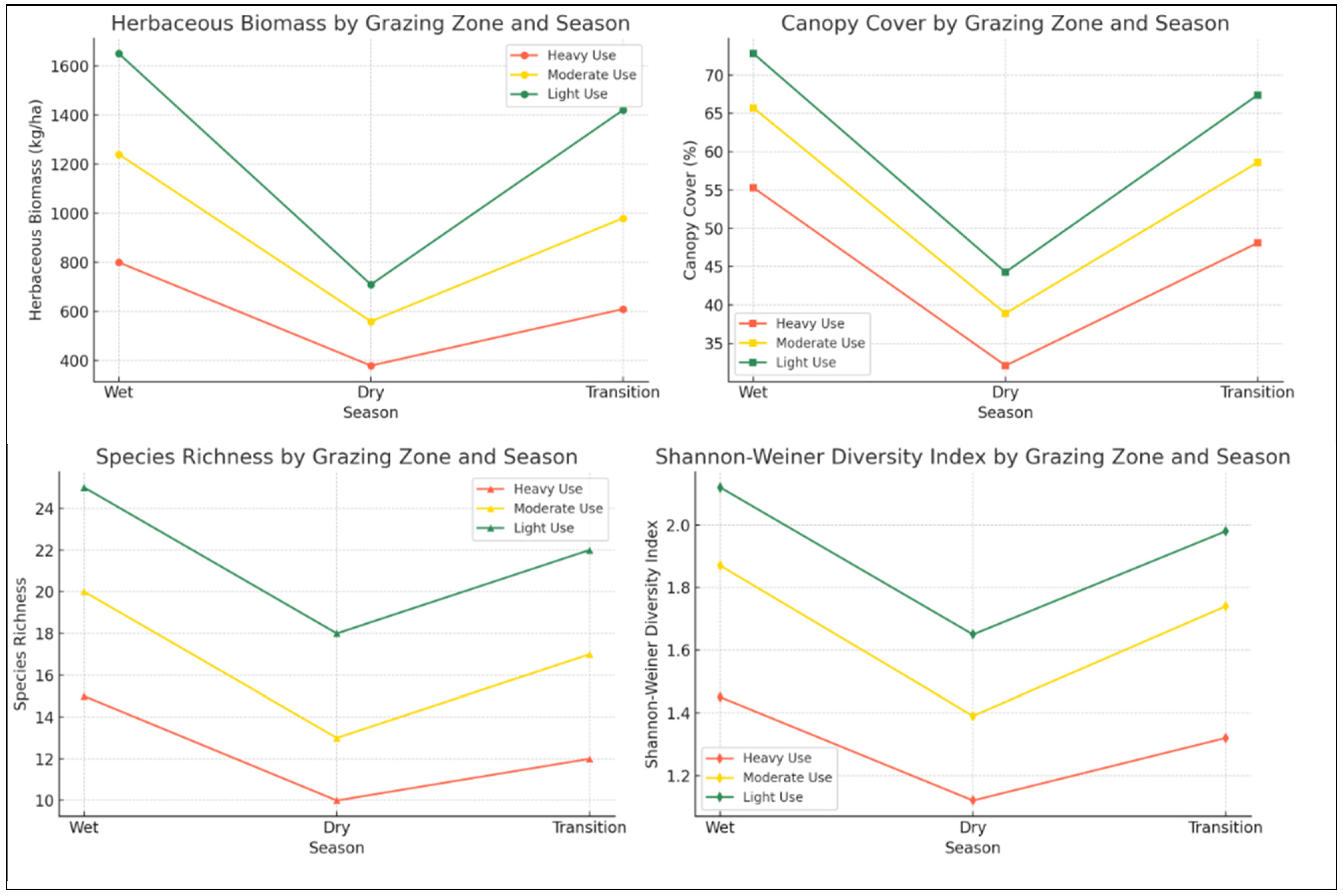
Seasonal fluctuations in herbaceous biomass and cover were prominent, with wet-season growth enhancing forage availability, especially in lightly grazed and riparian areas [
10,
36]. Heavily grazed zones, however, showed significant dry-season declines due to the compounded effects of grazing and moisture stress, leading to reduced forage for livestock [
37]. In contrast, moderately and lightly grazed zones retained higher biomass during the dry season, supported by drought-resistant species like
Cenchrus ciliaris [
38]. Riparian zones, with access to groundwater, exhibited resilience to seasonal droughts, serving as critical dry-season refuges [
39]. Effective management of grazing in riparian and heavily grazed zones can prevent vegetation loss, sustaining rangeland resilience.
4.4. Woody Species Characteristics and Utilization by Cattle and Goats
The composition and characteristics of woody species varied significantly across grazing zones, influencing their utilization by cattle and goats. Heavily grazed zones exhibited lower woody species richness and density compared to moderately and lightly grazed areas. This pattern can be attributed to intense grazing pressure and trampling, which impede the recruitment and growth of woody seedlings and increase vulnerability to herbivory [
5,
10]. Heavy grazing not only reduces species richness but also alters the structural attributes of woody vegetation, such as basal area and canopy cover, which can have cascading effects on forage availability and habitat quality for both livestock and wildlife [
40]. In contrast, lightly grazed areas had significantly higher woody species richness and density, likely due to reduced browsing pressure and less frequent disturbances. The presence of diverse and dense woody vegetation in lightly grazed zones suggests that lower grazing intensities allow for better recruitment and survival of woody species, promoting a balanced coexistence of grasses and shrubs [
41]. This finding aligns with the grazing optimization theory, which posits that moderate to light grazing can enhance plant community diversity by creating spatial heterogeneity in plant resources, thereby preventing the dominance of a few competitive species [
42].
Woody species such as
Acacia karroo and
Dichrostachys cinerea were more abundant in heavily grazed zones, indicating their resilience and ability to thrive under high grazing pressure. This finding is consistent with other studies which identify these species as indicators of rangeland degradation due to their ability to proliferate in overgrazed environments where the herbaceous layer is depleted [
4,
10]. The encroachment of such woody species can significantly alter rangeland dynamics, as their dense growth forms reduce grass cover and compete for resources, leading to decreased forage availability for grazers like cattle [
39]. The dominance of
Dichrostachys cinerea, for instance, has been reported to impede grass regrowth and reduce overall pasture productivity, thereby impacting livestock-carrying capacity (Samuels et al., 2016) [
12].
The utilization of woody species by cattle and goats also reveals distinct patterns, reflecting their different dietary preferences and foraging behaviors. Cattle, being predominantly grazers, primarily utilized woody vegetation during the dry season when herbaceous forage was scarce. Their preference for woody species with palatable leaves and accessible branches, such as
Terminalia sericea and
Combretum molle, underscores the role of these plants as supplementary forage resources during periods of forage scarcity [
16]. According to [
21], such species provide critical nutrients, especially protein and minerals, which help maintain cattle body condition and reduce weight loss during dry spells. Goats, on the other hand, are mixed feeders with a strong preference for browse, showing a higher utilization of woody species across all seasons. They consume a wider range of woody species, including less palatable ones like
Senegalia mellifera and
Euclea divinorum, due to their ability to selectively browse on leaves, twigs, and even bark [
22]. This adaptability allows goats to exploit a broader range of forage resources and maintain productivity under varying environmental conditions [
4]. The higher utilization of woody species by goats, especially in heavily grazed zones, underscores their competitive advantage in degraded rangelands where herbaceous forage is limited [
20].
Moreover, the results indicate that the characteristics of woody species, such as canopy cover, basal area, and shrub height, significantly influence livestock dietary preferences. Cattle tend to utilize woody species with larger basal areas and lower canopy heights, as these features allow easier access to foliage and browsing material [
10]. However, goats prefer species with dense canopies and taller heights, which provide a greater variety of browsing options, including leaves, pods, and fruits [
43]. This finding is supported by the work of [
39], who noted that goats’ ability to reach higher browse material and selectively feed on specific plant parts gives them a foraging advantage in areas with dense woody encroachment. The impact of grazing intensity on woody species utilization is also evident. Increased browsing pressure in heavily grazed zones leads to significant changes in woody plant structure and regeneration dynamics. High browsing intensity can reduce shoot and leaf biomass, affecting the reproductive capacity of woody plants and their ability to withstand environmental stress [
40]. This, in turn, can lead to shifts in species composition, favoring resilient but less palatable species such as
Ziziphus mucronata, which further reduces the forage quality available to livestock [
13].
4.5. Livestock Diet Overlap and Resource Competition Index
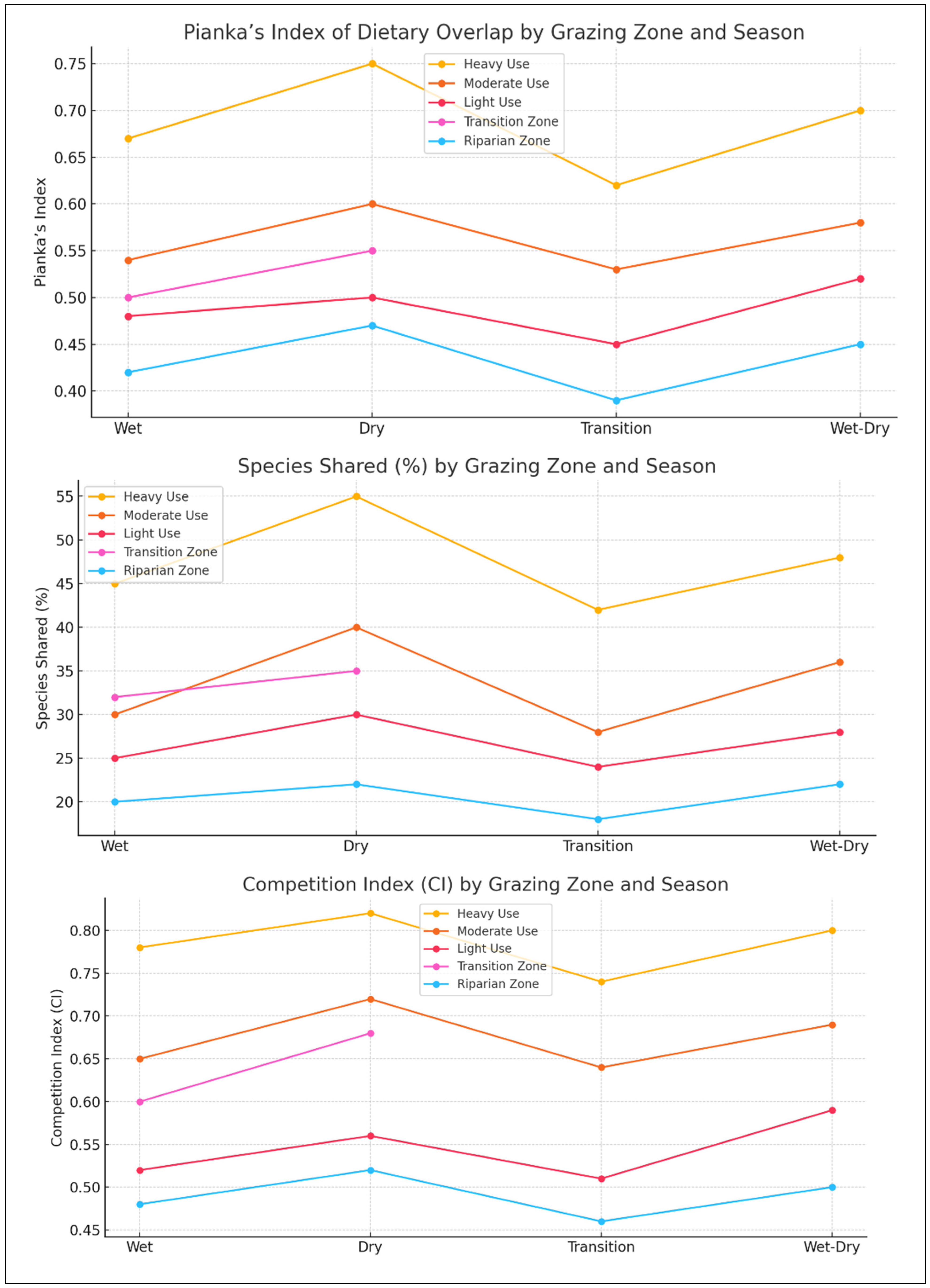
The analysis of livestock diet overlap and the resource competition index reveals important insights into interspecific resource utilization and the potential for competition between cattle and goats in different grazing zones. The degree of dietary overlap between cattle and goats varied significantly across grazing intensities, reflecting differences in their foraging strategies and resource availability in each zone. In heavily grazed areas, dietary overlap was highest, suggesting a high degree of competition for limited forage resources. This observation aligns with the findings of [
44], who reported that when forage availability is low, competition between herbivores intensifies, potentially leading to reduced intake and body condition for one or both species. The high dietary overlap in heavily grazed zones indicates that both cattle and goats are forced to consume similar forage species, such as
Acacia karroo and
Dichrostachys cinerea, which are known to proliferate in degraded areas [
45]. This overlap is likely driven by the scarcity of preferred herbaceous species and the encroachment of less palatable woody species, which become the primary available forage. This pattern can be detrimental to cattle, which primarily rely on grasses for their diet, as they may be outcompeted by goats, which are better adapted to browsing [
17]. Goats, being more flexible feeders, can shift their diet more readily to include a higher proportion of woody species and browse, thus gaining a competitive advantage in environments where grass availability is low [
10].
In contrast, moderate and lightly grazed zones exhibited lower levels of diet overlap between the two species. In these areas, the greater availability and diversity of forage resources, including a balanced mixture of grasses and woody species, reduce direct competition for the same plant species. This finding aligns with the niche differentiation theory, which posits that resource partitioning in more heterogeneous environments allows coexisting species to exploit different forage resources, thereby minimizing direct competition [
46]. For example, in lightly grazed zones, cattle predominantly graze on perennial grasses such as
Themeda triandra and
Digitaria eriantha, which are abundant and provide high nutritional quality [
9]. Meanwhile, goats focus on a mix of grasses and palatable shrubs such as
Grewia flava and
Boscia albitrunca, reducing the likelihood of significant dietary overlap [
3]. The resource competition index further supports these findings by quantifying the intensity of competition between the two species across grazing zones. The index values were highest in heavily grazed areas, indicating severe competition for limited forage. High competition indices in degraded rangelands have been documented in several studies, including in work by [
39], who found that heavy grazing pressure leads to the overutilization of key forage species, resulting in lower dietary intake and poorer animal performance. The implications of high competition are particularly concerning for cattle, which are less adaptable to shifts in forage type compared to goats. Cattle often experience reduced weight gain and reproductive performance under conditions of high dietary competition [
7].
In moderately grazed zones, the competition index was significantly lower, indicating that both species were able to utilize different forage resources with minimal overlap. This finding aligns with studies by [
40], which observed that moderate grazing promotes a more diverse plant community, thereby supporting niche partitioning among herbivores. The presence of a diverse herbaceous layer, combined with sufficient woody cover, provides complementary forage resources that meet the dietary needs of both grazers and browsers without intense competition [
47]. The lowest competition index was recorded in lightly grazed zones, where resource availability is highest and dietary overlap is minimal. This finding underscores the importance of maintaining moderate to low grazing intensities to promote forage heterogeneity and reduce interspecific competition [
5]. Sufficient grass biomass and a diverse array of woody species enable cattle and goats to select forage that aligns closely with their feeding preferences. For instance, goats can utilize taller shrubs and woody species for browsing, while cattle have access to an abundance of high-quality grasses, resulting in reduced competition and improved livestock productivity [
10]. These results are critical for rangeland management, particularly in areas where multiple livestock species graze simultaneously. Managing grazing intensity to maintain a balance between herbaceous and woody vegetation can reduce resource competition and enhance the productivity of both cattle and goats. Strategies such as rotational grazing and controlled browsing can help mitigate the negative impacts of overgrazing and promote the sustainable use of rangeland resources [
39]. By ensuring that grazing pressure is kept within sustainable limits, rangeland managers can support higher forage availability and quality, thereby minimizing competition and promoting the coexistence of different livestock species [
48].
4.6. Seasonal Changes in Woody Species Recruitment and Mortality Rates
The results indicate significant seasonal variations in woody species recruitment and mortality rates across different grazing zones, likely influenced by varying grazing pressure, climatic conditions, and species-specific resilience. The heavy-use zones displayed negative net woody regeneration for all assessed species, suggesting that recruitment is unable to keep pace with high mortality rates. This trend is consistent with the findings of [
10], who reported that high grazing pressure often leads to elevated mortality in young woody plants due to trampling and browsing by livestock, compounded by competition with herbaceous species for limited soil moisture and nutrients. In particular, the species
Acacia karroo and
Dichrostachys cinerea exhibited higher mortality rates compared to their recruitment in heavily grazed zones, resulting in a net loss of 25 and 30 plants/ha, respectively. These results reflect the susceptibility of these species to intensive browsing, which disrupts their growth and regeneration patterns [
13]. Intensive grazing pressure often reduces the capacity of woody species to establish and survive, as younger saplings are repeatedly browsed or damaged by livestock. Additionally, heavy grazing tends to alter soil structure and reduce seedling establishment, further contributing to the observed negative net regeneration [
16]. Conversely, in moderate- and light-use zones, a more balanced relationship between recruitment and mortality was observed, with positive net woody regeneration values across most species. This positive net regeneration in moderate-use zones, for example, for species like
Terminalia sericea (+50 plants/ha) and
Combretum molle (+46 plants/ha), suggests that reduced grazing pressure allows for better recruitment and survival rates. This pattern aligns with the findings of [
2], who noted that moderate grazing can promote a diverse plant community by reducing the dominance of certain species and allowing others to regenerate successfully. The presence of sufficient soil cover and lower trampling intensity in these zones likely provides a more conducive environment for seedling establishment and growth, supporting higher regeneration rates.
Interestingly, the light-use zones showed the highest recruitment rates across all assessed species, with
Sclerocarya birrea and
Terminalia prunioides achieving net gains of +87 and +74 plants/ha, respectively. These findings indicate that minimal disturbance fosters the best conditions for woody species regeneration. Studies by [
14,
39] support this observation, suggesting that light grazing reduces the intensity of competitive interactions and physical damage to young plants, thereby enhancing their survival and growth potential. Furthermore, light-grazing zones typically experience less soil compaction, which improves water infiltration and nutrient availability—key factors for successful woody species recruitment [
36]. The riparian zones exhibited the highest net woody regeneration values, particularly for species such as
Vachellia tortilis (+95 plants/ha) and
Faidherbia albida (+98 plants/ha). This trend can be attributed to the more favorable microclimatic and soil moisture conditions typically found in riparian areas, which enhance seedling establishment and survival compared to drier upland areas [
49]. The high regeneration in these areas, despite being accessible to livestock, may also be due to the resilience of these riparian species, which are adapted to periodic disturbances such as flooding and grazing. As a result, riparian zones act as refuges for woody species, allowing them to maintain viable populations even under moderate grazing pressures [
10]. However, the observed patterns of recruitment and mortality suggest that without proper management, heavy-use zones may continue to experience a decline in woody species density and diversity, potentially leading to shifts in vegetation structure and function over time. Negative net woody regeneration values in these zones could contribute to long-term woody species loss and an increase in bare ground, exacerbating soil erosion and land degradation [
4]. Therefore, targeted management interventions, such as the rotational grazing or seasonal resting of heavily grazed areas, are essential to reduce pressure on young plants and promote their successful establishment [
39].
4.7. Soil Characteristics Across Grazing Zones
The analysis of soil characteristics across different grazing zones revealed distinct variations in key properties such as pH, organic carbon content, total nitrogen, available phosphorus, and bulk density. These variations demonstrate the impact of grazing intensity on soil health and fertility and align with findings from similar studies in rangeland ecosystems, which indicate that grazing pressure affects soil nutrient dynamics, physical structure, and overall soil quality [
17,
50]. Specifically, soil pH values increased progressively from heavy-use zones (pH 5.8) to light-use zones (pH 6.8), with statistically significant differences (
p = 0.03). The lower pH in heavily grazed areas can be attributed to high grazing intensity, which compacts the soil, reduces infiltration, and increases surface runoff, thereby promoting the leaching of base cations such as calcium and magnesium [
10]. As grazing intensity decreases, reduced soil compaction and increased vegetative cover help mitigate leaching, stabilize pH levels, and create more favorable conditions for plant growth [
51].
Organic carbon content exhibited a clear gradient, ranging from 1.2% in heavy-use zones to 2.4% in light-use zones (
p = 0.04), indicating that grazing intensity significantly influences soil organic matter (SOM) accumulation. Heavy grazing often leads to the removal of plant biomass, reduced root density, and less organic matter input into the soil [
40]. Consequently, soil organic carbon tends to be lower in heavily grazed areas due to the diminished contribution of leaf litter and root exudates, which are key sources of organic carbon [
23]. In contrast, light-use zones experience reduced disturbance and higher vegetative cover, facilitating greater accumulation of organic residues and enhancing SOM content. This pattern is consistent with findings from [
52], who reported that moderate to light grazing can improve soil carbon content by promoting plant growth and organic matter inputs. Total nitrogen content showed a similar trend, with significantly lower values in heavy-use zones (0.07%) compared to light-use zones (0.12%) (
p = 0.02). The reduction in nitrogen levels in heavily grazed areas can be linked to increased soil erosion, lower soil organic matter (SOM) content, and reduced nitrogen fixation by leguminous species due to overgrazing [
4]. Legumes, which contribute nitrogen to the soil through biological nitrogen fixation, are particularly sensitive to heavy grazing pressure. Their decline in heavily grazed areas results in lower nitrogen availability in the soil [
21]. In contrast, moderate- and light-use zones maintain a higher proportion of legumes and vegetative cover, which improves nitrogen retention and cycling within the soil system [
10]).
Phosphorus availability also varied significantly across grazing zones, with the lowest concentration recorded in heavily grazed areas (5.6 mg/kg) and the highest in light-use areas (12.5 mg/kg) (
p = 0.01). Phosphorus availability is strongly influenced by organic matter content and soil microbial activity, both of which are typically reduced under high grazing pressure [
15]. Heavy grazing can disrupt the phosphorus cycle by compacting the soil, reducing microbial activity, and limiting the decomposition of organic matter, leading to lower phosphorus availability [
14]. In contrast, light grazing promotes microbial activity and organic matter decomposition, enhancing phosphorus mineralization and availability to plants [
5]. Bulk density values were significantly higher in heavy-use zones (1.4 g/cm
3) compared to light-use zones (1.1 g/cm
3) (
p = 0.05), indicating increased soil compaction in heavily grazed areas. Soil compaction, often associated with intensive grazing, reduces pore space, limits water infiltration, and restricts root growth [
39]. These effects can have a cascading impact on plant growth and soil nutrient availability, ultimately leading to reduced soil productivity and vegetation cover [
4]. In contrast, light grazing minimizes compaction and maintains soil structure, thereby improving bulk density and supporting a more stable and productive soil environment [
11].
4.8. Proximate Nutrient Composition of Fecal Samples from Cattle and Goats
The analysis of the proximate nutrient composition of fecal samples from cattle and goats across varying grazing zones revealed distinct differences in crude protein, fiber content, digestibility, and ash percentages. These findings highlight the influence of diet quality and grazing intensity on nutrient intake and digestion. They are consistent with similar studies on livestock nutrition, which demonstrate that fecal nutrient content serves as a reliable indicator of forage quality and animal nutritional status [
12,
41]. Crude protein (CP) content in fecal samples was notably higher in light-use grazing zones for both cattle (15.1 ± 0.6%) and goats (14.7 ± 0.5%) compared to the lower values observed in heavy-use zones (12.4 ± 0.8% for cattle and 11.8 ± 0.9% for goats). The reduced CP content in heavily grazed areas suggests that animals are consuming lower-quality forage with diminished protein availability, likely due to overgrazing and the resultant decline in high-quality, protein-rich forage species [
6,
10]. Overgrazing can deplete preferred forages such as legumes, leading to a shift in diet composition toward grasses and woody plants that may have lower protein content, thereby reducing overall protein intake [
53]. In contrast, light grazing maintains a higher abundance of protein-rich species, allowing animals to select higher-quality forage, which is reflected in the increased fecal protein content [
18].
Fiber content, as measured by neutral detergent fiber (NDF), was highest in fecal samples from heavy-use zones for both cattle (60.3 ± 1.3%) and goats (62.1 ± 1.4%). This suggests that animals in these areas are consuming more mature and fibrous plant material, which has lower digestibility and energy content [
10]. High fiber content in feces is often an indicator of low forage quality, as increased fiber is associated with lower nutrient availability and reduced intake of digestible nutrients [
17]. These findings align with other studies which show a strong positive correlation between grazing pressure and dietary fiber, resulting from the depletion of preferred forage species and a subsequent reliance on less palatable, more fibrous vegetation [
20]. The lower fiber content observed in light-use zones (50.7 ± 1.2% for cattle and 52.6 ± 1.1% for goats) indicates higher-quality forage with a lower proportion of mature stems and higher leaf-to-stem ratios, thereby enhancing nutrient digestibility [
22]. Digestibility values followed a similar trend, being significantly higher in light-use zones (72.8 ± 1.8% for cattle and 71.3 ± 1.9% for goats) compared to heavy-use zones (65.2 ± 2.1% for cattle and 64.1 ± 2.4% for goats). This variation in digestibility is linked to forage quality, as light-grazing zones typically contain a greater abundance of green, leafy material that is easier to break down and assimilate [
50]. Digestibility is a key determinant of energy intake, and the lower digestibility rates observed in heavily grazed zones can negatively impact animal performance due to reduced energy extraction from consumed forage, ultimately leading to decreased weight gain and milk production [
49]. The results corroborate findings from other rangeland studies, which report that digestibility declines under heavy grazing due to the increased consumption of woody and fibrous materials [
45].
Ash content was also highest in the feces of animals grazing in heavy-use zones (7.5 ± 0.5% for cattle and 8.0 ± 0.6% for goats) compared to light-use zones (6.1 ± 0.3% for cattle and 6.5 ± 0.4% for goats). High ash content in feces can indicate a greater intake of soil and low-quality plant material, often associated with overgrazed pastures where nutrient-rich forage is scarce [
10]. Animals in degraded areas may ingest more soil while grazing close to the ground or consume plants with high mineral content, resulting in elevated fecal ash levels [
38]. This trend is supported by [
15], who found that increased grazing pressure led to higher ash content in feces due to soil contamination and lower forage quality. Interestingly, goats consistently exhibited higher fiber and ash content compared to cattle across all grazing zones, while cattle showed higher crude protein and digestibility values. These differences are likely due to species-specific foraging strategies and digestive adaptations [
10]. Goats are known to be more selective feeders, often consuming a greater proportion of woody plants and shrubs that have a higher fiber and ash content than the grasses preferred by cattle [
4]. Additionally, the rumen morphology and microbial populations of cattle are better adapted to breaking down fibrous material, which could explain their higher digestibility and protein utilization compared to goats [
36].
4.9. Rangeland Degradation and Restoration Strategies
Rangeland degradation remains a pressing challenge, primarily driven by overgrazing, poor land management, and climate variability. These factors contribute to vegetation loss, soil erosion, and reduced productivity, negatively affecting livestock and livelihoods [
54,
55]. The findings highlight that grazing management practices, such as rotational grazing and controlled grazing zones, can mitigate degradation by promoting balanced plant recovery and soil stability, aligning with principles outlined by [
27,
32]. Restoration efforts, particularly in heavily degraded areas, should focus on reseeding with native species and reducing grazing intensity to facilitate ecosystem recovery and sustain biodiversity. Effective restoration can help reverse degradation, improve forage availability, and support livestock productivity in the long term [
10,
56].
4.10. Implications for Livestock Management and Future Rangeland Sustainability
The impact of grazing on forage quality and ecosystem resilience has crucial implications for livestock management. As observed, heavy grazing reduces forage nutritional quality, demanding supplemental feeding strategies to meet livestock dietary needs [
15]. By contrast, moderate grazing supports higher forage quality and biodiversity, which are beneficial for livestock growth and ecosystem health [
17]. Seasonal variations further underscore the need for adaptive management; maintaining livestock productivity requires access to critical grazing refuges, such as riparian zones, during dry periods to offset forage scarcity [
25]. The integration of these findings suggests that sustainable rangeland management should prioritize adaptive grazing strategies, restoration, and the selective thinning of woody species to balance forage quality and accessibility. Future strategies could benefit from continued monitoring of species composition and resilience indicators, aiding the development of region-specific approaches for sustainable livestock systems in semi-arid rangelands [
42,
51].
4.11. Study Limitations
This study has several notable limitations. First, it employed a cross-sectional design, capturing only a snapshot of the interactions between vegetation and livestock at a specific point in time. Consequently, it fails to account for seasonal and annual variations in vegetation dynamics and livestock productivity. This limitation restricts our understanding of how these interactions evolve over longer periods, which is crucial for effective rangeland management. Second, this study primarily focused on biophysical factors influencing livestock productivity, neglecting the potential impact of socio-economic factors such as herd management practices, breed differences, and market conditions. These variables can significantly affect livestock performance and are essential for a more comprehensive understanding of productivity drivers. Additionally, the reliance on fecal analysis to infer dietary preferences may introduce inaccuracies, as this method may not fully capture complete dietary intake, particularly when distinguishing between similar forage types. This study also examined only two livestock species, cattle and goats, limiting its applicability to other grazers or browsers in similar ecosystems. Finally, this research did not consider the potential interactions between domestic livestock and wildlife, which could significantly influence grazing dynamics and resource competition. This omission may lead to an incomplete understanding of how grazing intensity impacts rangeland vegetation and livestock productivity.






{kind=link}
{kind=link}
{kind=link}
{kind=link}